
Abstract
BACKGROUND:
Protective effects of Ruscus extract on macromolecular permeability depend on its capacity to stimulate muscarinic receptors on endothelial cells and induce the release of endothelium derived relaxing factors (EDRFs).
OBJECTIVE:
To investigate if these effects depend only on activation of muscarinic receptors or if EDRFs release are also necessary. We have also investigated the participation of Ruscus extract on muscarinic-induced release of EDRFs on microvascular diameters.
METHODS:
Hamsters were treated daily during two weeks with Ruscus extract (50, 150 and 450 mg/kg/day) and then macromolecular permeability induced by histamine and arteriolar and venular diameters after cyclooxygenase (COX) and nitric oxide synthase (NOS) inhibitors: indomethacin and Nω-Nitro-L-arginine (LNA), respectively applied topically at 10–8M, 10–6M and 10–4M were observed on the cheek pouch preparation.
RESULTS:
Ruscus extract decreased macromolecular permeability in a dose-dependent fashion and did not affect microvascular diameters. NOS and COX inhibitors enhanced its effect on microvascular permeability. NOS inhibition reduced arteriolar diameter and COX blocking decreased arteriolar and venular diameters at the lowest dose and increased them at higher doses of Ruscus extract.
CONCLUSION:
The protective effect of Ruscus extract on macromolecular permeability seems to be mediated only via muscarinic receptors. Muscarinic activation attenuated vasoconstrictive tone through cyclooxygenase-independent endothelium derived relaxing factors.
Keywords
Introduction
Chronic venous disease (CVD) is one of the most prevalent diseases worldwide, affecting approximately 1%of the general population [1] and encompasses a large spectrum of signs and symptoms that varies from asymptomatic telangiectasias to painful venous ulcers.
The pathophysiology of CVD comprises several mechanisms that leads to venous hypertension, inflammation, endothelial damage, tissue and venous remodeling and cutaneous modifications. The severity of CVD is closely related to venous hypertension magnitude [1]. The transmission of high venous pressure to dermal microcirculation induces plasma and red blood cells extravasation to interstitium [2, 3]. Plasma leakage induces lymphatic overload, edema, blood hyperviscosity and red blood cell aggregation, that reduce their distribution in the microcirculation [4] leading to poor tissue oxygenation [5]. Red blood cell degradation products and plasma proteins that leaks to interstitium are potent leukocyte chemotactic factors and trigger an inflammatory response [6]. Leukocytes infiltrate the microcirculation by capillary plugging or by adhesion to post-capillary venules [7]. Leukocyte trapping in the microcirculation induces capillary hypoperfusion, reactive oxygen production (ROS) and proteolytic enzymes release which enhances tissue degradation [7] and leads to skin damage and leg ulcer [5].
It has been well documented in the literature that low shear stress plays an important role in the activation of the inflammatory cascade that promotes the development of venous disturbances and also leads to upregulation of angiogenesis, reflected by increased levels of endogenous positive regulators of angiogenesis [8].
Another hallmark of varicose veins is the excessive dilatation and tortuosity. The dilatation of the varicose veins is a result of the compensatory alterations in the structure and function of the vascular wall due to high hydrostatic pressures in veins [9]. In fact, vascular smooth muscle cells from varicose veins present reduced contractility in response to direct depolarization (induced by KCl), adrenergic stimulus and endothelin [10].
Ruscus aculeatus (butcher’s broom) extract is a well-recognized venotonic drug [11]; however, its mechanism of action is not completely stablished [12]. In isolated canine saphenous veins, Ruscus extract causes significant increase in tension [13, 14] due to post-junctional α1 and α2-adrenergic receptor stimulation of the vascular smooth muscle cells and release of noradrenaline stored in adrenergic nerve endings [15, 16]. Moreover, it was demonstrated that Ruscus extract has anti-inflammatory and antioxidant properties [17]. Using the cheek pouch preparation Bouskela and coworkers [18] demonstrated that Ruscus extract reduced leukotriene B4, histamine and bradykinin induced increase in microvascular permeability. More recently, it was shown that this protective effect on endothelial cell barrier function is mediated by endothelial cell muscarinic receptors stimulation [12].
In endothelial cells, type 3 muscarinic receptors (M3), when activated by an agonist (e.g. acetylcholine) induce the release of endothelial derived relaxing factors (EDRFs) such as nitric oxide (NO) and prostacyclin (PGI2). In order to investigate if the beneficial effects of Ruscus extract on microvascular permeability are due to EDRFs release and to evaluate the contribution of EDRFs, released by Ruscus extract stimulation of M3 receptors, on arteriolar and venular diameters, animals were treated daily with Ruscus 50, 150, 450 mg/kg/day during two weeks, and received topical applications of Nω-Nitro-L-arginine (L-NA) and indomethacin, the non-selective nitric oxide synthase (NOS) and cyclooxygenase (COX) inhibitors, respectively on the cheek pouch preparation.
Methods
Ruscus extraction
The dry extract of Ruscus titrated in sterolic heterosides was obtained from Ruscus aculeatus L. rhizomes and roots by hydroalcoholic extraction (Pierre Fabre Laboratories).
Animal procedures
The experimental protocol and all animal procedures were approved by the Ethical Committee of the State University of Rio de Janeiro, Brazil (registered with the number of 021/2017 and approved on March 28, 2017) in accordance to Guide for the Care and Use of Laboratory Animals [19].
Experiments were performed on 96 male Syrian hamsters (Mesocricetus auratus, Anilab, Paulinia, SP, Brazil) weighing 120–171 g (7 to 10 weeks old). All animals were fed with chow for small rodents commercially available (Nuvilab, Nuvital, Curitiba, Paraná, Brazil) and received tap water ad libitum.
Animals were distributed into four groups and received freshly prepared Ruscus extract solution in three different doses (50, 150 and 450 mg/kg/day) or filtered water (placebo group), by oral route, twice a day (at 8:00 AM and 5:00 PM), for two weeks. The last dose of Ruscus extract or filtered water was given two hours before the induction of anesthesia.
The anesthesia was induced by intraperitoneal injection of 0.1–0.2 ml of xylazine (10 mg/kg, Anasedan, Ceva Saúde Animal Ltda, Paulínia, SP, Brazil) plus ketamine (200 mg/kg, Dopalen, Ceva Saúde Animal Ltda, Paulínia, SP, Brazil) and maintained by α-chloralose (Sigma Chemicals, St. Louis, MO, USA, 100 mg/kg) administered via femoral vein. The temperature was maintained at 37.5°C throughout the surgery and experimental procedures using a heating pad controlled by a rectal thermistor. A tracheal tube was inserted to facilitate spontaneous breathing and the femoral artery was cannulated for pressure and heart rate measurements (PowerLab, AD Instruments, New South Wales, Australia).
The hamster was placed on a microscope stage similar to that described by Duling [20] with minor modifications [21]. The cheek pouch was gently everted and pinned with four to five needles into a circular well filled with silicone rubber to provide a plane bottom layer, thus avoiding stretching of the tissue but preventing shrinkage. In this position, the pouch was submerged in a superfusion solution that continuously flushed the pool of the microscope stage. Before the pouch was pinned, larger arterioles and venules were located with the aid of a Zeiss binocular stereomicroscope (Carl Zeiss Microscopy GmbH, Goettingen, Germany).
Fashioning of a single-layer preparation started with an incision of the upper layer to swing a triangular flap to one side. The exposed area was dissected at X10-16 under the stereomicroscope, and the fibrous, almost avascular, connective tissue covering the vessels was removed with ophthalmic surgical instruments. The dissected part of the pouch was 125–150μm thick. Pouches with petechial formations or those without blood flow in all vessels were discarded (five preparations were discarded due to this problem during the investigation).
The superfusion solution was a HEPES-supported HCO3-buffered saline solution (composition in mM: NaCl 110.0, KCl 4.7, CaCl2 2.0, MgSO4 1.2, NaHCO3 18.0, HEPES 15.39 and HEPES Na+ salt 14.61); temperature of the solution was maintained at 36.5°C, and superfusion rate varied from 4 ml/min (arteriolar and venular diameter measurements) to 6 ml/min (macromolecular permeability evaluation). pH was set to 7.40 by bubbling the solutions continuously with 5%CO2 in 95%N2.
For arteriolar and venular diameters and macromolecular permeability experiments, Nω-Nitro-L-arginine (L-NA, Sigma-Aldrich, St. Louis, MO, USA) and indomethacin (Sigma-Aldrich, St. Louis, MO, USA), the non-selective nitric oxide synthase (NOS) and cyclooxygenase (COX) inhibitors, respectively, were applied topically during 10 min in the following concentrations 10–8 M, 10–6 M and 10–4 M in a cumulative dose-response curve (or increasing concentrations).
Histamine (Sigma-Aldrich, St. Louis, MO, USA), and crescent concentrations of Indomethacin and L-NA were topically applied to avoid any systemic effects that would complicate the analysis of results.
For macromolecular permeability measurements, preparations were placed under an intravital microscope (Leica DMLFS, Wetzlar, Germany) equipped with a 100 W Hg DC lamp and specific filters, and allowed to rest for 30 min. After completion of the preparative procedure, fluorescein isothiocyanate-dextran (FITC-dextran, molecular weight 150,000 Dalton; Bioflor HB, Uppsala, Sweden), was given in a dose of 25 mg/100 g body weight as an i.v. injection of a 5%solution in 0.9%saline [22].
The total observed area of the pouch, 1 cm2, was roughly circular and observations of the number of leaky sites (= leaks) were made by scanning manually the total area at selected time intervals, i.e., at 2, 5, 7, 10, 15, 20 and 30 min after the initiation of each topical application of histamine: In these experiments, we made four topical applications of histamine (5μM applied for 5 min). The first one after 30 min rest of the preparation (baseline measurement), the second one after 10 min topical application of the lowest concentration (10–8 M) of either L-NA or indomethacin, applied 40 min after the injection of FITC-dextran, the third one after 10 min topical application of the middle concentration (10–6 M) of either L-NA or indomethacin and the fourth one after 10 min topical application of the highest concentration (10–4 M) of either L-NA or indomethacin. In the results. we report the number of leaks/cm2 obtained 5 min after the beginning of the topical application of 5μM of histamine because it is at this point that we observed the highest number of leaks.
For measurements of arteriolar and venular diameters, the preparations were allowed to rest for 30 min under intravital microscope. If after this time there was (a) an indication of good vascular tone; (b) brisk blood flow in all parts of the vascular bed, including the larger veins (where individual erythrocytes should not be discernible in the image of the blood stream) and (c) no tendency for leukocytes to adhere to the vessel wall, the experiments were performed by taking DVD recordings of each of three selected arterioles and venules per preparation both in initial control conditions and after each experimental intervention [21]. The microvessels were selected based on the facility to return to them at the same site, repeatedly during the experiment. The TV monitor display was used to obtain arteriolar and venular internal diameter measurements by an image shearing monitor (IPM model 907). In practice, we used DVD replay for final determination of vessel diameters since greater attention could be given to this measurement than was possible during the conduct of the experiment. In this study, the mean diameter of three arterioles and three venules per preparation were evaluated.
Statistical analysis
Results are presented as mean±SEM, unless otherwise mentioned. For intra-group comparisons Friedman’s test for nonparametric repeated measures was used followed by post-hoc Dunn’s test. Comparisons between groups were conducted by non-parametric Kruskal-Wallis test followed by Dunn’s test. Results were considered significant when P < 0.05.
Results
Effects of Ruscus extract treatment and topical application of indomethacin on macromolecular permeability induced by histamine
Topical application of crescent concentrations of indomethacin (10–8M, 10–6M and 10–4M) attenuated the macromolecular permeability increase induced by topical application of 5μM of histamine, in a dose-dependent manner. In all treatment groups, topical application of indomethacin at 10–4 M caused a significant reduction in the number of leaks compared to baseline (P < 0.001) and indomethacin at 10–8 M (P < 0.05). In addition, topical application of indomethacin at 10–6 M, significantly attenuated microvascular permeability in comparison to baseline (P < 0.05) in all groups (Fig. 1).

Effects of different doses of Ruscus extract treatment (50, 150 and 450 mg/kg/day) on microvascular permeability induced by topical application of histamine (5μM) in the presence of crescent doses of indomethacin (10–8M, 10–6M and 10–4M) applied topically on cheek pouch preparations. a) Effects of Ruscus extract treatment on microvascular permeability induced by topical application of histamine on hamster cheek pouch preparations subjected to dose-response curve of indomethacin topically applied. b) Effects of Ruscus extract treatment on microvascular permeability induced by topical application of histamine in hamster cheek pouch preparations at baseline condition. c) Effects of Ruscus extract treatment on microvascular permeability induced by topical application of histamine in hamster cheek pouch preparations during topical application of indomethacin 10–8M. d) Effects of Ruscus extract treatment on microvascular permeability induced by topical application of histamine in hamster cheek pouch preparations during topical application of indomethacin 10–6M. e) Effects of Ruscus extract treatment on microvascular permeability induced by topical application of histamine in hamster cheek pouch preparations during topical application of indomethacin 10–4M. Data were expressed as mean±SEM. *Significantly different within group at P level of 0.05. ***Significantly different within group at P level of 0.001. # Significantly different between groups at P level of 0.05. ## Significantly different between groups at P level of 0.01.
On the first topical application of 5μM of histamine (baseline), animals treated with 150 and 450 mg/kg/day of Ruscus extract presented significantly lower macromolecular permeability increase (P < 0.05 and P < 0.01, respectively) in comparison to placebo group (Fig. 1).
On the second topical application of 5μM of histamine (indomethacin 10–8 M), was observed a significant reduction of leaks in animals treated with 450 mg/kg/day of Ruscus extract in comparison to ones treated by filtered water (placebo) and Ruscus extract 50 mg/kg/day (P < 0.05 and P < 0.01, respectively) (Fig. 1).
On the third topical application of 5μM of histamine (indomethacin 10–6 M), no significant differences between treatment groups were observed (Fig. 1).
On the fourth topical application of 5μM of histamine (indomethacin 10–4 M), animals treated with 150 and 450 mg/kg/day of Ruscus extract presented significantly lower macromolecular permeability increase (P < 0.01) in comparison to placebo group (Fig. 1).
The topical application of crescent concentrations of L-NA (10–8 M, 10–6 M and 10–4 M) proportionally reduced the macromolecular permeability elevation elicited by 5μM of histamine topically applied. In all treatment groups, topical application of L-NA at 10–4 M caused a significant reduction in leaky sites compared to baseline (P < 0.001). Placebo, Ruscus extract 150 mg/kg/day and Ruscus extract 450 mg/kg/day with topical application of L-NA at 10–4 M presented significantly lower number of leaks in comparison to L-NA at 10–8 M topically applied (P < 0.05, P < 0.01 and P < 0.05, respectively) Moreover, it was observed that topical application of L-NA at 10–6 M, significantly attenuated microvascular permeability in relation to baseline in placebo and Ruscus extract 450 mg/kg/day groups (P < 0.05) (Fig. 2).
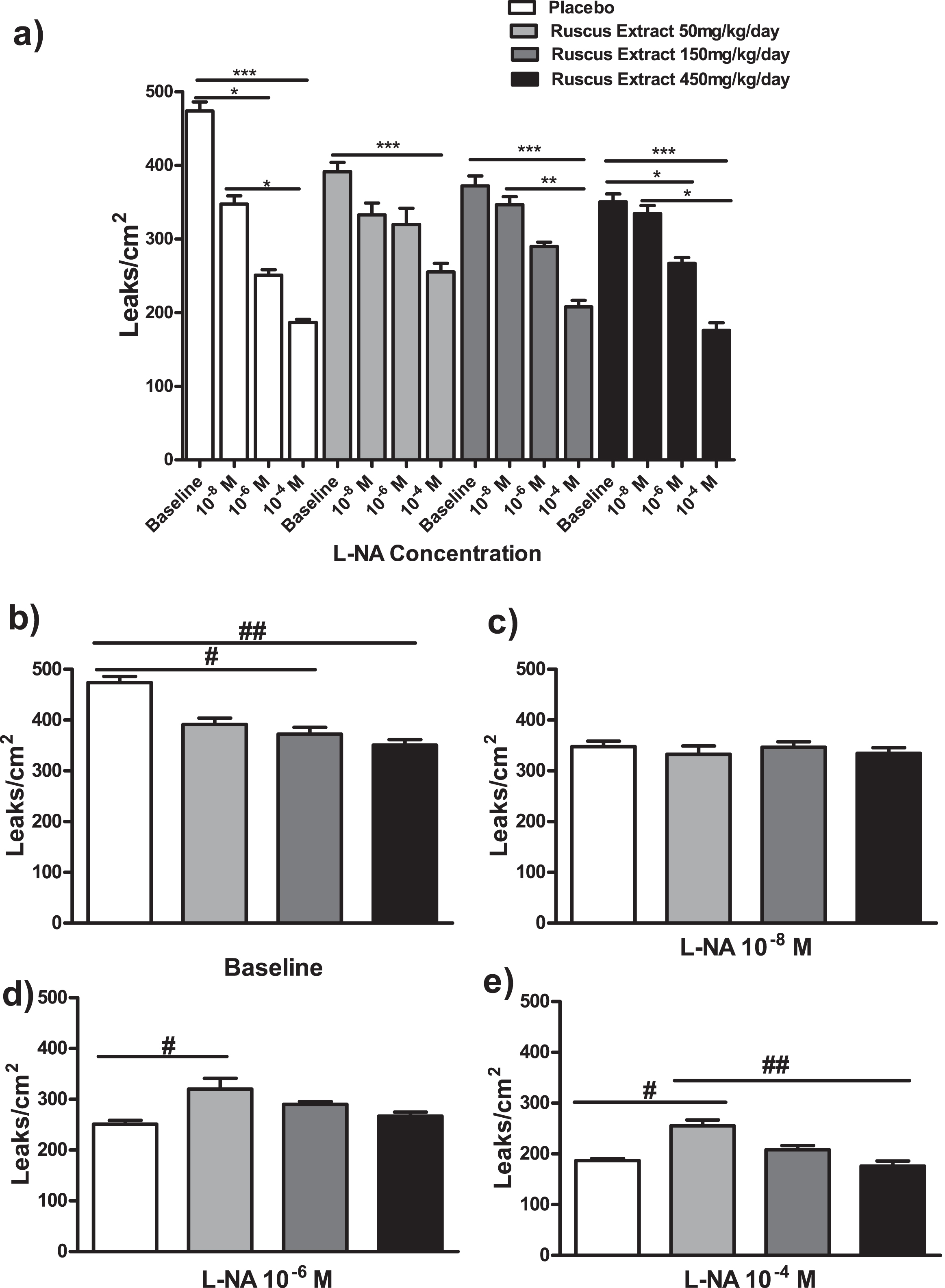
Effects of different doses of Ruscus extract treatment (50, 150 and 450 mg/kg/day) on microvascular permeability induced by topical application of histamine (5μM) in the presence of crescent doses of Nω-Nitro-L-arginine (L-NA, 10–8M, 10–6M and 10–4M) applied topically on cheek pouch preparations. a) Effects of Ruscus extract treatment on microvascular permeability induced by topical application of histamine in hamsters subjected to dose-response curve of L-NA topically applied on hamster cheek pouch preparations. b) Effects of Ruscus extract treatment on microvascular permeability induced by topical application of histamine in hamster cheek pouch preparations at baseline condition. c) Effects of Ruscus extract treatment on microvascular permeability induced by topical application of histamine in hamster cheek pouch preparations during topical application of L-NA 10–8M. d) Effects of Ruscus extract treatment on microvascular permeability induced by topical application of histamine in hamster cheek pouch preparations during topical application of L-NA 10–6M. e) Effects of Ruscus extract treatment on microvascular permeability induced by topical application of histamine in hamster cheek pouch preparations during topical application of L-NA 10–4M. Data were expressed as mean±SEM. *Significantly different within group at P level of 0.05. ***Significantly different within group at P level of 0.001. #Significantly different between group at P level of 0.05. ## Significant different between groups at P level of 0.01.
On the first topical application of 5μM of histamine (baseline), hamsters treated with 150 and 450 mg/kg/day of Ruscus extract presented significantly lower macromolecular permeability increase (P < 0.05 and P < 0.01, respectively) when compared to placebo group (Fig. 2).
On the second topical application of 5μM of histamine (L-NA 10–8 M), no significant differences between treatment groups were noticed (Fig. 2).
On the third topical application of 5μM of histamine (L-NA 10–6 M) animals treated with 50 mg/kg/day of Ruscus extract presented significantly greater number of leaky sites (P < 0.05) in comparison to placebo group (Fig. 2).
On the fourth topical application of 5μM of histamine (L-NA 10–4 M), hamsters treated with 50 mg/kg/day of Ruscus extract presented significantly higher number of leaks when compared to placebo (P < 0.05) and of Ruscus extract 450 mg/kg/day (P < 0.01) groups (Fig. 2).
The topical application of crescent concentrations of indomethacin (10–8 M, 10–6 M and 10–4 M) elicited different responses in arteriolar and venular diameters according to Ruscus extract treatment doses (Fig. 3 and Fig. 4).

Effects of different doses of Ruscus extract treatment (50, 150 and 450 mg/kg/day) on arteriolar diameter in the presence of crescent doses of indomethacin (10–8M, 10–6M and 10–4M) applied topically on hamster cheek pouch preparations. a) Effects of Ruscus extract treatment on arteriolar diameter on hamster cheek pouch preparation subjected to dose-response curve of indomethacin topically applied. b) Effects of Ruscus extract treatment on arteriolar diameter of hamster cheek pouch preparations at baseline conditions. c) Effects of Ruscus extract treatment on arteriolar diameter on cheek pouch preparations during topical application of indomethacin 10–8M. d) Effects of Ruscus extract treatment on arteriolar diameter on cheek preparations during topical application of indomethacin 10–6M. e) Effects of Ruscus extract treatment on arteriolar diameter on cheek pouch preparations during topical application of indomethacin 10–4M. Data were expressed as mean±SEM. *Significantly different within group at P level of 0.05. **Significantly different within group at P level of 0.01. ***Significantly different within group at P level of 0.001. # Significantly different between groups at P level of 0.05. ## Significantly different between groups at P level of 0.01. ### Significantly different between groups at P level of 0.001.
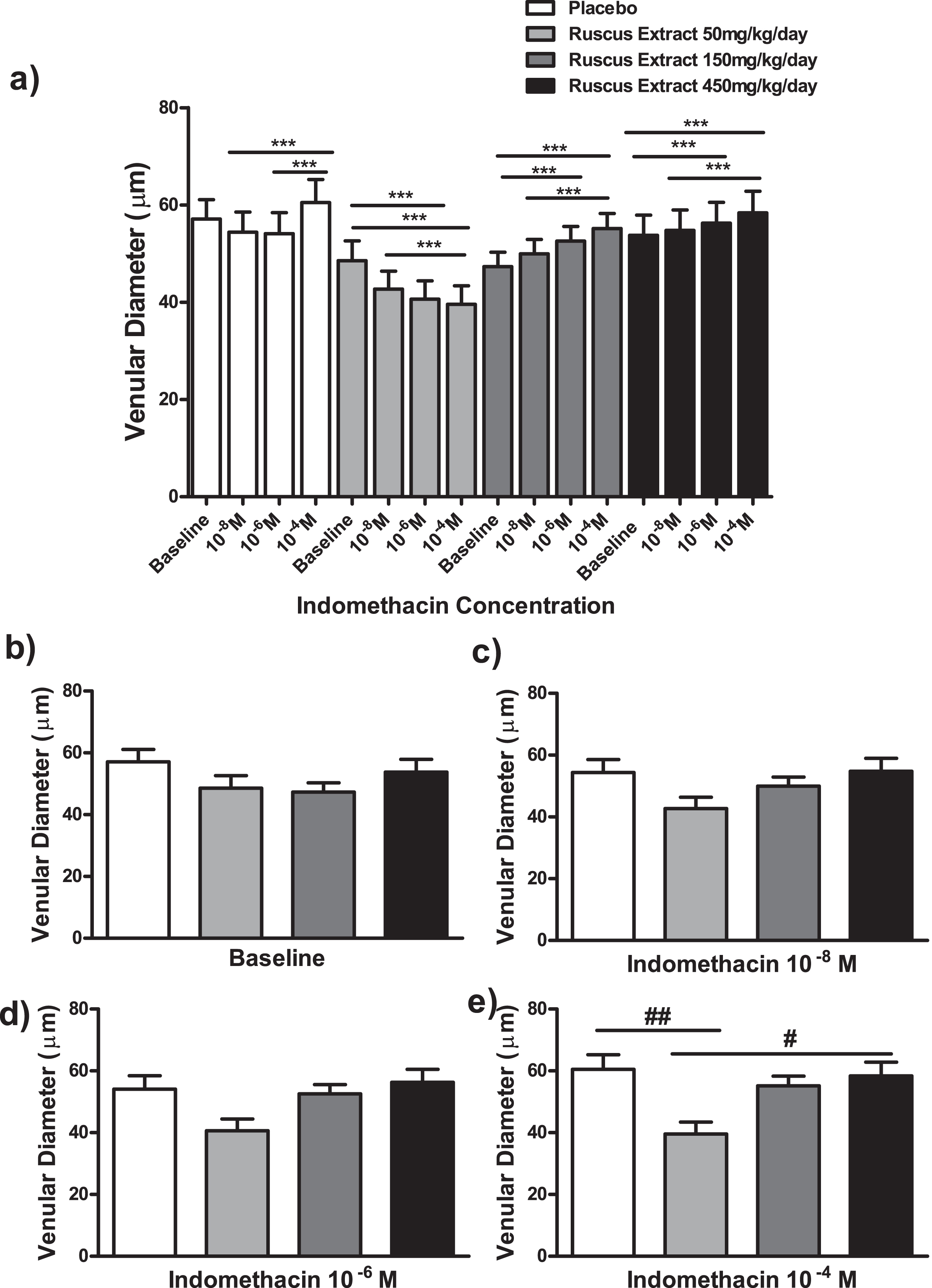
Effects of different doses of Ruscus extract treatment (50, 150 and 450 mg/kg/day) on venular diameter in the presence of crescent doses of indomethacin (10–8M, 10–6M and 10–4M) applied topically on cheek pouch preparations. a) Effects of Ruscus extract treatment on venular diameter on cheek pouch preparations subjected to dose-response curve of indomethacin topically applied. b) Effects of Ruscus extract treatment on venular diameters on cheek pouch preparations at baseline conditions. c) Effects of Ruscus extract treatment on venular diameter on cheek pouch preparations during topical application of indomethacin 10–8M. d) Effects of Ruscus extract treatment on venular diameter on cheek pouch preparations during topical application of indomethacin 10–6M. e) Effects of Ruscus extract treatment on venular diameter on cheek pouch preparations during topical application of indomethacin 10–4M. Data were expressed as mean±SEM ***Significantly different within group at P level of 0.001. # Significantly different between group at P level of 0.05. ## Significantly different between groups at P level of 0.01.
Ruscus extract treatment at 50 mg/kg/day, as expected, elicited a dose-dependent decrease in arteriolar and venular diameters. Indomethacin at 10–4 M elicited a significant reduction of arteriolar and venular diameters when compared to baseline and indomethacin at 10–8 M (P < 0.001), additionally, indomethacin at 10–6 M elicited a significant decrease in arteriolar and venular diameters in comparison to baseline (P < 0.001). Conversely, in placebo group the topical application of indomethacin at 10–4M caused a significant increase in arteriolar diameter when compared to baseline (P < 0.001) and topical applications of indomethacin at 10–6 M (P < 0.05) and 10–8 M (P < 0.01). Regarding venular diameter, indomethacin at 10–4 M elicited a significant increase in comparison to indomethacin administration at 10–6 M and at 10–8 M (P < 0.001). Moreover, in groups treated with Ruscus extract 150 and 450 mg/kg/day, indomethacin induced a dose dependent increase in arteriolar and venular diameters. Indomethacin at 10–4 M elicited a significant increase in arteriolar and venular diameters in comparison to baseline and indomethacin at 10–8 M (P < 0.001). Indomethacin at 10–6 M promoted significant increase in arteriolar and venular diameters (P < 0.001) in comparison to baseline (Fig. 3 and Fig. 4).
At baseline, no significant differences concerning arteriolar and venular diameters between groups was observed (Fig. 3 and Fig. 4).
Topical application of indomethacin at 10–8 M caused a significant decrease in arteriolar diameter in Ruscus extract 50 mg/kg/day treated group (P < 0.05) in comparison to placebo (Fig. 3).
Indomethacin at 10–6 M, topically applied, promoted a significant reduction of arteriolar diameter in Ruscus extract 50 mg/kg/day (P < 0.01) and 450 mg/kg/day (P < 0.05) treated groups when compared to placebo group (Fig. 3).
The application of indomethacin 10–4 M topically, elicited a significant reduction of arteriolar diameter in Ruscus extract 50 mg/kg/day treated group in relation to placebo (P < 0.001), Ruscus extract 150 mg/kg/day (P < 0.05) and 450 mg/kg/day (P < 0.01) treated groups (Fig. 3).
Concerning venular diameters, no significant differences were observed between groups at baseline and during topical applications of indomethacin at 10–8 M and 10–6 M (Fig. 4). During topical application of indomethacin at 10–4 M, it was observed a significant reduction of venular diameter in comparison to placebo (P < 0.01) and in comparison to Ruscus 450 mg/kg/day (P < 0.05) treated groups (Fig. 4).
Topical administration of crescent concentrations of L-NA (10–8 M, 10–6 M and 10–4 M) evoked a dose response reduction in arteriolar and venular diameters in all treatment groups, with exception of placebo group regarding venular diameter (Fig. 5 and Fig. 6).
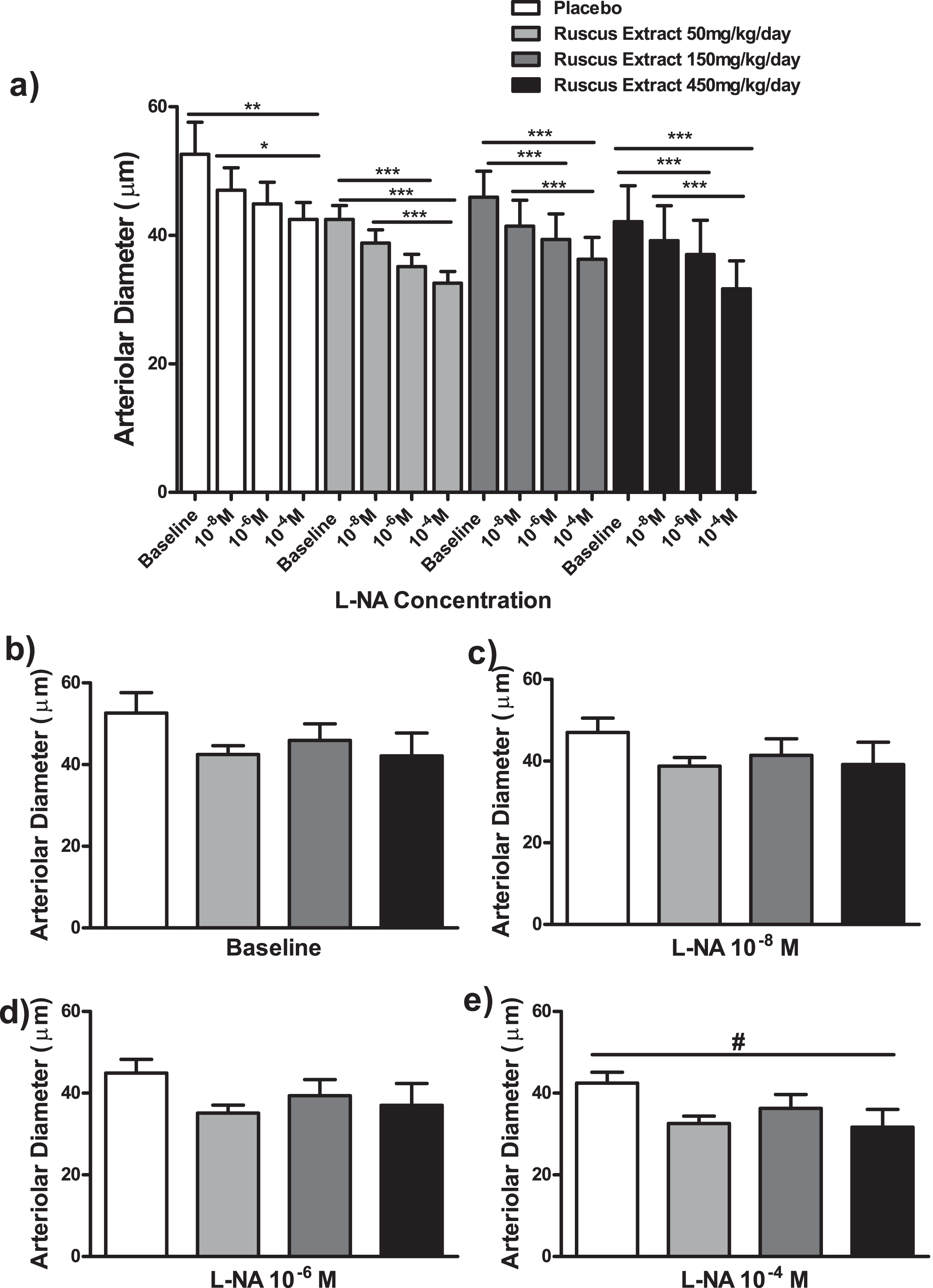
Effects of different doses of Ruscus extract treatment (50, 150 and 450 mg/kg/day) on arteriolar diameter in the presence of crescent doses of Nω-Nitro-L-arginine (L-NA, 10–8M, 10–6M and 10–4M) applied topically on cheek pouch preparations. a) Effects of Ruscus extract treatment on arteriolar diameter on cheek pouch preparations subjected to dose-response curve of L-NA topically applied. b) Effects of Ruscus extract treatment on arteriolar diameter on cheek pouch preparations at baseline conditions. c) Effects of Ruscus extract treatment on arteriolar diameter on cheek pouch preparations during topical application of L-NA 10–8M. d) Effects of Ruscus extract treatment on arteriolar diameter on cheek preparations during topical application of L-NA 10–6M. e) Effects of Ruscus extract treatment on arteriolar diameter on cheek pouch preparations during topical application of L-NA 10–4M. Data were expressed as mean±SEM. *Significantly different within group at P level of 0.05. **Significantly different within group at P level of 0.01. ***Significantly different within group at P level of 0.001. # Significantly different between groups at P level of 0.05.
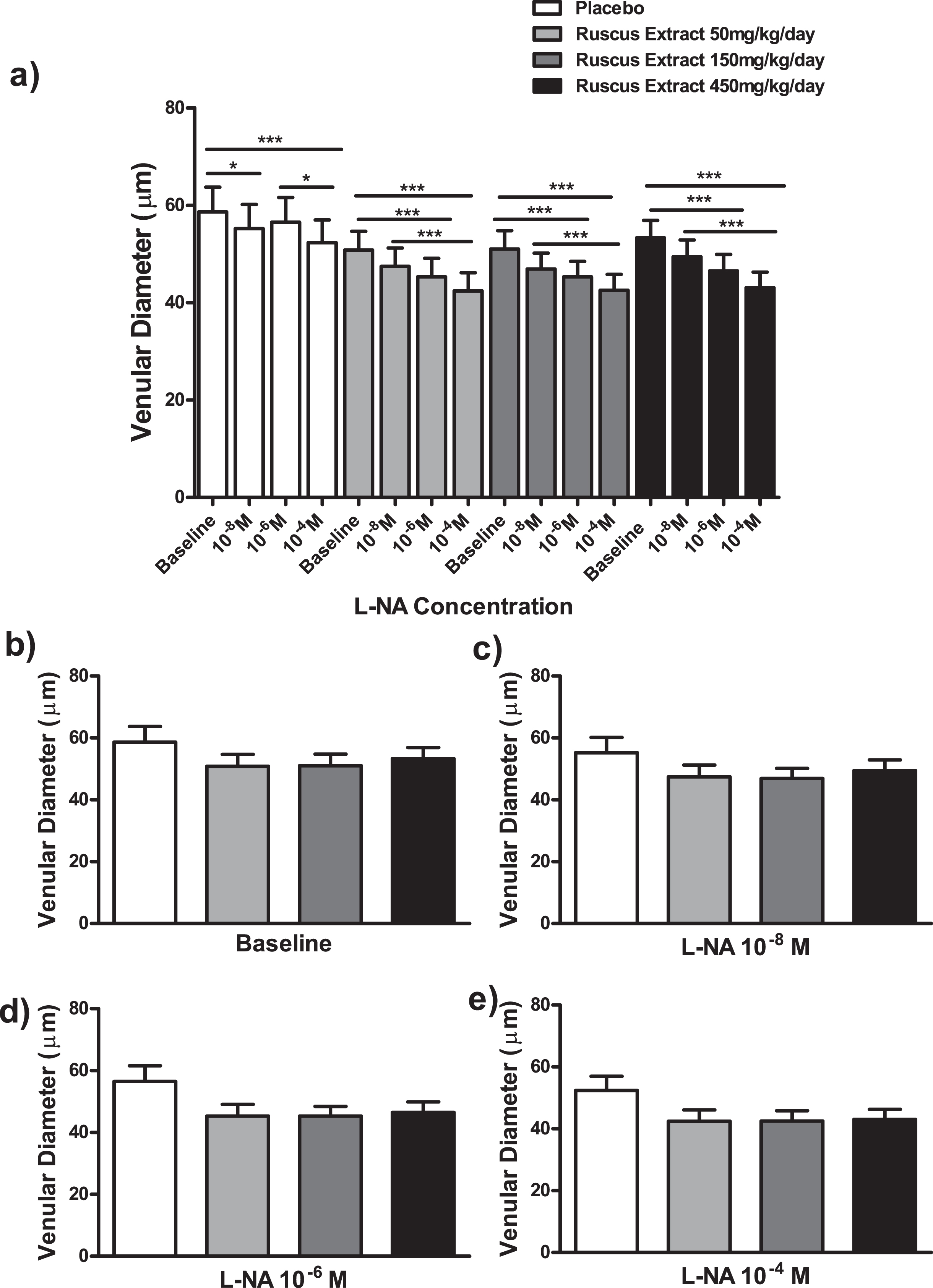
Effects of different doses of Ruscus extract treatment (50, 150 and 450 mg/kg/day) on venular diameter in the presence of crescent doses Nω-Nitro-L-arginine (L-NA, 10–8M, 10–6M and 10–4M) applied topically on cheek pouch preparation. a) Effects of Ruscus extract treatment on venular diameter on cheek pouch preparations subjected to dose-response curve of L-NA topically applied. b) Effects of Ruscus extract treatment on venular diameter on cheek pouch preparations at baseline condition. c) Effects of Ruscus extract treatment on venular diameter on cheek pouch preparations during topical application of L-NA 10–8M. d) Effects of Ruscus extract treatment on venular diameter on cheek preparations during topical application of L-NA 10–6M. e) Effects of Ruscus extract treatment on venular diameter on cheek pouch preparations during topical application of L-NA 10–4M. Data were expressed as mean±SEM *Significantly different within group at P level of 0.05. ***Significantly different within group at P level of 0.001.
L-NA at 10–4 M elicited a significant reduction of arteriolar diameter when compared to baseline (P < 0.01) and indomethacin at 10–8 M (P < 0.05). On the other hand, L-NA at 10–4 M elicited a significant decrease in venular diameter in comparison to baseline (P < 0.001) and L-NA 10–6 M (P < 0.05). L-NA at 10–8 M evoked a significant reduction in venular diameter in comparison to baseline (P < 0.05). In groups treated with Ruscus extract 50, 150 and 450 mg/kg/day, topical application of L-NA at 10–4 M evoked a significant reduction of arteriolar and venular diameters in comparison to baseline and 10–8 M (P < 0.001). In addition, topical application of L-NA at 10–6 M evoked a significant decrease of arteriolar and venular diameters in comparison to baseline (P < 0.001) in groups treated with Ruscus extract 50, 150 and 450 mg/kg/day (Fig. 5 and Fig. 6).
No significant differences concerning arteriolar and venular diameters between treatment groups was observed at baseline and during topical applications of LNA at 10–8 M, 10–6 M and 10–4 M (only in the case of venular diameter) (Fig. 5 and Fig. 6). The application of L-NA at 10–4 M, topically, induced a significant reduction of arteriolar diameter in Ruscus extract 450 mg/kg/day treated group (P < 0.05) in relation to placebo (Fig. 5).
The major findings of this study were: a) treatment with Ruscus extract at different concentrations (50, 150 and 450 mg/kg/day) during two weeks elicited a protective effect of the endothelial barrier function diminishing macromolecular permeability increase in post capillary venules, induced by histamine and that these effects were enhanced by topical applications of crescent concentrations of non-selective NOS and COX inhibitors; b) Different daily doses of Ruscus extract did not affect arteriolar and venular diameters. On the other hand, NOS inhibition caused a dose dependent reduction on arteriolar diameter and COX blocking elicited a biphasic response on arteriolar and venular diameters, with vasoconstriction at lowest dose and vasodilatation at higher doses of Ruscus extract treatment.
CVD is characterized by increased venous pressure that is transmitted from large veins to venules and capillaries of the skin that become chronically dilated, dense elongated and tortuous [23]. Concomitant deterioration of glycocalyx and endothelial function occur, leading to increased microvascular permeability, edema, red blood cells extravasation and leukocyte recruitment and activation [24]. In CVD, endothelial dysfunction through a plethora of different pathways triggers the production of inflammatory factors including cytokines, chemotactic factors, proteases that aggravate and, ultimately, perpetuate the inflammatory process [25, 26]. Inflammatory stimuli, elicits an impairment of endothelial cell barrier, which leads to an increase in permeability and edema formation [27]. Histamine induced a rapid transient increase in venular permeability as consequence of endothelial cell gap formation [28]. These gaps’ formation is mainly attributed to endothelial cell contraction, a phenomenon that involves actin and nonmuscle myosin interaction, and that is dependent of Ca2 +, calmodulin and ATP and the phosphorylation of myosin light chain (MLC) by MLC kinase (MLCK) [29–31]. More recently, Adderley and coworkers [32] demonstrated that Histamine, via H1 receptor, increased microvascular permeability by MLCK, RhoA/Rho Kinase (ROCK) and p38 MAP (mitogen activating protein) kinase activation. Moreover, the stimulation of H1-receptor leads to tyrosine phosphorylation of VE-cadherin, which induced its dissociation from β-cathenin and its sequestration from adherens junctions [32–35].
Previous study has demonstrated that Ruscus extract presented an inhibitory effect on increased microvascular permeability induced by histamine in hamster cheek pouch preparation [34]. Recently, Rauly-Lestienne and coworkers [12] demonstrated that this protective effect is due, in part, to muscarinic receptor stimulation, particularly M1 and M3 subtypes. In fact, activation of M3 receptor has been pointed out as an important requirement for the maintenance of endothelial barrier function and adherens junctions’ integrity [37, 38]. Since Ruscus extract has the property to activate endothelial M3 receptors which are responsible to induce NO and prostanoids synthesis (especially PGI2), we hypothesized that they were involved in the mechanism by which Ruscus extract-activated M3 receptors diminishes microvascular permeability increase induced by histamine. Crescent concentrations of indomethacin, a non-selective COX inhibitor, caused a proportional and significant decrease in microvascular permeability in all groups, but this protective effect was more pronounced in animals treated with higher doses of Ruscus extract (150 and 450 mg/kg/day) in comparison to placebo group. The main prostanoid synthesized by endothelial cells is PGI2, which augments cytosolic cAMP, well recognized factor that strengthens barrier function both in vitro and in vivo [39–44] and activates two types of effector molecules: Protein kinase A (PKA) and exchange protein directly activated by cAMP (Epac). Epac1, but not PKA, stimulates the formation of cell junctions, thereby increasing the barrier function and protection against various inflammatory mediators [44]. We have supposed that, if diminishing cAMP intracellular levels, microvascular permeability would increase, but we observed the opposite effect. We believe that Ruscus extract exert its protective effects on endothelial cell barrier by means of M3 receptor activation. On the other hand, this protection is independent of COX pathway. It is possible that indomethacin reduced ROS production via COX inhibition [45], and this effect was potentialized by the antioxidant property of Ruscus extract [17].

Schematic representation of Ruscus extract protective mechanism on microvascular permeability. Ruscus extract activates type 3 muscarinic receptor (M3) on endothelial cells surface, which confers the maintenance of endothelial barrier function and adheres junction’s integrity. After topical application of histamine, Ruscus extract, via M3 stimulation, prevents microvascular permeability increase. Topical administration of NO and prostacyclin (PGI2) synthesis inhibitors, reduces macromolecular permeability, suggesting that the protective effects of Ruscus extract on endothelial barrier function and intercellular junctions is dependent of M3 activation, but is not dependent of NO and PGI2.
Several studies have demonstrated that inhibition of NO synthesis reduces the increase of microvascular permeability [46–48] and has been considered a key mediator of macromolecular permeability, by eNOS-NO-sGC-PKG, the classical pathway and by the emerging alternative S-Nitrosation pathway [49]. The extracellular regulated kinase-1/2 (ERK-1/2) activated by the conventional pathway phosphorylates the adherens junctions’ proteins, disrupting the endothelial cell barrier. The S-Nitrosation, i.e. the covalent modification of thiol groups of cysteine residues by NO, of junctional proteins (β-catenin and p120 catenin) causes their internalization and disassembly of junctional complex. Both classical and emerging eNOS-NO pathways act independently or in a parallel way to increase microvascular permeability after inflammatory stimuli. Similarly, in macromolecular permeability experiments with topical application of indomethacin, the experiments of permeability with topical application of L-NA also have shown, at baseline, that Ruscus extract activation of M3 receptors per se prevented the endothelial cell barrier disruption after histamine administration. Additionally, the inhibition of NO synthesis by crescent L-NA concentrations, gradually diminished number of leaky sites, including in placebo group. It seems that crescent concentrations of L-NA proportionally inhibited histamine induced increase on macromolecular permeability in all groups (including placebo) and this inhibition became significant at greater daily doses of Ruscus extract (150 and 450 mg/kg/day). During topical applications of L-NA 10–6 M and 10–4 M, the group treated with Ruscus 50 mg/kg/day presented a significant increase in number of leaks in comparison to placebo and the group treated with Ruscus extract 450 mg/kg/day. It seems that NO production exceeded its inhibition elicited by L-NA. The possible mechanism by which Ruscus extract protects endothelial barrier function and maintains intercellular junctions’ integrity is depicted on Fig.7.
Concerning arteriolar and venular diameters, topical application of indomethacin elicited a biphasic response in animals treated with Ruscus extract. The group treated with the lowest daily dose presented gradual vasoconstriction, that became significant at higher concentrations of indomethacin. It is known that Ruscus extract induces a dose-dependent increase of contractile activity in veins [13, 14] that is due to a direct effect on post junctional α1 - and α2-adrenergic receptors in smooth muscle cells and to noradrenaline release from adrenergic nerve endings [15, 16]. In endothelium, it was demonstrated that α2-adrenergic agonists are capable to induce endothelium-dependent relaxation in a number of isolated blood vessels [50]; however, Usui and coworkers [51], observed that vasoconstriction induced by noradrenaline was due to α2- adrenergic stimulation in endothelial cells from canine basilar arteries, and that this effect was inhibited by acetylsalicylic acid, a non-selective COX blocker, evidencing the release of COX-dependent vasoconstrictors in that preparation. We believe that in the group treated with Ruscus extract, 50 mg/kg/day, there was a predominance of α1 - and α2- adrenergic stimuli in smooth muscle cells in relation to α2- adrenergic stimulation in endothelial cells, resulting in increased arteriolar and venular tone. In the presence of crescent concentrations of indomethacin, occurred a dose-dependent decrease of arteriolar and venular diameters, probably due to inhibition of COX-derived vasodilators, mainly PGI2, as consequence of M3 receptor activation. On the other hand, in animals treated with 150 and 450 mg/kg/day, topical application of indomethacin promoted an unexpected significant increase in arteriolar and venular diameters. One possible explanation, for these unexpected observations, is the fact that Ruscus extract at highest doses release greater amounts of noradrenaline which stimulate α2-adrenergic receptors in endothelial cells resulting in COX-derived vasoconstrictor release. Indomethacin, inhibited COX-derived vasoconstrictor release and ultimately elicits an increase of arteriolar and venular diameter proportional to indomethacin concentrations. Moreover, it is feasible that M3 receptor stimulation promoted the release of COX-independent EDRFs causing additional vasodilatation in these groups.
Regarding arteriolar and venular diameters, crescent concentrations of L-NA induced a dose dependent vasoconstriction in all groups. It seems that in Ruscus extract treated groups, EDRFs release, induced by M3 receptor stimulation, counteracted the vasoconstriction induced by α1 - and α2-adrenergic receptors stimulation. However, in the presence of crescent concentrations of L-NA occurred dose-dependent vasoconstriction due to decreased levels of NO.
In conclusion, this work evidenced that the protective effect of Ruscus extract on endothelial cell barrier function via muscarinc receptor activation is not dependent of prostanoids and nitric oxide and suggests that muscarinic activation attenuated the vasoconstriction induced by α1 and α2-adrenergic receptors stimulation by means of cyclooxygenase-independent endothelium derived relaxing factors, mainly nitric oxide.
Conflict of interest
No conflict of interest to declare.
Funding
This work was funded by an Investigator proposed research project to Pierre Fabre’s Laboratory.
Footnotes
Acknowledgments
Authors would like to thank the assistance of Mr. Paulo José Ferreira Lopes and Mr. Claudio Natalino Ribeiro for their care of the animals.