
Abstract
This study aimed to investigate the effects of arachidonic acid metabolite epoxyeicosatrienoic acid (EETs) in the apoptosis of endothelial cells induced by tumor necrosis factor-alpha (TNF-α). After human umbilical vein endothelial cells were cultured, TNF-α/ActD, 14, 15-EET, and HMR-1098 were added, respectively, into the culture medium. The apoptosis level of endothelial cells was detected by flow cytometry. After TNF-α/ActD induced endothelial cell apoptosis, flow cytometry staining showed that endothelial cell apoptosis increased significantly, and the apoptotic cells were significantly reduced after the addition of 14, 15-EET. However, the apoptotic cells significantly increased after the addition of HMR-1098. Western Blot results showed that the phosphorylation levels of LC3-II and AMPK were increased after TNF-α/ActD induction, and the increase was noticeable after the addition of 14, 15-EET. However, the phosphorylation levels of LC3-II and AMPK significantly decreased after the addition of HMR-1098. The activity of Caspase-8 and -9 decreased significantly after the addition of 14, 15-EET but increased after the addition of HMR-1098. Arachidonic acid can inhibit TNF-α induced endothelial cell apoptosis by upregulating autophagy.
Introduction
The prevalence and mortality of cardiovascular diseases among urban and rural residents in China have increased in the past decades. In the proportion of disease death, cardiovascular disease was the leading cause of death [1]. Various cells participate in the pathogenesis of cardiovascular diseases. Endothelial cells are foremost among these [2].
Endothelial cells are monolayer flat cells located in the innermost layer of blood vessels and have a wide range of functions. They are vulnerable to external stimulation and damage, including apoptosis and autophagy. Atherosclerosis is known to be the main pathophysiology of cardiovascular diseases, and endothelial cell apoptosis is the main feature of atherosclerosis formation [3]. Arachidonic acid is widely found in endothelial cells, and its metabolism of epoxyeicosatrienoic acids (EETs) has a clear anti-apoptosis effect on endothelial cells [3], but the specific mechanism has not been clarified. As two different forms of programmed cell death (PCD), the correlation between autophagy and apoptosis has focused on basic research in recent years. Apoptosis is the spontaneous and orderly death of cells controlled by genes, while autophagy is a process where cells use lysosomes to degrade their damaged organelles and macromolecular substances, which is a unique phenomenon of eukaryotic cell life. Both apoptosis and autophagy of endothelial cells play an important regulatory role in the development of atherosclerosis [4, 5]. EETs also affect platelet function and diminish platelet aggregation [6]. It has also been found that inhibiting the PI3K-AktmTOR signaling pathway can activate autophagy, and it can protect HUVECs cells from oxidative stress damage and inhibit their apoptosis [7].
In recent years, it has been found that arachidonic acid can produce EET through the metabolism of cytochrome P450 (CYP) surface oxidase. EET can activate the autophagy pathway and inhibit the apoptosis of endothelial cells induced by starvation. This suggests that the apoptosis effect of EET could play a role by activating the autophagy pathway. AMPK is a protein kinase that senses the energy state in cells and regulates metabolism, playing an important role in regulating autophagy.
The present study aims to investigate whether autophagy is also involved in the protective effect of EETs on endothelial cells and its specific mechanisms.
Methods
Materials
Human umbilical vein endothelial cells (HUVECs) were obtained from ScienCell Research Laboratories (Carlsbad, CA, USA. Batch No. 20181107). The human umbilical vein endothelial cell line came from the frozen cell line in our laboratory. The endothelial cell culture medium, growth factor, and Annexin V-FITC kit are products of Life Technologies. HMR-1098, 14, 15-EET, IC3-II (microtubule-associated protein light chain 3-II) antibodies, p-AMPK, t-AMPK antibodies are all Sigma products. This study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of Hainan Maternal and Child Health Hospital (No. 2019-09). All participants provided written informed consent.
Incubation of human umbilical vein endothelial cells with TNF-α, ActD, EET or HMR-1098
Human umbilical vein endothelial cells were cultured in the medium of endothelial cells with calf serum concentration of 10% in the plastic well plates. Part of the old medium was removed from the cultured endothelial cells every other day, and the new medium was replaced. The cultured cells were randomly divided into four groups: Control group, TNF-α/ActD group, TNF-α/ActD+14, 15-EET group (EET group), and TNF-α/ActD+14, 15-EET+HMR-1098 (EET+HMR-1098 group). The blank group was cultured conventionally. The TNF-α/ActD group was treated with TNF-α (10 ng/mL) and ActD (5 ng/mL), respectively, and cultured for 24 hours to induce apoptosis of endothelial cells. In the EET group (eicosapentaenoic acid), TNF-α (10 ng/mL) and ActD (5 ng/mL) were added to induce apoptosis while 14, 15-EET (100 nmol/L) was added. In the EET+HMR-1098 group, TNF-α (10 ng/mL) and ActD (5 ng/mL) were added to induce apoptosis, while 14, 15-EET (100 nmol/L) and HMR-1098 (10 ummol/L) were added. The cells in each group were cultured for 24 hours after the following experimental studies.
Flow cytometry of cell apoptosis
Cell apoptosis was detected by flow cytometry. Trypsin-EDTA (0.5% Trypsin/0.02% EDTA) was added to the cells in each group. The cells were collected into a 10 mL centrifuge tube, and the cell number was adjusted to (1–5)×106/mL. The culture medium was discarded by low-speed centrifugation. Then the cells were rinsed with buffer solution, and the number of cells was adjusted to about 1×106/m. Then 5 ul PI and annexin-V were added into 195 ul cell liquid, mixed well, and allowed to react for 25 minutes. The cells were taken up. Flow cytometry detected the apoptosis rate of endothelial cells. The cells with PI - / annexin V + were early apoptotic cells.
Western Blot
Western Blot was used to take samples with the same total protein content (40–60 ug), and 3*buffer was added, respectively. After boiling at 95°C for three minutes in a water bath, the samples were prepared for loading. In the pre-prepared SDS polyacrylamide, electrophoresis was continued until the protein and dye reached the bottom of SDS polyacrylamide and kept overnight at 4°C and 25V. The transfer membrane was placed in a TBS-T solution containing 5% skim milk powder and sealed for two hours at room temperature. TBS-T liquid was used to wash the film with shock for 15 minutes each time. After four washes, the corresponding primary antibody was added and incubated overnight. The horseradish peroxidase-labeled secondary antibody (1:1000) was added to the solution of TBS-T and then incubated with the transfer membrane at room temperature for one hour. The TBS-T liquid film was washed three times. Finally, ECL chemiluminescence was used in the darkroom for imaging, photographing, and scanning.
Statistical methods
Data are expressed as mean±standard deviation (SD). The data from more than two groups were evaluated by 1-way ANOVA, followed by the Newman–Keuls test. P < 0.05 was considered a significant difference.
Results
Effect of autophagy induced by TNF- on endothelial cell apoptosis detected by flow cytometry
Figure 1 shows cell apoptosis detected by flow cytometry. The apoptosis cells in the TNF-α/ActD group were significantly higher than the control group. Compared to the TNF-α/ActD group, apoptosis cells in the EET group decreased significantly (P < 0.05). Compared to the EET group, apoptosis cells in the EET+HMR-1098 group increased significantly (P < 0.01).
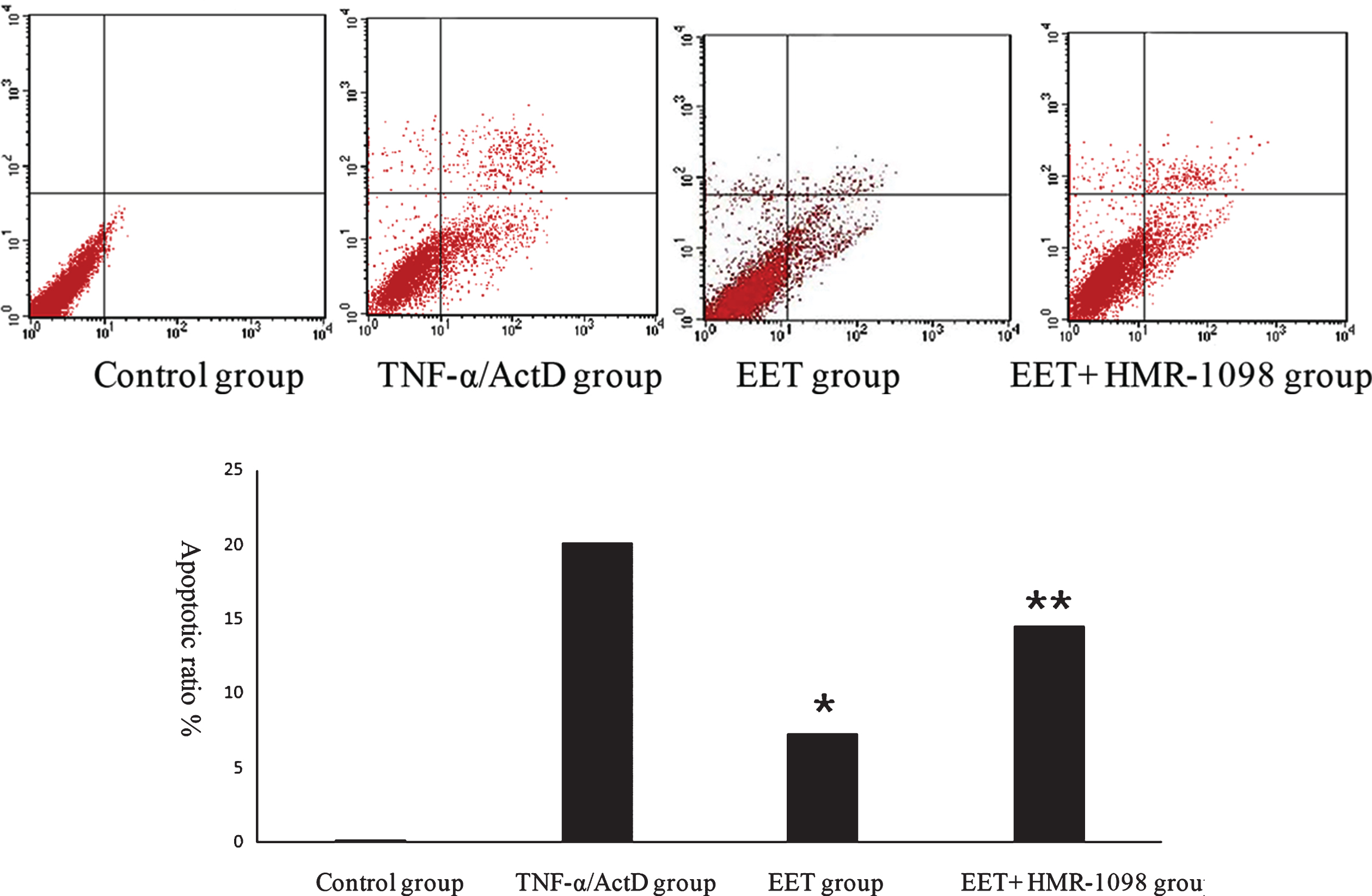
Apoptosis rate was analyzed by flow cytometry after TNF-α /ActD induced apoptosis. A to D: Typical picture of flow cytometry analysis of apoptosis rate in four groups. A: Control group. B: TNF-α/ActD group. C: EET group. D: EET+HMR-1098 group. E: Quantitate results of A to D. (* Denotes comparison between TNF-αgroup and EET group P < 0.05,** comparison between EET group and EET+HMR-1098 group P < 0.05).
Figure 2 shows the Western Blot result of LC3 and AMPK. Figure 3B shows that compared with the control group, none of the proteins’ expressions changed significantly in the TNF-α/ActD group. In the EET group, compared with the TNF-α/ActD group, LC3-II and p-AMPK increased significantly (P < 0.05), while LC3-I decreased significantly (P < 0.05). Compared with the EET group, in the EET+HMR-1098 group, LC3-II and p-AMPK decreased significantly (P < 0.01), while LC3-I increased significantly (P < 0.01). However, the total AMPK did not change in any of the groups.

A: Typical illustration of Western Blot results, p-AMPK is the phosphorylation level of AMPK, and z-AMPK is the total AMPK. B: Shows the results of three experiments of density scanning analysis. (* Denotes the comparison between TNF-α group and EET group P < 0.05, ** the comparison between EET group and EET+HMR-1098 group P < 0.05).
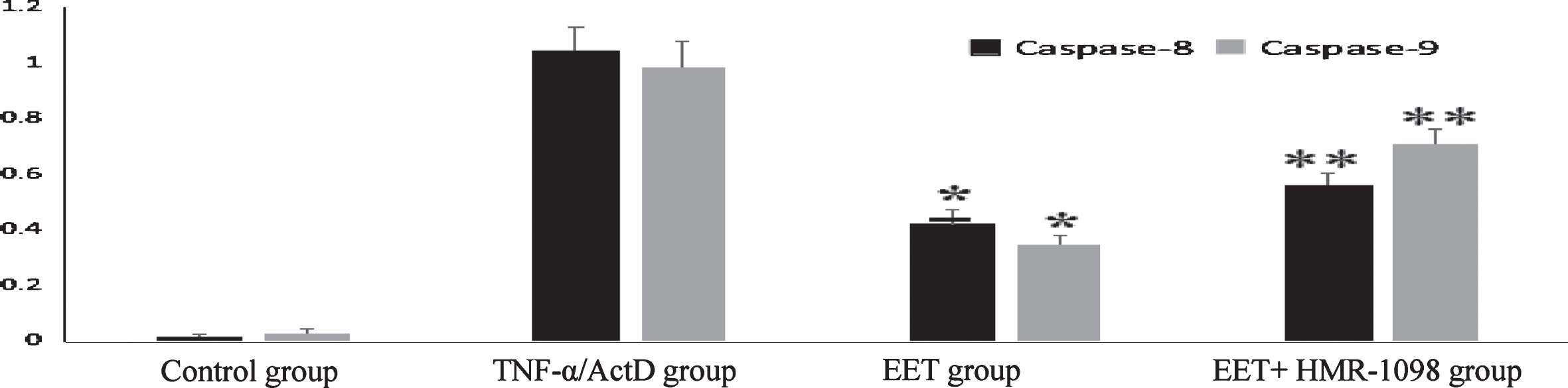
Activity results of Caspase-8 and Caspase-9.
The results showed that the activity of Caspase-8 and -9 increased after induction of TNF- /ActD and decreased significantly after the addition of 14, 15-EET (P < 0.05), but increased again after the addition of HMR-1098 (P < 0.01).
Discussion
The relationship between autophagy and apoptosis has been a popular topic in recent years. It is known that endothelial cell apoptosis and autophagy play an important regulatory role in the development of atherosclerosis [8, 9]. Jung et al. [6] found that stimulation of human umbilical vein endothelial cells with advanced glycation end products can promote the formation of autophagic bodies and the transformation of IC3-II, thus playing a protective role. Guo et al. found that curcumin can activate autophagy by inhibiting the PI3K-AktmTOR signaling pathway, protect HUVEC cells from oxidative stress damage, and inhibit their apoptosis [7].
Our earlier experiments had confirmed that EET can protect the endothelium and that autophagy plays a role in the anti-apoptosis of endothelial cells, but the specific mechanism is not clear [10]. Studying the autophagy of endothelial cells helps to understand the specific protective mechanism of EET.
AMPK is a protein kinase that senses the energy state in cells and regulates metabolism, playing an important role in the regulation of autophagy [11]. As a special ion channel, the pmKATP channel is characterized by significant channel activity inhibition with the increase of intracellular ATP concentration [12–14]. Samokhvalov et al. showed that EETs could regulate the activity of AMPK and play a role in anti-starvation-induced endothelial cell apoptosis by activating the hunger-induced pmKATP channel in endothelial cells [15]. The above experiments confirmed that arachidonic acid plays an important role in regulating autophagy and apoptosis in endothelial cells by activating the pmKATP-AMPK-mTOR signaling regulation pathway. HMR-1098 is a pmKATP-specific blocker that can block pmKATP channels in endothelial cells.
Current studies have shown that apoptosis and autophagy are interrelated and mutually restricted. The interaction between the Caspase family has attracted more and more attention. Tang et al. [16] clarified that Caspase-8 is an important pathway for regulating apoptosis and autophagy. Fang et al. [17] found that Beclin-1, a key protein in autophagy, can promote apoptosis by enhancing the activity of Caspase-9, suggesting that Caspase may be an important link between autophagy and apoptosis. In this study, the activity of Caspase-8 and Caspase-9 increased significantly under TNF-α induced endothelial cell apoptosis. However, the activity of Caspase significantly decreased after the addition of 14, 15-EET. This confirmed that the activation of Caspase was an important link in the anti-apoptosis of endothelial cells of 14, 15-EET. The addition of EET and the administration of HMR-1098 inhibited the AMPK pathway, and the activity of the Caspase family significantly increased, suggesting that 14, 15-EET may activate the AMPK pathway, inhibit the activity of the Caspase family, upregulate autophagy, and play an anti-apoptotic role.
In the study of autophagy, ATG8 has attracted extensive attention. In yeast, ATG8 is coupled with PE to form ATG8-PE [18], which is a necessary element for autophagy. LC3 is a homolog of ATG8-PE conjugate body in mammals. LC3 is involved in the formation of the autophagosome membrane, which can be cleated into LC3-I and LC3-II [19]. The results of this study showed that the LC3-II/LC3-I ratio increased during TNF-α induced endothelial cell apoptosis, suggesting that autophagy of endothelial cells was activated during TNF-α-induced endothelial cell apoptosis. After the addition of 14, 15-EET, the LC3-II/LC3-I ratio increased significantly, which confirmed that 14, 15-EET could activate the autophagy pathway and play an anti-apoptotic role. Endothelial cells treated with HMR-1098 and 14, 15-EET decreased significantly, suggesting that 14, 15-EET induction may be involved in anti-apoptotic effects through potassium channel pmKATP. Because AMPK is a protein kinase in cells that senses the energy state and regulates metabolism, it works by increasing phosphorylation levels. In this experiment, the phosphorylation level of AMPK and the total AMPK levels were detected by Western Blot. The results showed that 14, 15-EET significantly upregulated the phosphorylation of AMPK without increasing the total AMPK level. This indicates that 14, 15-EET exerts autophagy by increasing the phosphorylation of AMPK. However, after the addition of HMR-1098, the phosphorylation of AMPK was significantly reduced, and the expression of LC3-II was significantly inhibited. This suggested that 14, 15-EET upregulated autophagy by activating pmKATP and AMPK, thus playing an anti-apoptotic role.
Conclusions
Arachidonic acid can inhibit TNF-α induced endothelial cell apoptosis by upregulating autophagy. This may be achieved by activating the pmKATP channel and activating AMPK, inhibiting the activity of the Caspase family, upregulating autophagy, and thus playing an anti-apoptotic role. In this study, the mechanism of arachidonic acid in regulating endothelial cell function, protecting endothelial cells, and maintaining vascular stability was formulated theoretically. To further understand the occurrence and development of diseases, especially cardiovascular diseases, to provide a new theoretical basis for the diagnosis and treatment of these diseases.
Funding
This study was supported by Hainan Natural Science Foundation (No. 817321).
Conflict of interest
The authors declare that there is no conflict of interest.