
Abstract
INTRODUCTION:
Adipose derived Stem Cells (ASCs) have been proven to play a key role in tissue regeneration. However, exposure to large amounts of cigarette smoke can drastically diminish their function. Erythropoetin (EPO), can modulate cellular response to injury. Therefore, we investigated the ability of EPO to restore the regenerative function and differentiation capacity of ASCs.
MATERIAL AND METHODS:
Human ASCs were isolated from abdominoplasty samples using standard isolation procedures. Cell identity was established by means of Fluorescence Activated Cell Scanning. Subsequently, isolated ASCs were cultivated with cigarette smoke extract both with and without EPO. Parameters investigated included cellular metabolic activity, adipogenic and osteogenic differentiation capacity, and in vitro wound closure capacity. For further enhancing wound closure, EPO was combined with Granulocyte Macrophage Colony Stimulating Factor (GM-CSF) or Stromal Derived Factor-1 alpha (SDF-1 a).
RESULTS:
Cigarette smoke reduces adipogenic differentiation, the osteogenic differentiation capacity as well as the in vitro wound healing ability of human derived ASCs. EPO did not change metabolic activity of ASCs significantly. The addition of EPO could partially restore their function. The combination of EPO with GM-CSF or SDF-1 did not result in a synergistic effect regarding wound healing ability.
CONCLUSION:
Exposure to cigarette smoke significantly reduced the regenerative potential of ASCs. Treatment of ASCs exposed to cigarette smoke with EPO has the potential to partially restore their function.
Introduction
Stem cell-based therapies remain at the forefront of tissue regeneration as they represent a unique approach with exceptional healing potential [1]. Therapeutic use of these multipotent progenitor cells for treatment of chronic wounds has shown promising results in basic research as well as in pre-clinical studies [2–5]. With their differentiation capacity and their ability to release cytokines and soluble growth factors, these cells represent pivotal players for almost every regeneration process [4].
However, the mechanisms of stem cell-based therapies in terms of complex signaling cascades, environmental influences, or epigenetic modulation are still not fully understood. While it is undoubted that Mesenchymal Stem Cells and in particular Adipose derived Stem Cells (ASCs) play a key role in tissue regeneration, smoking of cigarettes has been proven to diminish their function [6–8]. Recent studies have confirmed that even a small amount of cigarette smoke exposure can have harmful effects on the regenerative potential of ASCs [6].
Erythropoietin (EPO) is an endogenous hormone secreted by fibroblasts of the renal parenchyma, as reaction to the partial oxygen pressure in the peripheral blood [9]. EPO acts via two receptors- a homodimer EPO specific receptor, and a second heterodimer receptor, that needs dimerization with the CD131 receptor, that represents a target for GM-CSF, and several interleukins [10].
Similar to EPO the activity of the chemokine stromal cell-derived factor-1 alpha (SDF-1α) is central in ischemic neovascularization and in the response to ischemic events. SDF-1α mediates the mobilization and trafficking of stem or progenitor cells expressing the receptor CXCR4 [9–12]. It was previously demonstrated that induction of SDF-1α is dependent on stabilization of hypoxia-inducible factor-1 alpha (HIF-1α), a transcription factor that serves as a central regulator in the tissue response to hypoxia [4]. HIF-1α orchestrates this response by inducing cell-type specific gene expression changes that result in increased erythropoietin (EPO) production in kidney and liver.
EPO is the most important hematopoietic growth factor controlling the differentiation of erythrocytes in the bone marrow. In addition, Jelkmann et al. demonstrated that EPO has the ability to modulate cellular responses in different types of injuries, and is therefore, an interesting additive for wound healing purposes [11].
Tissue injury results in a variety of very specific cascades of cellular and factor based reactions [12]. Resulting in wound repair being one of the most difficult biological processes that relies on an interplay of several pathways. The precise function of these pathways is modulated by a complex signalling system that includes several growth factors, cytokines and chemokines and leads to the regulation of the three phases of wound healing: inflammation, formation of granulation tissue and tissue remodelling [12, 13].
Comprehensive communication among all these features is crucial for complete and effective tissue regeneration. Loss or depletion of one of these factors as evidenced by patients that are aged, diabetic, malnourished, suffer from malignancy, hypoperfusion, infection or are obese, leads to a breakdown of this molecular system and consequently, to the formation of chronic wounds [14, 15].
Smoking has been proven to be one of the factors contributing to impaired wound healing [16–18]. Cigarette smoke contains a complex mixture of chemicals with about 7,000 different compounds. 250 of these are known to be deleterious and can rapidly be absorbed into the blood flow [19]. The toxic effects of cigarette smoke considerably increases the risk of cardiovascular disease, rheumatoid arthritis, osteoporosis and cancer [20–23]. It also undermines the conditions necessary for proper tissue repair. Cigarette smoke causes slower cell division and DNA damage, inhibits healing time, and the ability to repair tissues by reducing nutritional blood flow [6]. Nevertheless, the exact mechanisms of how cigarette smoke interplays with wound healing, which pathways are affected, and even more importantly, the possibility to counteract its effect, are still under investigation. Therefore, we explored the efficacy of EPO in the reversal of smoke related changes to ASCs.
Material and methods
All studies have been carried out in accordance with the Ethical Commission of the Technical University of Munich (2754/10). All patients have given informed consent to take part in this study.
Human ASC isolation and cultivation
Human ASCs were obtained from lipoaspirates from patients undergoing fat removal surgeries at the Department of Plastic Surgery and Hand Surgery at the University Hospital rechts der Isar (Munich, Germany). The cells were isolated according to a previously described protocol. ASCs were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Biochrom, Berlin, Germany) supplemented with 10% Fetal Bovine Serum Gold (FBS; PAA, Pasching, Austria) and 1% antibiotic/antimycotic solution (Capricorn Scientific, Ebsdorfergrund, Germany). Medium was changed twice a week and cells were expanded till they were ready to be used at passage three. All experiments were carried out with at least three donors (N = 3) in triplicate (n = 3).
Fluorescence activated cell scanning (FACS) - Analysis
Cultivated cells in passage 1 to 3 were treated with Trypsin, washed twice with PBS, and then characterized for ASC phenotype by flow cytometry. We used a commercially available standardized human ASC flow cytometry kit (MSC Analysis Kit, BD Stemflowtrademark). This assay is based on the phenotypic signature of multipotent mesenchymal stem cells described by The Mesenchymal and Tissue Stem Cell Committee of the International Society for Cellular Therapy (ISCT). This is a multicolor flow cytometry assay containing pre-conjugated and pre-titrated cocktails of the ISCT-defined positive expression markers (PerCP-Cytrademark5.5 CD105/APC CD73/FITC CD90) and negative expression markers (PE-CD45/CD34/CD11b/CD19/HLA-DR). Unstained controls, isotype controls, antibody-stained probes as well as cellular compensation controls were used.
Cigarette smoke extract preparation
Cigarette smoke extract (CSE) was prepared freshly prior to each experiment and applied to cells within 10 minutes, according to ISO 3308 : 2012 conditions. Briefly, a gas-washing bottle with 25 ml DMEM was connected to a peristaltic pump with an adjustable suction volume (Watson-Marlow 501, Rotterdam, Netherlands). Three Marlboro red cigarettes, with the filter removed, were smoked for 15 minutes. Once a minute (for about two seconds), the valve was closed in order to mimic inhalation. The resulting extract was then filtered (100 μm mesh; BD Biosciences, San Jose, CA, USA) to remove particulate matter and then further filtered (0.22 μm sterile syringe filter; TPP, Trasadingen, Switzerland) to sterilize the extract before being added to the cells at the respective dilution of (% v/v).
Metabolic activity testing
The metabolic activity of the ASCs was determined using MTT (3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide) assay (Sigma, Carlsbad, CA). ASCs were seeded in 48 well plates with 5000 cells per well. Twenty-four hours after the initial seeding, the medium was removed and replaced with fresh medium or medium containing Epoetin beta (NeoRecormon, Roche, Frankfurt, Germany) in four different concentrations. The cells were incubated for another 12, 24, 48 or 72 hours. MTT reagent was added to each well and incubated for 2 h under standard cell culture conditions. DMSO was used to dissolve the resulting formazan and the optical density was measured with a Mithras LB 940 absorbance reader (Berthold Technologies, Bad Wildbad, Germany) at 560 nm. Blank values from empty wells were then subtracted from sample values.
Adipogenic differentiation
Cells were seeded at a density of 21,000/cm2 and left to attach for 2–3 days, upon which, they were stimulated with CSE and adipogenic induction medium (Lonza). Medium changes alternated between induction and maintenance medium following manufacturer’s instructions. After 21 days in culture, Oil Red O (Sigma-Aldrich, St Louis, MO, USA) was used to stain and quantify the resulting lipid droplets. Briefly, the cells were fixed with formalin and rinsed with PBS, and then treated with 60% isopropanol to eliminate background interference. The wells were then allowed to dry and Oil Red O solution was added for 10 min. Subsequently, they were rinsed with water and imaged. To elute the Oil Red O solution, 100% isopropanol was added for 10 min on an orbital shaker and the optical density of the solution was measured at 490 nm, 0.5 sec.
Osteogenic differentiation
Cells were seeded at a density of 3,100/cm2 and left to attach overnight before being stimulated with CSE and osteogenic induction medium (Lonza). After 21 days in culture, the cells were stained with Alizarin Red S (Sigma-Aldrich) to visualize and quantify calcium deposition. Briefly, the cells were fixed with formalin, rinsed with deionized water, and stained with 0.5% Alizarin Red S solution. They were then rinsed with deionized water and imaged. The quantification of the staining was carried out and the absorbance was read at 530 nm.
Evaluation of ASC migration
Migration of ASCs was tested by using an in vitro Wound Healing and Migration Assay (Ibidi GmbH, Martinsried, Germany). 1*104 ASCs were seeded in a 24 well plate with 70 μl DMEM and the ibidi chambers according to the manufacturer’s instructions. After 24 hours, the chambers were removed, and EPO-100, SDF-1a (0.5 μl/ml), GM-CSF (0.5 μl/ml) alone and in combination were used as cultivation-mediums. Evaluation of gap-closure was performed after 48 hours.
Statistical analysis
Differences between groups were assessed using one-way ANOVA test. A p-level of <0.05 was considered significant. All data is given as mean values±the standard deviation. Analysis was performed using the SPSS software package.
Results
Plastic adherence is a valuable method to obtain ASCs
Cultivated cells were analyzed regarding their surface marker profile. We found that the majority of cells fulfill the extensively published ASC-criteria [24–26]. Therefore, we confirm, that our harvesting method and selection by means of plastic adherence represents a simple and valuable method to obtain ASCs (Fig. 1).
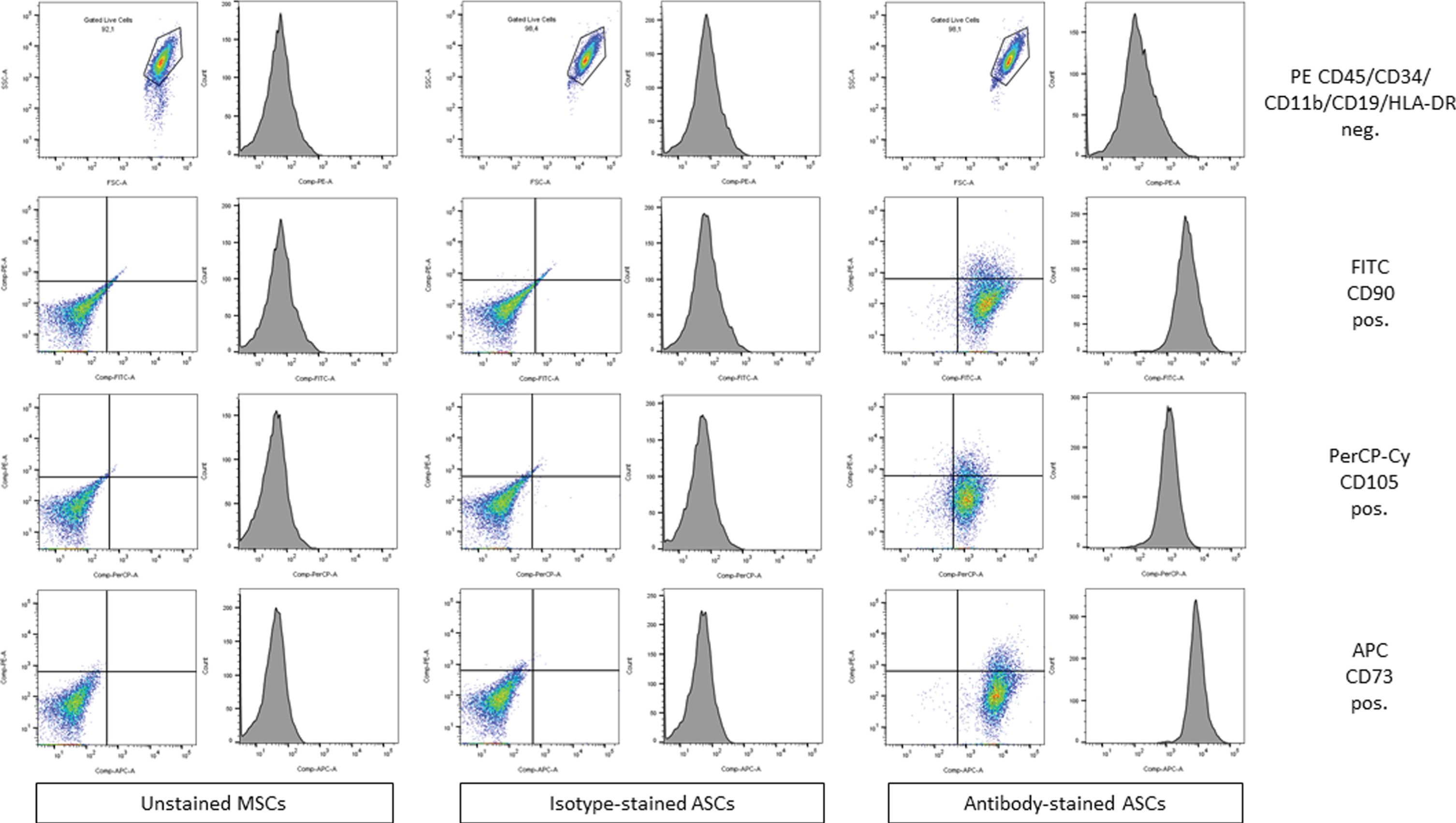
FACS based analyzes of ASCs that have been isolated based on their plastic adherence. CD = cluster of differentiation
As indicated in Fig. 2, no significant results were found. However, a trend towards temporary decreased metabolic activity in EPO treated ASCs was observed. This effect was dose dependent. Where the EPO-25 group showed an optical density of 0.097 (±0.045) after 24 hours of incubation, and the EPO-200 group showed even lower values (0.042 (±0.018)). After 72 hours of incubation, this effect was completely reversed in all groups.
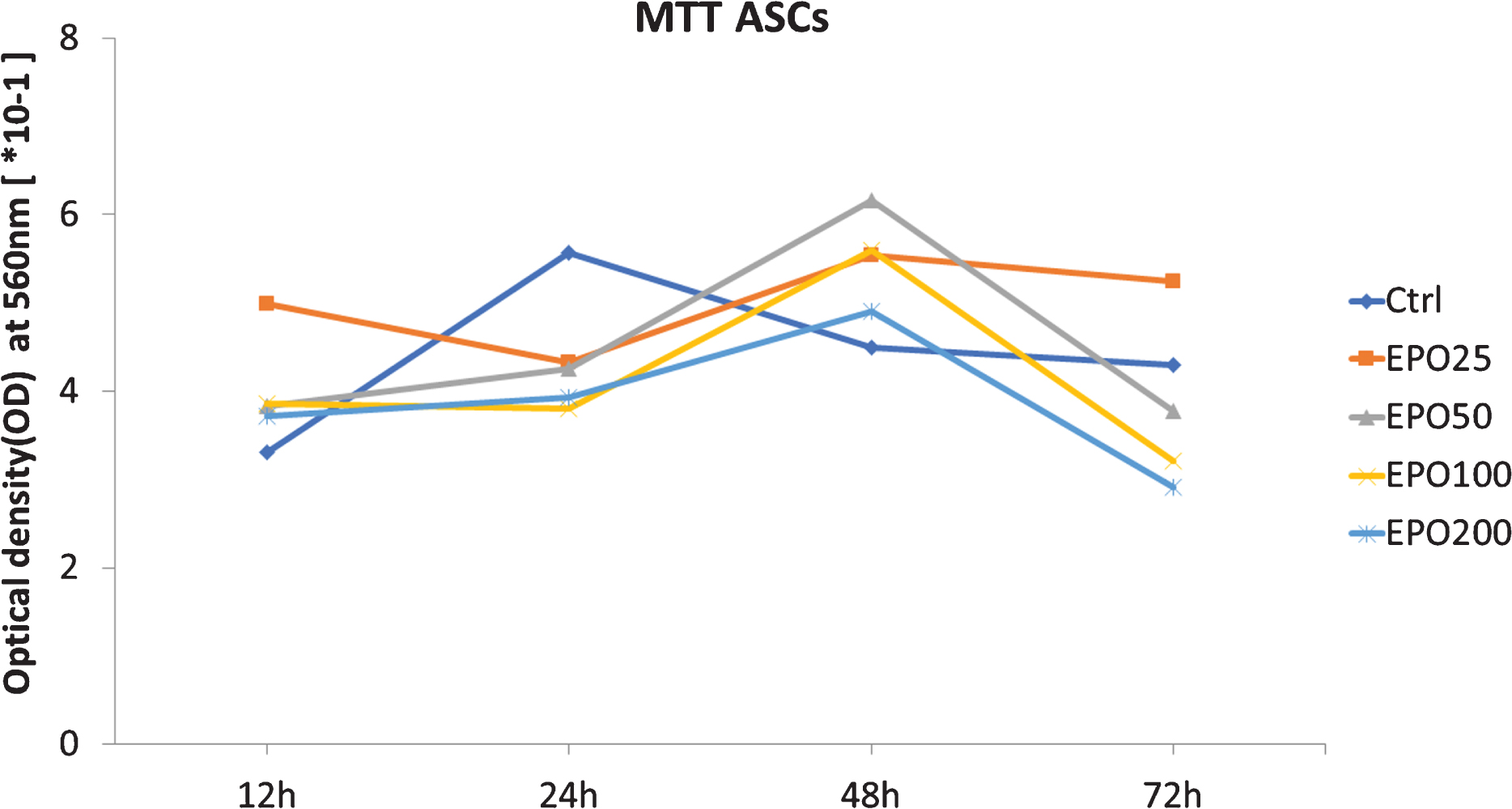
MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide)-assay for measuring metabolic activity of cells.
A trend towards reduced adipogenic differentiation in ASCs that have been treated with CSE (p = 0.163) was observed. When treating cells with diminished adipogenic differentiation capacity due to exposure to CSE, we were only able to demonstrate, that EPO tends to partially restore this ability (p = 0.188) (Fig. 3).
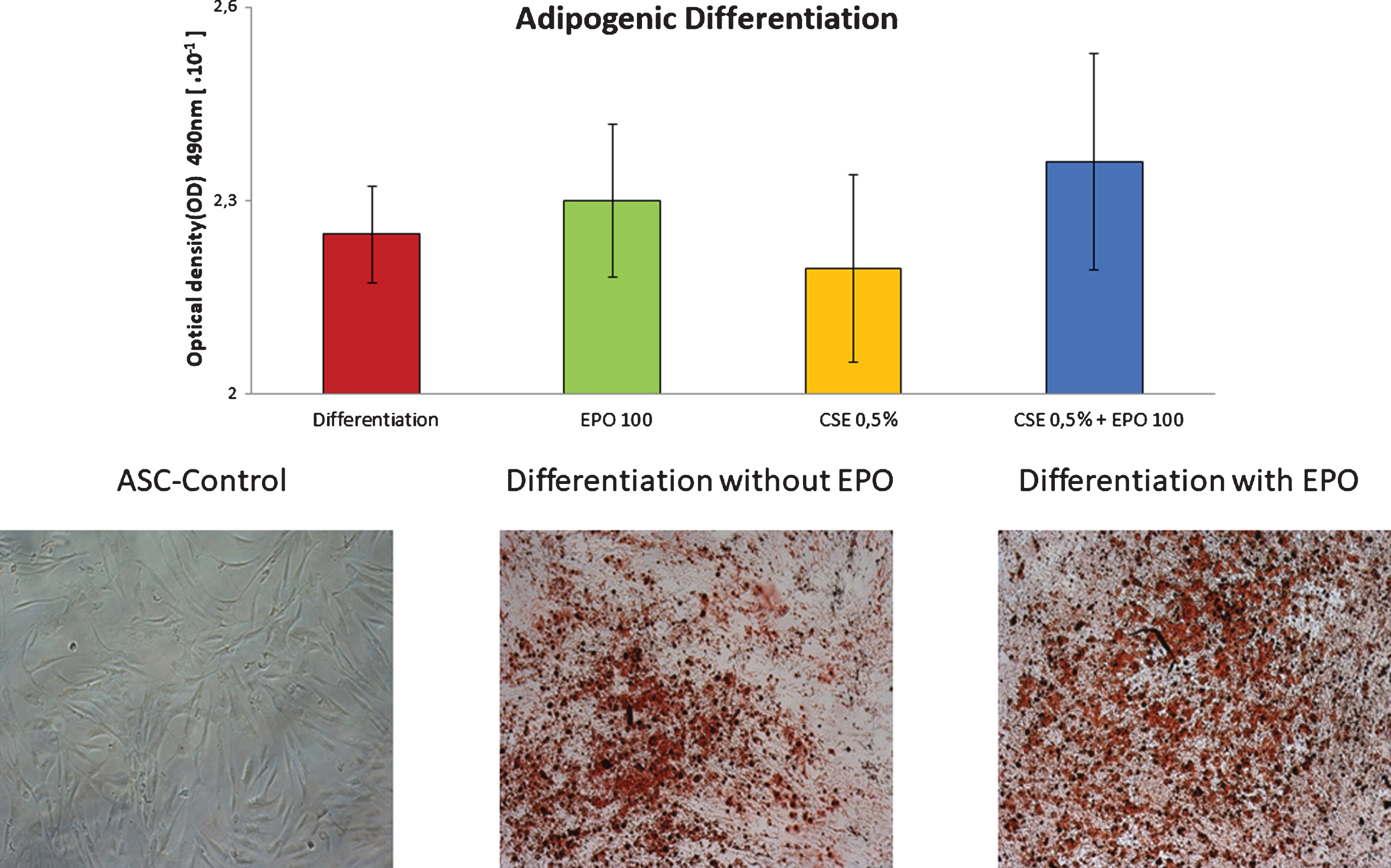
Adipogenic differentiation of MSCs, treated with cigarette smoke extract, with or without EPO-100.
EPO significantly increases osteogenic differentiation (p = 0.00014). While CSE significantly reduces ASCs osteogenic differentiation capacity (p = 0.0000996), EPO can restore the cigarette smoke-attenuated osteogenic differentiation of ASCs (p = 0.000000757) (Fig. 4).

Osteogenic differentiation of MSCs, treated with cigarette smoke extract, with or without EPO-100. CSE significantly reduces ASCs differentiation capacity (p = 0.0000996), EPO significantly increases it (p = 0.00014). In addition, EPO can fully restore the cigarette smoke-attenuated osteogenic differentiation of ASCs (p = 0.000000757).
When performing scratch assays, we found significantly enhanced gap closure of ASCs treated with EPO-100 (p = 0.027). To further enhance and investigate molecular pathways, we used GM-CSF and SDF-1a, two agents that have been shown to significantly interact with the EPO pathway (27). GM-CSF as well as SDF-1a showed a significant increase in wound closure after 24 hours, either alone or in combination with EPO-100 (p = 0.048). Combining EPO either with GM-CSF (p = 0.688) or SDF-1a (p = 0.463) did not lead to a synergistic improvement of ASC migration (Fig. 5).
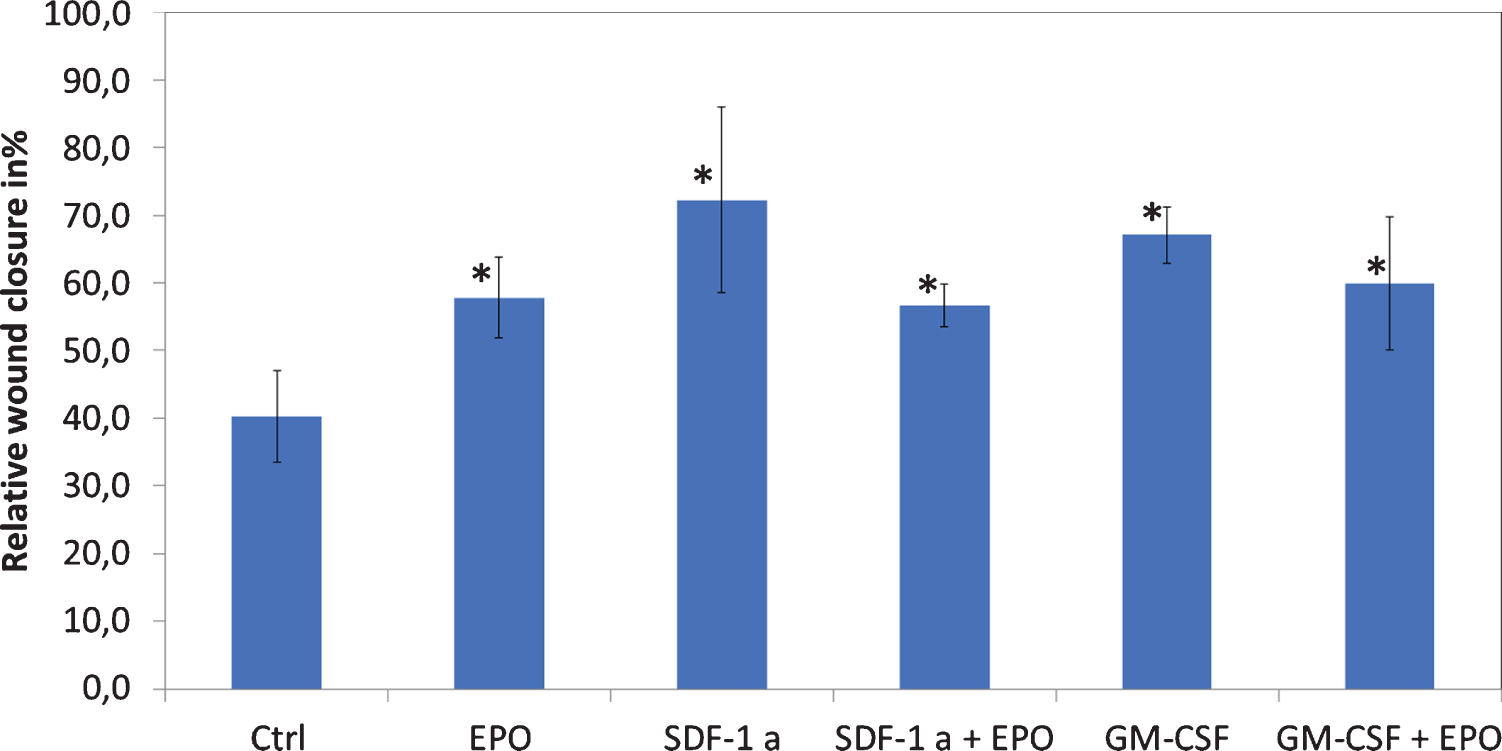
In-vitro wound closure assay of ASCs. SDF-1a = Stromal derived factor-1 alpha; GM-CSF = Granulocyte Macrophage-colony stimulating factor. We found significantly enhanced closure time when treating ASCs with EPO-100 (p = 0.027).
Adipose derived stem cells are crucial for tissue regeneration and repair. They are able to synthesize a variety of cytokines and growth factors, which have essential effects on local cellular dynamics. Therefore, these cells represent trophic mediators releasing pro-angiogenic, chemotactic, and immunomodulatory factors that play significant roles in wound healing, bone and cartilage remodeling, and overall tissue regeneration [8, 9].
Due to their multi potency, their self-renewal capacity, and long-term viability, ASCs are capable of separating into cells of multiple line ages by performing asymmetric divisions [28]. Therefore, they could be beneficial for the development of regenerative therapies.
Nevertheless, several patient dependent factors, such as diabetes, aging, or smoking have been proven to significantly diminish ASC function [1, 29–32]. A recently published study by Barwinska et al. showed that cigarette smoke (CS) significantly compromises ASC related vasculogenesis [32]. While this study highlights that deficiencies in ASC function can be induced by CS, the authors do not provide effective solutions to reverse these effects. Therefore, strategies to reverse these effects are urgently needed.
In previous studies, our group demonstrated major negative impact of cigarette smoke on ASCs. High dose CS leads to significantly increased cell-death [6], smaller amounts (0.5%) of CSE was used in our setting. Confirming the findings of Wahl et al., we were not able to find any significance in regards to migration and adipogenic differentiation. However, contrary to Wahl et al. we found a significantly enhanced osteogenic differentiation ability in the EPO-100 treated group while using the same protocol. These differences might be explained by a high donor-depended variability. Additionally, Wahl et al. did find changes in transcription factor expression, but not in Alizarin Red S staining.
While both, Wahl et al. and Barwinska et al. focused on exploring the mechanisms of how smoking effects ASCs, our study aims to reverse these changes. Most likely, anti-inflammation, inhibition of apoptosis, enhanced angiogenesis and stimulation of regenerative protein secretion can be seen as underlying mechanisms for the beneficial effects of ASCs [33, 34]. All of these mechanisms are explicitly highlighted by Barwinska to contribute to diminished ASC-function, when being exposed to cigarette smoke.
In addition to presenting EPO as a sufficient agent against CS-related changes, we compared EPO to other drugs, which have demonstrated benefit to ASCs. The SDF-1a reduction has already shown to correlate with decreased ASC migration in vitro, while administering SDF-1 could enhance it [35, 36]. Therefore, we used SDF-1a as positive control and found similar results to EPO with an enhanced wound closure. Interestingly there was no synergistic effect, when being combined with EPO. On the contrary the effect was diminished, which could result from a competition for the same receptor hetero-dimers and signaling pathways [37].
Furthermore, Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF) has been tested in regards to its migration rate alone as well as in combination with EPO. Once again, in confirmation with recent findings, GM-CSF did influence ASCs by stimulating their migration. However, the combination of EPO and GM-CSF displayed a weaker effect than GM-CSF alone. This phenomenon has already been presented in previous studies, which have investigated efficacy of EPO and GM-CSF on liver regeneration. The fact that EPO and GM-CSF demonstrate a highly homologous molecular structure might explain this phenomenon [38, 39], based on potential competition regarding receptor binding. Additionally, GM-CSF has exhibited a decrease in EPO-receptors in hematopoietic stem cells [40]. Similar mechanisms could explain our observations.
The usage of EPO to reverse smoke related changes represents an elegant method with a broad and promising translational potential. Many stem cell based therapies are significantly diminished by factors such as smoking [41, 42]. Concurrently, many of these therapies are being tested in phase 1 clinical trials attempting to cure morbidities such as cardiac infarction, or injuries that are closely related to nicotine abuse [43, 44]. Therefore, strategies to reverse smoke induced changes on the regenerative capacities of ASCs are urgently needed. EPO, either as additive for cultivation of ASCs or as a therapeutic agent for patients can fill this gap.
In summary, the regenerative potential of ASCs is largely affected by cigarette smoke. EPO has the ability to restore functionality and the differentiation capacity of ASCs. This represents a novel therapeutic paradigm to correct cellular function compromised by cigarette smoke exposure.
Conclusion
The present study demonstrates that EPO is able to partially restore the differentiation capacity of ASCs as a means to reverse cigarette smoke induced changes. Nevertheless, the precise molecular mechanisms are currently understudied and should be further investigated to enable clinical translation of this novel therapeutic approach.
Conflict of interest
None of the authors has any financial or personal interest in the products mentioned in this article.