
Abstract
BACKGROUND:
Sepsis is the result of a dysregulated host immune response to an infection. An ideal therapy would target both the underlying infection and the dysregulated immune response. DIBI, a novel iron-binding polymer, was specifically developed as an antimicrobial agent and has also demonstrated in vivo anti-inflammatory properties.
OBJECTIVE:
This study aimed to further investigate the effects of DIBI with and without the antibiotic imipenem (IMI) in colon ascendens stent peritonitis (CASP)-induced experimental sepsis.
METHODS:
Vehicle, DIBI and/or IMI were administered in C57BL/6 mice after CASP surgery. Intestinal leukocyte activation and capillary perfusion was evaluated by intravital microscopy. Moreover, bacterial load in peritoneal lavage fluid and blood, and plasma cytokine levels were assessed. In a second series of experiments, surgery to repair the colon was performed at 5 hr and these mice were followed for long-term survival over 7 days.
RESULTS:
DIBI reduced leukocyte adhesion, improved capillary blood flow, and decreased key plasma cytokines levels. DIBI also improved survival of infected mice and greatly improved IMI efficacy. Survivors treated with IMI and DIBI were found to be free of systemic infection.
CONCLUSIONS:
DIBI has promising potential for sepsis treatment including its use as a sole or an adjunct therapeutic with antibiotics.
Introduction
The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3) defined sepsis as a life-threatening organ dysfunction due to the dysregulated host immune response to infection [1]. In North America as in the rest of the world, sepsis is a leading cause of death in surgical Intensive Care Units [2–7] affecting millions of people every year. The prevalence and the incidence of this condition is also rising by approximatively 9% every year in the USA, for example [8], with a corresponding increased financial burden on the health care system estimated at $15 billion/year in the USA alone [9–13].
During sepsis, both the initial infection and the ensuing dysregulated immune response are responsible for this life-threatening pathogenesis [14]. Sepsis is a complex clinical syndrome associated with a variety of different infections and despite various advances in medicine, there is still no specific treatment for the immune dysregulation occurring with this disease. The main goal of sepsis management is to first early diagnose and then control the source of infection, i.e., surgically and/or by using appropriate antimicrobial therapy, in a time-sensitive manner. Early detection of sepsis with prompt initiation of treatment substantially increases patient survival [15–18]. According to the 2016 Surviving Sepsis Campaign guidelines, initial treatment should include a broad-spectrum antibiotic such as imipenem (IMI) that has activity against various likely pathogens. This empiric antibiotic therapy is followed by more specific antibiotics once the specific infectious agent is identified from blood culture and has been tested for its antibiotic sensitivity profile [19]. There are other supportive therapies used to manage septic patients including intensive fluid resuscitation and vasopressors to control blood pressure [19–21].
Given the overall pathogenesis of sepsis, an ideal therapy would target both the infection and the dysregulated immune response. In this regard, iron sequestering agents generally referred to as iron chelators have an interesting therapeutic potential. Iron is an essential trace element required for the growth and replication of almost all microbial sepsis-causing pathogens. Iron also serves as the chemical catalyst for the Fenton reaction that produces reactive oxygen species (ROS) - mediators of the septic dysregulated immune response [2, 22]. However, under physiological conditions, ROS are also beneficially involved in pathogen degradation during phagocytosis, i.e., intra-phagosomal bacterial killing [23, 24]. Thus, the role of iron in sepsis can be regarded as a double-edged sword: bacteria utilize it as an essential nutrient for their growth during infection while leukocytes produce ROS through the iron-catalyzed chemical reaction to degrade bacteria [25–29]. Nonetheless, inflammatory dysregulation in sepsis stems from iron’s excess inflammatory promoting properties leading to (i) increased leukocyte-endothelial interaction, (ii) impaired capillary blood flow, and (iii) excessive release of plasma cytokines known as the “cytokine storm”. The appropriate reduction of iron-related ROS production can therefore be beneficial in sepsis. During infection, the host initiates a hypoferremic response that promotes intracellular iron retention to reduce its bioavailability to pathogens [29]. Iron chelators, molecules that bind iron with high affinity, could be used to mimic/facilitate this hypoferremic response. Correspondingly, some studies have shown improvements in experimental models of inflammation and sepsis using the FDA approved iron chelators (deferoxamine (DFO), deferiprone (DFP), and deferasirox (DFX)) [30–33]. However, these conventional clinical chelators were developed to treat hematological disorders as associated with pathologically greatly elevated body iron stores and not surprisingly have shown serious limitations for treating infection. Importantly, these hematological chelators either carry black box warnings as to their toxicities [34] or have been found to actually promote fungal [35] or bacterial infection [36].
DIBI, is a novel iron-binding polymer that was specifically developed as an antimicrobial agent. The molecule has a polyvinylpyrrolidone backbone containing nine 3-hydroxy-1-(β-methacrylamidoethyl)-2-methyl-4(1H) pyridinone (MAHMP) residues per molecule [37] as shown in Fig. 1 In contrast to conventional clinical iron chelators, DIBI has higher affinity for iron, sequesters iron away from microbes and does not display host toxicity [38–41]. DIBI’s anti-inflammatory effects have also been demonstrated using different models such as local and systemic endotoxin-induced uveitis, LPS-induced local and systemic inflammation, and experimental pneumonia [42–47]. Importantly given its 9 kDa molecular size it preferentially addresses the extracellular iron pools that are typically accessed by bacterial pathogens and therefore is less likely to affect the intracellular iron pools of phagocytes, i.e., as needed to kill ingested bacteria by ROS-mediated killing mechanisms.
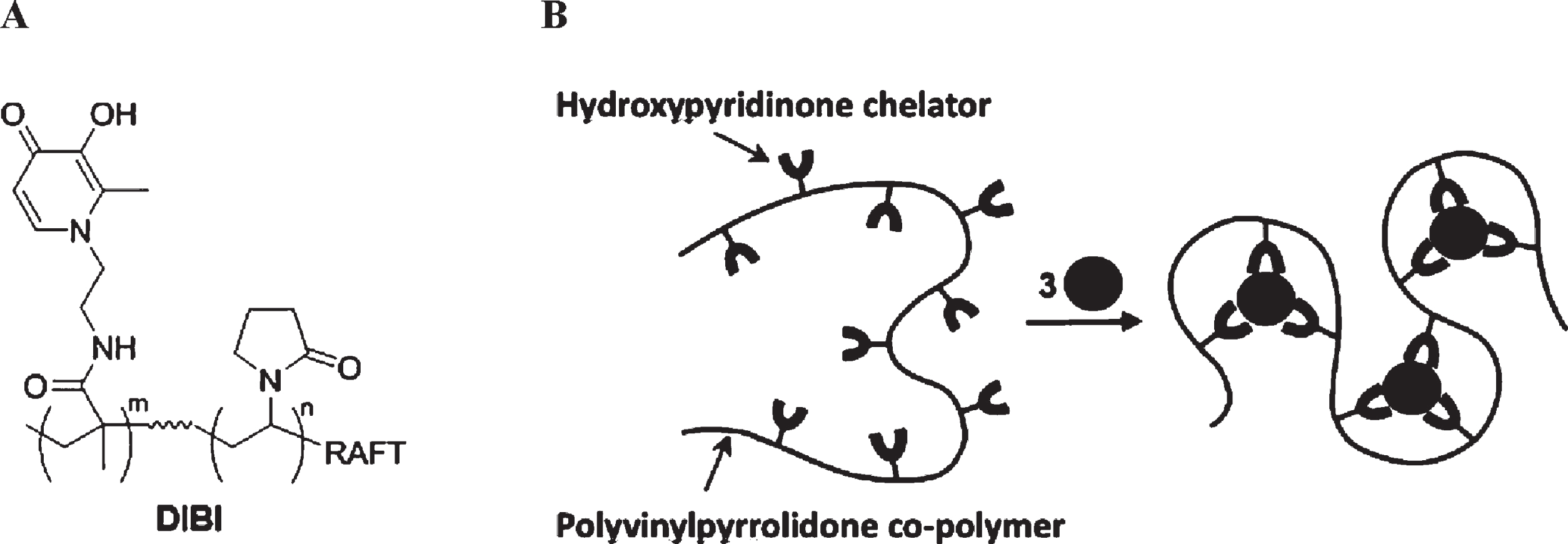
Structure of DIBI polymer. (A) chemical structure of DIBI; nine 3-hydroxy-1-(β-methacrylamidoethyl)-2-methyl-4(1 H)-pyridinone (MAHMP) residues on a PVP backbone; (B) representation of fully-coordinated hexadentate iron binding by DIBI [38].
Our previous studies had shown that DIBI can modulate infection and key aspects of sepsis-related leukocyte adhesion and cytokine release in an experimental CASP model of sepsis [48]. However, we had measured these various effects of DIBI both alone or with IMI, a full 16 hr after sepsis induction. The CASP model has the clear disadvantage of providing continuous infection from continuing fecal leakage to the peritoneal cavity [49] and therefore, this seriously complicates studies of infection, sepsis pathophysiology and therapeutic treatments.
The overall objective of the present study was to further evaluate the effects of DIBI both with and without IMI in polybacterial CASP- induced sepsis but following an early surgical intervention to repair the colon breach (CASP-I). This approach therefore included investigation of sepsis events much earlier after induction and importantly, it also provided a means to study the longer-term aspects of survival following the experimental therapeutic treatments.
Animal model and ethics statement
Male C57BL/6 mice (8–12 weeks, 20–30 g) from Charles River Laboratories International Inc. (Wilmington, MS) were used in the study. The mice were housed in ventilated cage racks in the Carleton Animal Care Facility (CACF) of Dalhousie University. Animals received standard rodent chow and water ad libitum. The University Committee on Laboratory Animals (UCLA) approved all experimental procedures, which followed the ethical guidelines of the Canadian Council on Animal Care. Ethical endpoints were in place for the duration of all experiments based on the recommendations of the UCLA.
Experimental models
CASP Model
CASP surgery was performed as previously described by Islam et al. [50] and with its experimental timelines as shown in Fig. 2. Briefly, animals were anesthetized with isoflurane and administered pre-emptive analgesia (0.1 mg/kg Buprenorphine subcutaneously (s.c.)). Anesthesia was monitored during the procedure by the pedal withdrawal reflex. A laparotomy was performed aseptically to expose the ascending colon. A 20 G 1–1/4 catheter (Jelcro, Smiths Medical, Kent, UK) was inserted to a depth of 2 mm into the wall of the ascending colon, approximately an inch distal to the ileocecal valve. The catheter was securely placed and sutured at a 45° angle from the wall of ascending colon, extending 2 mm outside of the ascending colon. The cecum was palpated to ensure fecal leakage through the stent into the peritoneum. Sham surgery was performed as above, but the catheter was sutured on the surface of the intact colon so as to preclude fecal leakage. The ascending colon was placed back into the abdominal cavity and the muscle and skin layers were closed. Treatment drugs (or saline) were given intraperitoneally (i.p.) before closing the abdomen, which represented T = 0hr. All mice were given 0.5 mL 0.9% saline (s.c) for fluid replacement.
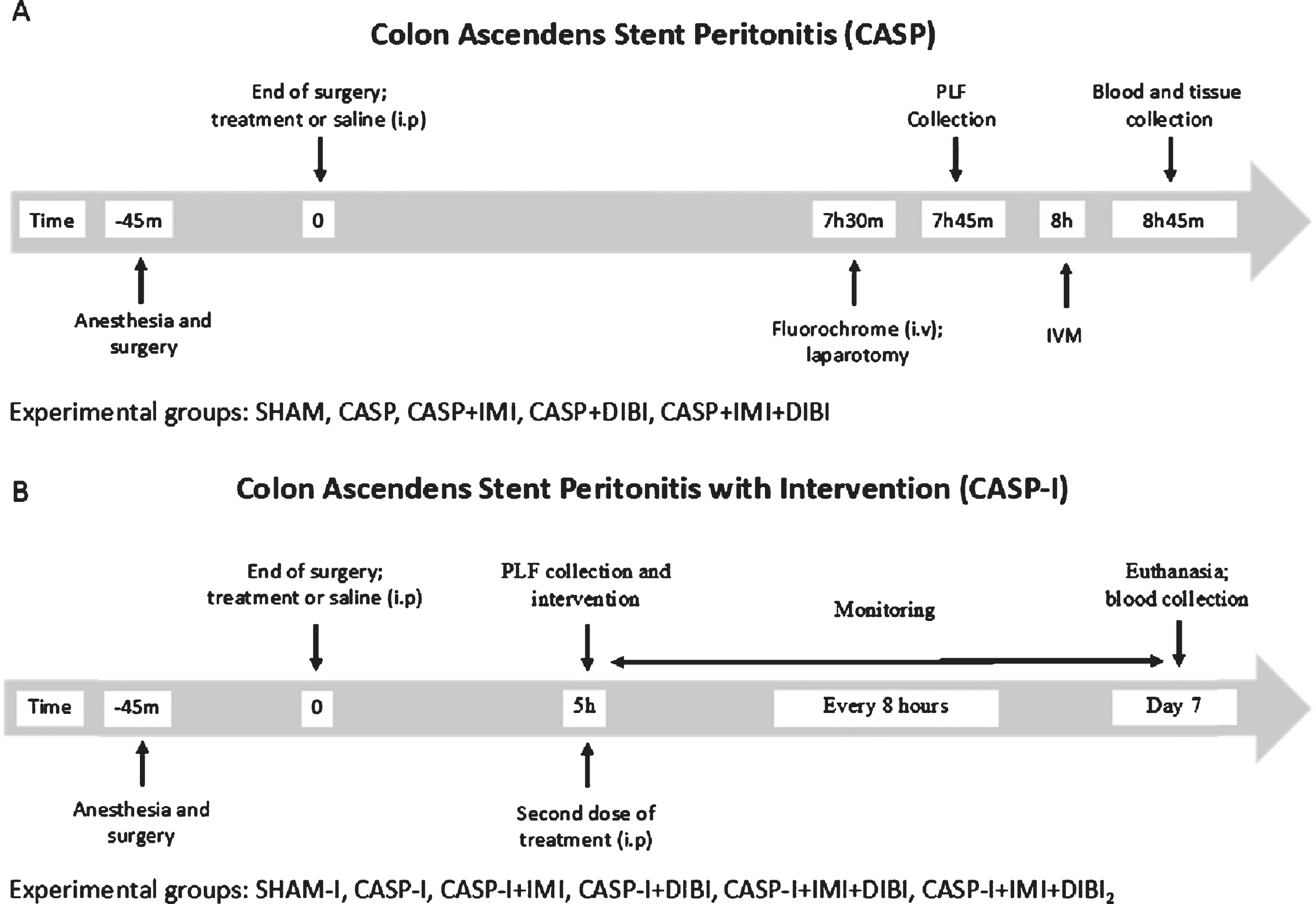
Timelines for CASP and CASP-I Sepsis Models.
In this model, animals received an intervention surgery to remove the CASP stent, the colon was surgically repaired (CASP-I), as described by Traeger et al. [51] with the experimental timelines as shown in Fig. 2 The initial induction surgery and treatments were performed as for CASP, as described above. After recovering for 5 hr from the initial CASP surgery, mice were anesthetized again for a second laparotomy to locate and remove the stent. Following stent removal, the intestinal perforation was sutured, and the abdominal cavity was washed with 2 mL of sterile saline. This PLF was recovered for bacterial enumeration. At the end of the surgery, animals were given 0.5 mL of saline for fluid replacement and allowed to recover. One group was given a second DIBI (DIBI2) administration after peritoneal lavage and close.
Monitoring and data collection
Animals were closely monitored every 2 hr until 8 hr post-surgery for CASP. IVM was then performed. For CASP-I animals, close monitoring was performed until 12 hr post-surgery. Additional ketoprofen was given after 24 and 48 hr. Clinical signs of infection, inflammation and pain were scored using the comprehensive Murine Sepsis Score (MSS) [52], at least every 8 hr. Surface temperature was recorded every 8 hr and weight was recorded every 24 hr. Animals received additional monitoring as necessary. Mice were euthanized if their MSS reached 15, or if other animal welfare concerns existed. Surviving mice were sacrificed at the end of day 7 (T = 168 hr). Blood from the survivors was plated on growth medium for determination of remaining blood bacterial burden and the peritoneal cavity was lavaged to provide PLF for bacterial enumeration.
Treatment groups
Control animals received a sham surgery without active fecal leakage and received a saline vehicle treatment. CASP and CASP-I infected control animals underwent CASP and CASP-I intervention surgeries, respectively and received a saline vehicle treatment. Treatment groups received IMI (25 mg/kg i.p.; Ranbaxy Pharmaceuticals Canada Inc., Mississauga, ON, Canada), DIBI (80 mg/kg) or a combination of IMI and DIBI. All treatments were administered immediately after stent insertion at T = 0 hr. An additional DIBI (80 mg/kg) treatment was administered to the CASP-I + IMI + DIBI2 group after surgical close following the intervention repair surgery laparotomy.
Intravital microscopy
IVM was performed 8 hr after CASP induction. Animals were anesthetized using sodium pentobarbital (90 mg/kg, 54 mg/ml; Ceva Sainte Animale, Montreal, QC, Canada) in a 1:1 dilution with 0.9% sodium chloride (NaCl). Depth of anesthesia was monitored using the pedal withdrawal reflex and additional anesthesia (9 mg/kg, 54 mg/ml; 1:10 dilution) was given as required. Fifteen minutes prior to IVM rhodamine-6G (0.05%, 1.5 ml/kg; Sigma-Aldrich, Oakville, ON, Canada) and fluorescein isothiocyanate-labelled bovine serum albumin (FITC-BSA, 5%, 1 ml/kg; Sigma-Aldrich, Oakville, ON, Canada) were administered i.v.. A laparotomy was performed and the terminal ileum was exposed for IVM. The mouse was placed on its side with a loop of the terminal ileum placed on the viewing platform of a specifically designed stage fixed to a heating pad [53, 54]. A glass slide was placed over the intestine. A continuous flow of 0.9% NaCl heated to 37°C (7 ml/hr) was administered to maintain physiological conditions.
An epifluorescent microscope (Leica DMLM, Wetzlar, Germany) and a light source (LEJ EBQ 100; Carl Zeiss, Jena, Germany) enabled visualization of the intestinal microcirculation. Videos were captured using a digital EM-CCD camera C9100-02 and AC-adapter A3472-07 (Hamamatsu, Herrsching, Germany). Velocity software (PerkinElmer, Waltham MA, USA) was used to acquire videos and store them digitally for later offline analysis. Leukocyte trafficking was assessed using rhodamine-6G staining and a green light filter, within both submucosal collecting venules (V1, diameter 50–100μm) and post capillary venules (V3, diameter 20–40μm). Functional capillary density (FCD) was assessed with FITC-bovine serum albumin (BSA) and a blue light filter with 6 videos being collected of randomly selected field areas of the intestinal muscle layer. Mucosal villi were also exposed for capillary evaluation. For each parameter, 6 videos of 30 seconds in length each were acquired. Animals were sacrificed following IVM with blood and tissue samples collected for further analyses.
Offline image analysis was completed using ImageJ software (version 2.0.0-rc-69/1.52p, National Institute of Health, USA). In V1 and V3 vessels, leukocyte rolling was quantified and the number of adherent leukocytes per area was recorded. FCD was measured by quantifying the total length of perfused vessels in a predefined area.
Bacterial enumeration
In the CASP model, peritoneal lavage fluid (PLF) samples were collected immediately following the laparotomy, prior to IVM. Pre-warmed normal saline (2 mL) was added to the abdominal cavity and this was gently mixed by gentle massage followed by recovery (typical 1.5 mL recovered). Blood from CASP mice was collected by cardiac puncture after IVM. For the CASP-I model, PLF samples (but not blood) were collected after the intervention colon repair surgery (T = 5 hr) and again along with blood, at sacrifice of survivors at 7 days. PLF and blood samples were serially diluted in sterile 1X PBS and plated on either tryptic soy agar (TSA; Millipore Sigma, Etobicoke, Ontario, Canada) or MacConkey (Millipore Sigma, Etobicoke, Ontario, Canada). TSA and MacConkey plates were incubated at 37°C under aerobic conditions. A second set of TSA plates was incubated under anaerobic conditions using a Gas Pak system (BD, Franklin Lakes, NJ, USA). The number of colony-forming units (CFU) per ml of PLF or blood was determined by plate counting of serially diluted plated samples after 16 hr growth.
Plasma cytokines
For the CASP model, whole blood samples taken by cardiac puncture after IVM were centrifuged at 3000 rpm for 10 minutes to separate plasma. Following centrifugation, plasma was stored at –80°C. Plasma levels of selected inflammatory cytokines and adhesion molecules (TNF-α, IL-6, IL-10, IL-1β, and soluble ICAM-1 (s ICAM-1), were analyzed using a custom-made Mouse Magnetic bead-based multiplex assay obtained from Bio-Rad (Mississauga, ON, Canada). Bio-Plex instruments containing Bio-Plex software (Bio-Rad, Mississauga, ON, Canada) according to protocols provided by the manufacturer. Samples were processed as described by the manufacturer and run in duplicate. The plate was read using the Bio-Rad 200 luminometer with Bio-Plex manager software. No cytokine samples were available for the CASP-I model groups as blood could not be usefully or safely taken from these mice.
Histology
Tissue samples from the terminal ileum were collected and fixed in 10% neutral buffered formalin (EK Industries, Joliet, IL) and prepared for histological examination. Tissues were processed and stained using hematoxylin and eosin. Tissues were analyzed in an investigator-blinded fashion and scored for tissue damage using the Chiu scoring system [55]: grade 0, normal histology; grade 1, development of subepithelial Gruenhagen’s spaces; grade 2, extension of Gruenhagen’s spaces with moderate lifting of the epithelium; grade 3, massive epithelial lifting and some denuded tips; grade 4, denuded villi, exposure of lamina propria and dilated capillaries, possible increase in cellularity of lamina propria and, grade 5, disintegration of lamina propria with hemorrhage and ulceration.
Statistical analysis
All data were analyzed using the software GraphPad Prism 8 version 8.2.0 272 (La Jolla, CA, USA). Normal distribution was evaluated by the Kolmogorov-Smirnov test. Statistical significance was assigned to P values that were <0.05. Normally distributed data were analyzed using one-way ANOVA followed by the Newman-Keuls test for multiple comparisons. Non-normally distributed data were analyzed using the Kruskal-Wallis test. Data were expressed as mean±standard error of mean (SEM). Survival data was analyzed using the Mantel-Cox (Log Rank) test.
Results
The overall objective of this study was to investigate the therapeutic responses to DIBI and/or IMI treatment including survival in a CASP-I infection/sepsis model. This CASP-I model includes both the initial CASP-induced peritonitis followed by an early surgical repair of the colon. It was therefore important to first investigate key pathophysiological aspects of a shorter term CASP sepsis course as our previous studies on these aspects were done a full 18 hr after CASP induction. Given overt clinical signs of infection were evident much earlier after CASP initiation, we chose 7.5 hr post CASP induction for these investigations.
Early pathophysiological responses in CASP-induced sepsis
Peritoneal and systemic infection
The surgical control group (SHAM) showed minimal bacterial burden (i.e., CFU near limit of detection (LOD)) in both the PLF and the blood of these mice (Fig. 3). For the untreated sepsis group (CASP) substantial bacterial burdens were evident both in the PLF at 7.5 hr and in the blood at 8.75 hr following sepsis induction. Thus, a substantial bacteremia as indicative of systemic spread of infection from the peritoneal cavity was evident relatively early in this infection. Differential bacterial counts also indicated that the peritonitis and especially the ensuing bacteremia were predominantly associated with Gram-negative facultative anaerobic/aerobic bacteria.
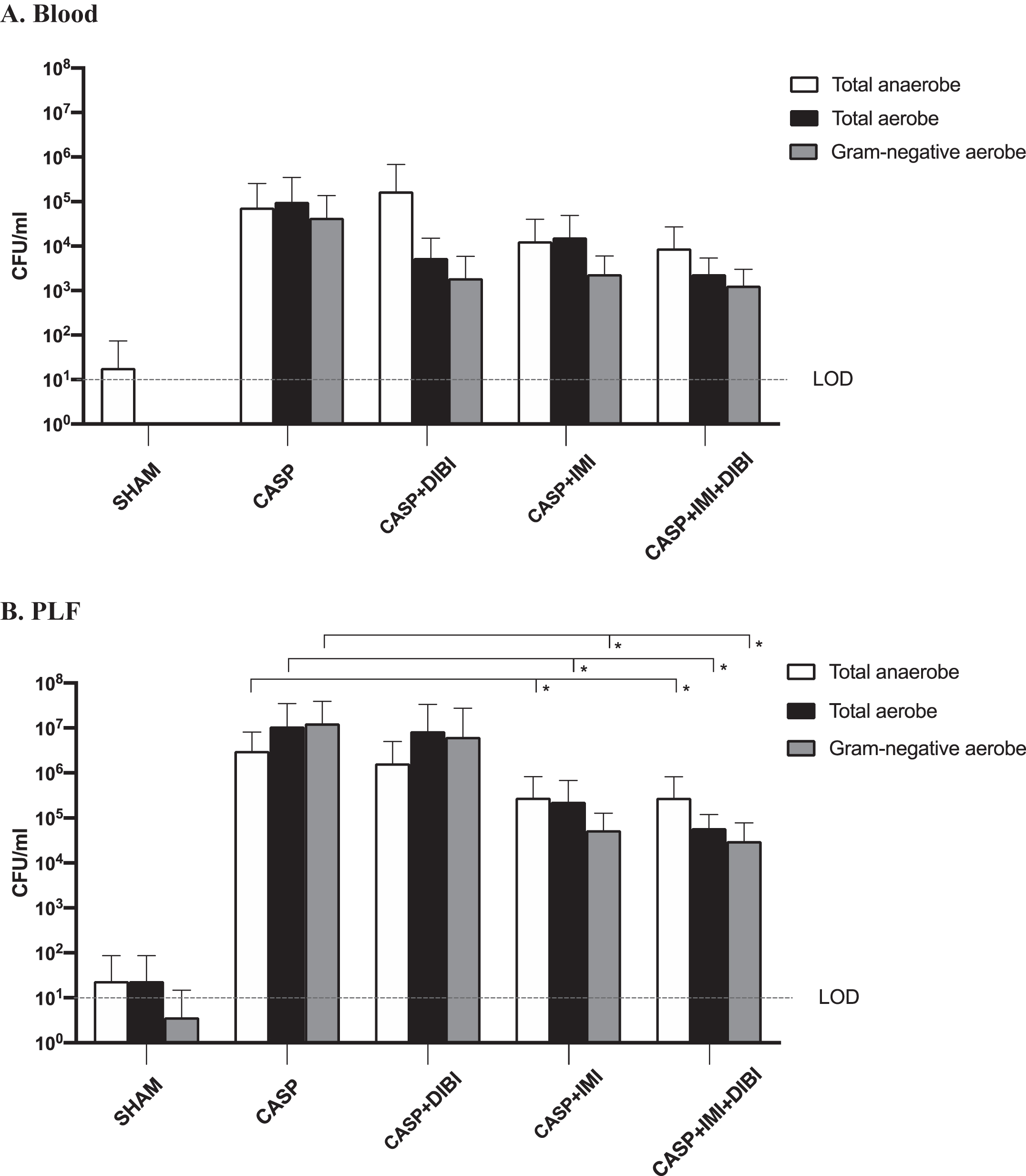
Effects of DIBI and/or imipenem on CASP-induced peritoneal and blood bacterial burdens. Total cultured aerobic and anaerobic bacteria in blood (A) and peritoneal lavage fluid (PLF) (B) as obtained at 8.75 and 7.5 hr after sepsis induction, respectively, for the different treatment groups were determined as CFU after plating on TSA and on MacConkey agar after incubation for 24 h at 37°C. *P < 0.05 compared to SHAM group; # P < 0.05 compared to CASP group. The Lower Limit of Detection (LOD) represent the lowest number of microorganisms that can accurately be estimated.
DIBI did not significantly reduce bacterial burdens in the peritoneal cavity as compared to the untreated CASP group although there was an indication of a reduction in bacteremia (non-statistically significant). However, IMI alone and IMI in combination with DIBI significantly decreased (approximately 100-fold reduction) peritoneal bacterial burdens as compared to the untreated CASP group (Fig. 3B). Both IMI and IMI + DIBI treatments showed non-statistically significant reductions in blood bacterial burdens. However, the blood bacterial burdens for all groups were relatively low (<105 CFU/mL for untreated mice) compared to peritoneal burdens (∼107 CFU/mL) at this early stage of infection.
Adherence of leukocytes in both collecting and post-capillary venules of the intestinal submucosa, assessed by IVM was the primary parameter used to assess systemic inflammation. The untreated sepsis group (CASP) exhibited a significant increase in the number of adhering leukocytes 8.5 hr after sepsis induction, i.e., 7.5X and 5X higher than for SHAM control mice in their collecting and post-capillary venules, respectively (Fig. 4A and B). DIBI, IMI, and the combination of IMI+DIBI, each reduced leukocyte adhesion over untreated infected control values by 38% for DIBI vs CASP, 58% for IMI vs CASP and 74% for DIBI + IMI vs CASP. DIBI only and IMI only treated mice reductions were not significantly different from each other but the combination of both agents (IMI + DIBI group) was the most effective and was significantly better than DIBI alone (p = 0.009) as to reduction of leukocyte adhesion in collecting venules (Fig. 4A). Post-capillary leukocyte adhesion was not significantly different among the treated groups 8.5 hr after sepsis induction (Fig. 4B).

Effects of DIBI and IMI on leukocyte adhesion and FCD. Intestinal intravital microscopy was performed with mice from DIBI and/or imipenem treated groups 8.5 h after CASP sepsis induction. Leukocyte adhesion in intestinal collecting (A) and postcapillary (B) venules. firmly adherent leukocytes per area of submucosa venules (<100μm vessel diameter) quantified as cells/mm2; FCD in the intestinal mucosa (C) and muscle (D) layers. Graphs represent FCD calculated as total length of perfused capillaries within a predetermined field and is quantified as cm/cm2. Each bar graph represents mean values±SEM (n = 5–8 per group). The white bar represents animals from the surgical control group, the black bar represents untreated septic group, and the gray bars represent treated septic groups. ∗P < 0.05 compared to SHAM group; #P < 0.05 compared to CASP group; ×P < 0.05 compared to DIBI group; yP < 0.05 compared to IMI group.
Compared to the surgical control SHAM, untreated sepsis (CASP) mice showed a significant decrease in FCD in their intestinal muscle and mucosa layers 8 hr after sepsis induction (Fig. 4C and D). FCD in the intestinal mucosa layer was significantly improved by all treatments as compared to untreated CASP values: 29% more perfusion in the DIBI group, 37.5% more in IMI group, and 45% more in the DIBI+IMI group. Here again, the combination treatment (IMI + DIBI group) was the only treatment that significantly improved FCD of the intestinal mucosa layer compared to both CASP (p < 0.0001) and DIBI alone (p = 0.0023) (Fig. 4C). In the intestinal muscle layer, neither DIBI nor IMI improved FCD as compared to untreated CASP. The combination treatment (IMI + DIBI) significantly increased FCD compared to the IMI only treatment group (Fig. 4D).
Cytokine response
TNF-α plasma levels were significantly increased by 8.75 hr after sepsis induction in the CASP group compared to the SHAM group (Fig. 5A). IMI and the combination treatment IMI + DIBI significantly reduced TNF-α plasma levels and to near control levels.
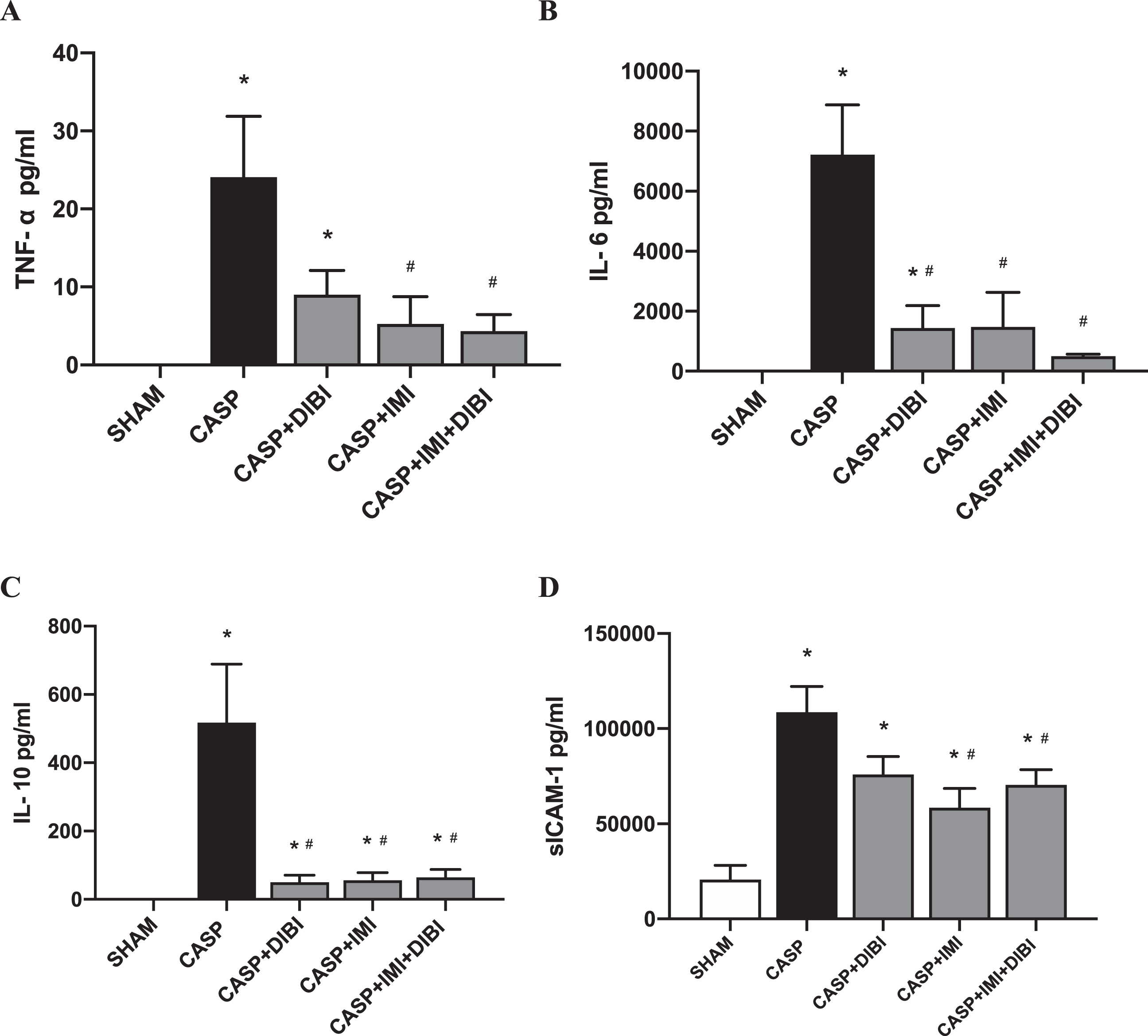
DIBI and/or imipenem effects on plasma inflammatory cytokine levels 8.5 hr after sepsis induction (pg/ml): (A) Tumor Necrosis Factor-Alpha; (B) Interleukin-6; (C) Interleukin-10; (D) sICAM-1. Each bar graph represents mean values±SEM (n = 7–14 per group). The white bar represents animals from the surgical control group, the black bar represents untreated septic groups, and the gray bars represent treated septic groups. ∗P < 0.05 compared to SHAM group; #P < 0.05 compared to CASP group.
For IL-6 and IL-10, CASP induced significant increases of the plasma levels of these cytokines by 8.75 hr after sepsis induction compared to the baseline (SHAM) levels (Fig. 5 B and C). All the treatments significantly decreased plasma IL-6 and IL-10 levels compared to the CASP untreated sepsis group.
CASP sepsis resulted in substantially increased sICAM-1 plasma levels compared to the SHAM group (Fig. 5D). DIBI did not attenuate CASP-induced increases of sICAM-1. However, IMI alone and the IMI + DIBI combination significantly decreased sICAM-1 plasma levels compared to untreated CASP animals.
Assessment of intestinal damage was performed on tissues collected at 8.75 hr after CASP induction with histological samples scored using the Chiu scoring system [55]. The surgical control group (SHAM) exhibited minimal morphological damage. Significantly higher levels of damage to the intestine were observed in untreated CASP mice compared to the SHAM group (Fig. 6) This damage at 8.75 hr was moderate corresponding to a Chiu score ∼2. Treatment with DIBI lowered but did not lessen this damage significantly; Chiu score was not significantly different than the CASP group, However, IMI alone and IMI + DIBI treatments significantly reduced intestinal damage compared to the untreated CASP group (Fig. 6A).
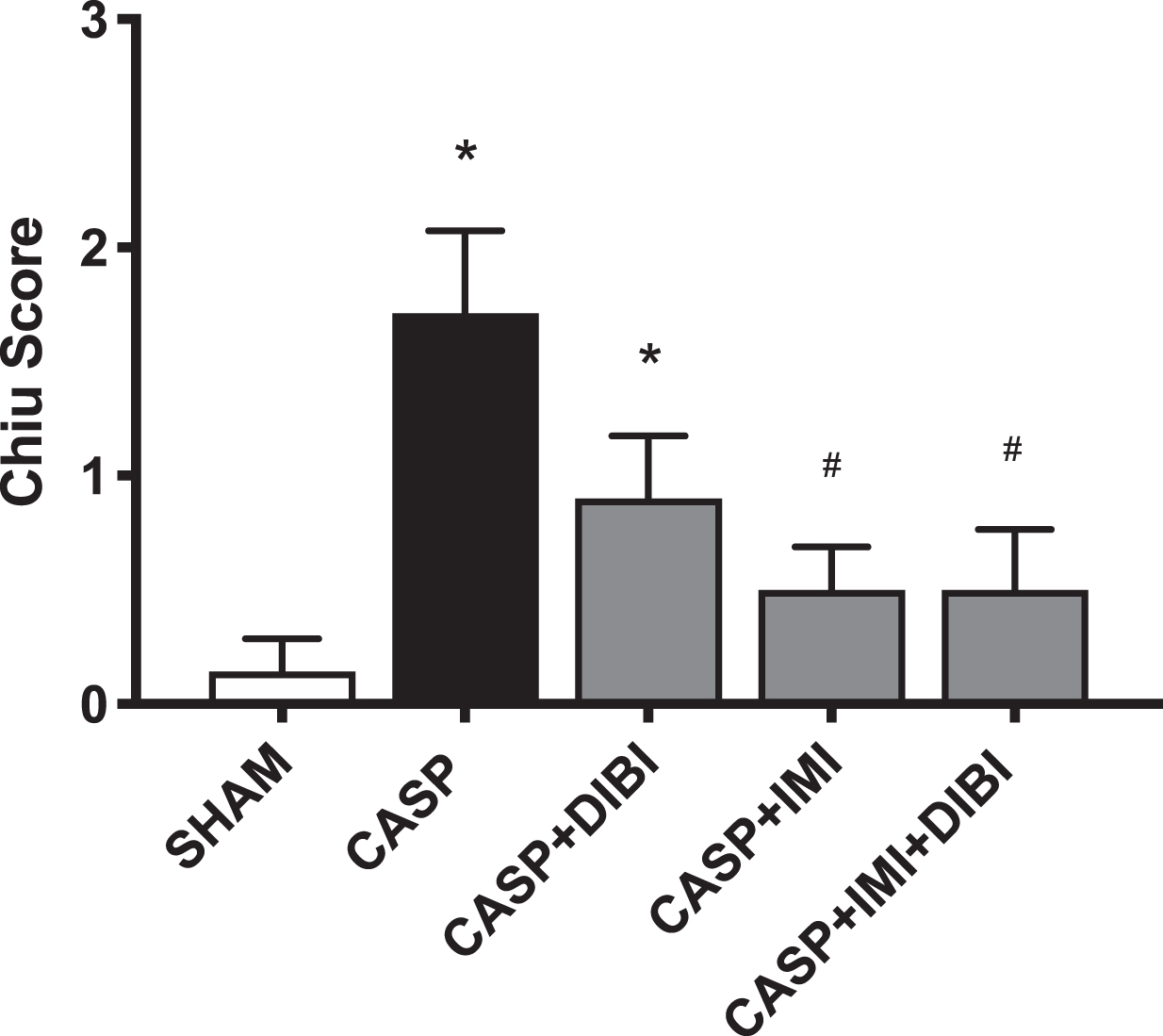
DIBI and imipenem reduction of CASP-induced intestinal tissue damage. Histological sections from mice in the various treatment groups as obtained at sacrifice at 8.75 h after CASP induced infection/sepsis were examined and scored using the Chiu score criteria [55]. Each bar represents mean values±SEM (n = 4–14 per group). ∗P < 0.05 compared to SHAM group; #P < 0.05 compared to CASP group.
The relatively high bacterial burdens that were observed particularly in the blood as early as 8.75 hr following CASP induction (see above Fig. 3) and the evident physical clinical signs of infection early after CASP sepsis induction prompted us to carry out the colon repair CASP-I intervention surgery earlier, i.e., after just 5 hr post CASP sepsis induction. We were concerned about the overall recoverability from a second laparotomy if this were to be left until later in the infection course.
PLF obtained during the colon repair surgery at 5 hr CASP showed correspondingly lower bacterial burdens, i.e., only around 106 CFU/mL which was approx. 10X lower than those observed for untreated infected controls seen at 8.75 hr (Fig. 3). While there appeared to be some reductions in PLF burdens in the IMI and IMI + DIBI treated groups, there were no significant differences among these various treatment groups at this very early stage of infection (data not shown), i.e., except in the case of the SHAM group which had bacterial burdens close to the LOD. As these CASP-I treatment groups were to be followed for survival after the colon repair it was not possible to obtain sufficient blood samples for blood bacterial burden determinations at 5 hr CASP sepsis induction.
The primary outcomes that were assessed for the various CASP-I treatment groups were the clinical course of infection and the long-term survival of mice from infection. The clinical MSS scores for the various treatment groups provided a sensitive grading for clinical symptoms and this showed some important differences among the groups as to the course and the severity of the infection (Fig. 7).
The CASP-I infected/untreated controls showed a rapidly rising MSS reaching a score of 10 by 24 hr with no mice surviving beyond 25 hr (Fig. 7) while the SHAM surgery group had an MSS = 0 and 100% survival for the entire 7-day study. The severity of symptoms in the IMI and the IMI + DIBI treated groups as reflected in MSS were reduced after 24 hr with an MSS approaching or reaching zero for the survivors in those groups. DIBI treatment on its own lowered the MSS and also prolonged survival but no survivors remained beyond 50 hr. A second DIBI treatment administered at 5 hr, i.e., just after at the second laparotomy, further lowered the MSS over that of either IMI or IMI + DIBI (time 0 administration) and also further improved overall survival.
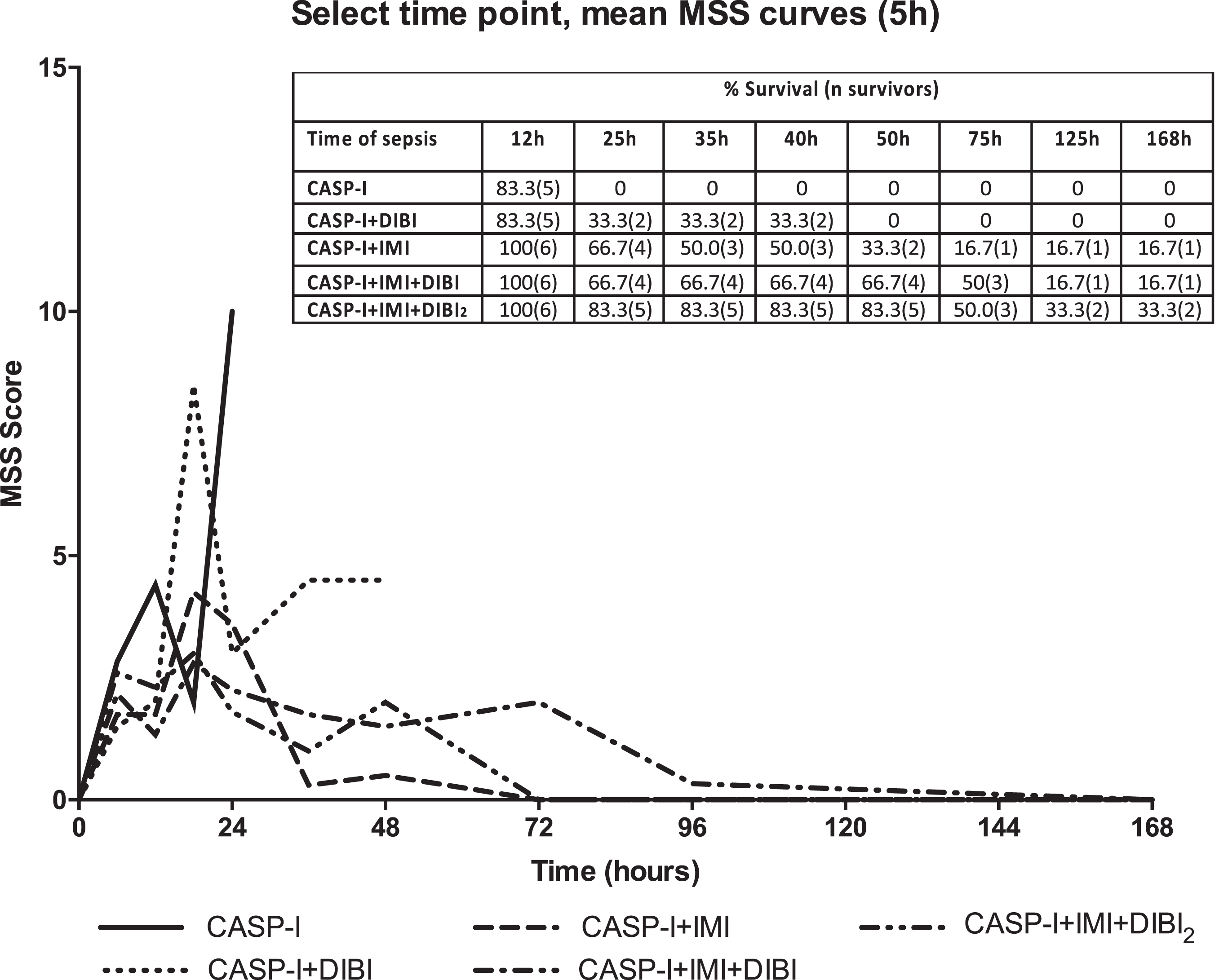
Mean murine sepsis scores for CASP-I treated groups. The data represents the mean murine sepsis scores (MSS) over time for the mice that received an intervention surgery at 5 hr, to remove the stent and close the colon puncture. CASP-I animals received saline at T = 0 hr and T = 5 hr. CASP + DIBI received DIBI (80 mg/kg) at T = 0 hr followed by saline at T = 5 hr. CASP-I + IMI received IMI (25 mg/kg) at T = 0 hr followed by saline at T = 5 hr. CASP-I + IMI + DIBI received IMI and DIBI at T = 0 hr and saline at T = 5 hr. CASP-I + IMI + DIBI2 received IMI and DIBI at T = 0 hr and a second dose of DIBI at T = 5 hr. All groups started with 6 animals. CASP-I: Colon ascendens stent peritonitis with intervention at T = 5 hr; DIBI: iron chelator; IMI: imipenem.
The survival data were analyzed using the Mantel-Cox (Log Rank) test for probability of survival and with Kaplan–Meier plots as shown in Fig. 8. Probabilities of survival for all the treated groups were found to be substantially higher than that for the untreated (CASP-I) group that showed average survival of only 14.4 hr. Average survival for the treatment groups were: DIBI alone = 29.7 hr; IMI alone = 57.7 hr; IMI + DIBI = 82.6 hr and IMI + DIBI(2X) = 94.4 hr.
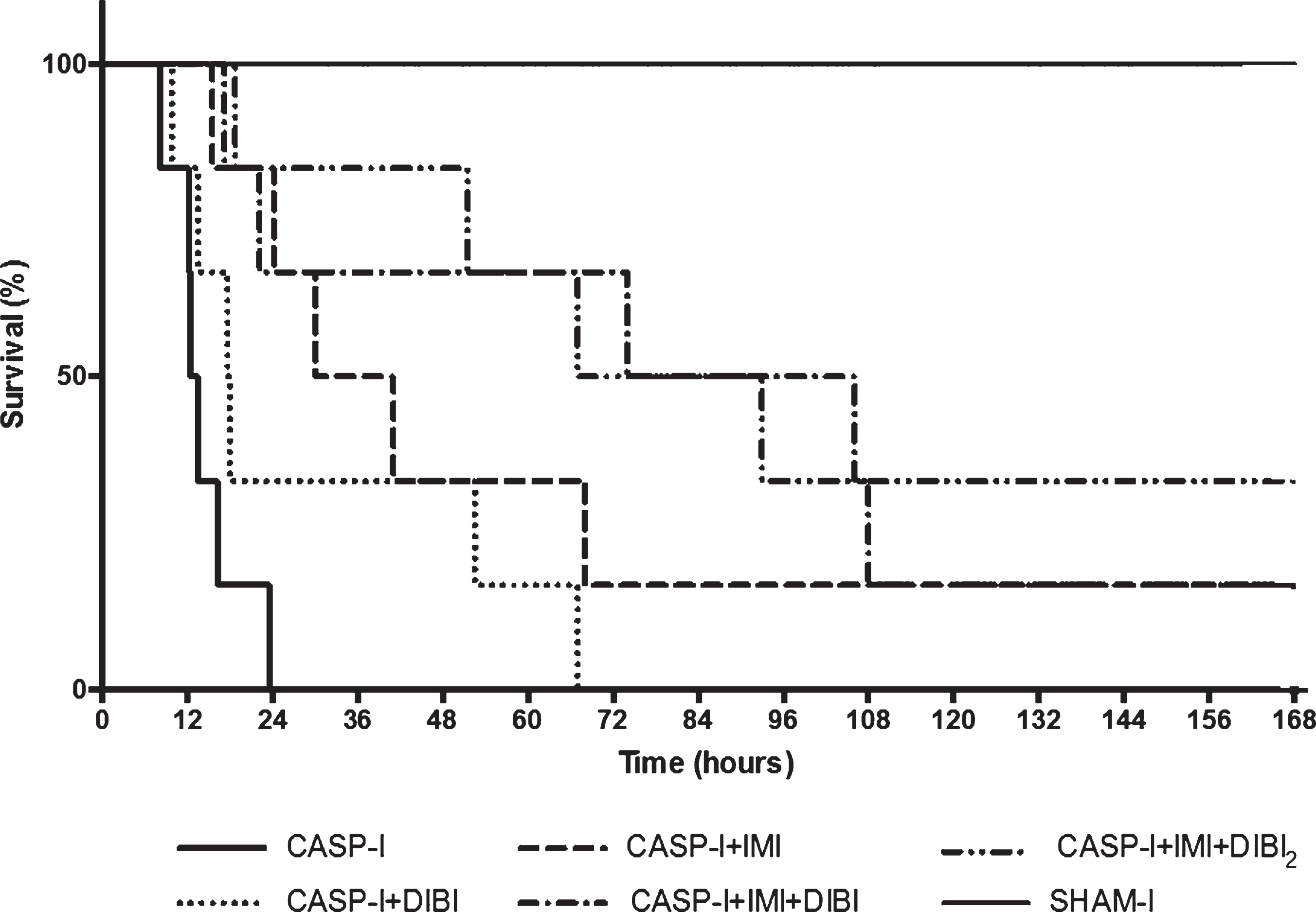
Survival of CASP-I treated groups. The data represents the percent survival of surviving mice that received an intervention surgery at T = 5 hr, to remove the stent and close the colon puncture. Sham-I (n = 5) and CASP-I (n = 6) animals received saline at T = 0 hr and T = 5 hr. CASP + DIBI (n = 6) received DIBI (80 mg/kg) at T = 0 hr followed by saline at T = 5 hr. CASP-I + IMI (n = 6) received IMI (25 mg/kg) at T = 0 hr followed by saline at T = 5 hr. CASP-I + IMI + DIBI (n = 6) received IMI and DIBI at T = 0 hr and saline at T = 5 hr. CASP-I + IMI + DIBI2 (n = 6) received IMI and DIBI at T = 0 hr and a second dose of DIBI at T = 5 hr. CASP-I: Colon ascendens stent peritonitis with intervention at T = 5 hr; DIBI: iron chelator; IMI: imipenem.
At the termination of the CASP-I experiment on day 7, survivors were euthanized and then PLF was obtained for determination of residual bacterial burdens. While relatively small but variable numbers of bacteria (range 0 to 105 CFU/mL) were found in the PLF of the survivors, no detectable bacteria were found in the blood of any of these survivors (Table 1). These results indicated a complete elimination of the systemic infection in all the survivors along with recovery from the associated sepsis, i.e., given survivor MSS values were zero. This apparent full recovery was in spite of the findings of some residual bacteria in the PLF that may not have been fully cleared from the peritoneal cavity, possibly due to residual fecal solids that were initially introduced to the cavity before surgical repair of the colon.
Discussion
The overall aim of this study was to assess the effects of DIBI, an iron sequestering agent with dual anti-infective and anti-inflammatory activities, in a clinically relevant model of infection-induced sepsis. CASP is a standardized and widely used model for polymicrobial abdominal sepsis in rodents including mice [56–61]. Using this model, it is possible to mimic the pathophysiology and clinical course of human abdominal sepsis. The CASP-I model we utilized brings increased clinical relevance because bowel perforation is a common cause of infection-sepsis in humans and typically a laparotomy is performed to repair the perforated bowel and to clean the abdominal cavity by fluid lavage [62].
Bacterial burdens in CASP-I treatment group survivors
Bacterial burdens in CASP-I treatment group survivors
*No CFU grew on plates but LOD = 10CFU.
Our previous studies using the CASP mouse model had shown therapeutic benefits of DIBI on its own and in combination with IMI both for reducing bacterial burdens but also for dampening key inflammatory aspects of the sepsis [48]. However, we had applied the treatments well after sepsis induction (i.e., after 3 hr and 13 hr) and then only examined for their effects a full 18 hr after the induction of the CASP sepsis. For increased clinical relevance we wanted to extend our investigations to include an early surgical repair of the colon breach. This approach therefore benefits from further study of the CASP-induced infection and the inflammatory responses early in the infection course.
Early spread of systemic infection
Bacteremia was evident at 8.75 hr post CASP induction and this was indicative of an early systemic spread of the infection, i.e., beyond the peritoneal cavity. Importantly, differential plating of bacteria found in blood demonstrated that the systemic spread of infection was primarily due to Gram-negative facultative anaerobes/aerobes, thus likely members of the Enterobacteriaceae. Moreover, we found mainly lactose fermenting species such as Escherichia coli. This result might be expected despite the mixed flora of fecal origin of the infection because the peritoneal cavity and the blood are both oxygenated compartments and therefore would not likely support growth of strict fecal anaerobes. The blood compartment would not be expected to support growth of strict anaerobes such as Gram-negative Bacteriodes spp. or Gram-positive Clostridium spp., although these were likely present initially in the peritoneal cavity, i.e., from fecal matter input. Gram negative bacterial septicaemia is often associated with severe sepsis, in part due to the rapid nature of the infection and because Gram-negative bacteria contain (and release) outer membrane lipopolysaccharides (LPS or endotoxin) [46].
Systemic infection response to treatment
DIBI, IMI and IMI + DIBI each provided apparent reduction of the bacterial burden by 8.75 hr but these reductions were not significant this early in the infection course, i.e., likely due to the relatively small blood bacterial burden present (just 105 CFU/mL blood) at this early stage. Our design of following treated animals for survival precluded reliable blood sampling and culturing. Our previous studies [48] where the effects of these treatments on the bacteremia had been assessed later in the infection course (18 hr) had shown significant bacterial reductions with IMI+DIBI. DIBI has also been shown in other studies to have in vivo antimicrobial activity both on its own and also in conjunction with various antibiotics including for the treatment of experimental antibiotic-resistant Acinetobacter baumannii pneumonia/septicemia in mice [47]. A baumannii is also a Gram-negative bacterium that often displays antibiotic resistance and is a major causative pathogen of serious human systemic infection/sepsis.
Early pathophysiological sepsis and treatment responses
Leukocyte activation/adhesion: During the systemic inflammation of sepsis, the circulatory system and thus all organs can be affected. We investigated the early impact of inflammation induced by CASP infection and the effects of both DIBI and IMI using IVM of the microcirculation of the small intestine, i.e., to assess various inter-related pathological aspects as discussed below.
Leukocyte activation and their endothelial wall interactions were evaluated by determining the number of adhering leukocytes in both collecting and post-capillary venules. As expected, a baseline level of leukocyte adhesion was measurable in both collecting and post-capillary intestinal submucosal venules of the surgical control SHAM group, likely due to trauma from the surgical procedures.
CASP-induced sepsis significantly increased leukocyte activation and adhesion 8.5 hr after CASP induction. Previous CASP studies that had assessed leukocyte recruitment and infiltration in various organs [42, 63] corroborate this increased activation of leukocytes in untreated mice. Our various treatments were found to reduce leukocyte adhesion in both collecting and post capillary venules early in the infection course (8.5 hr) as compared to untreated CASP mice but did not completely reverse this to control levels. Our previous investigations had obtained somewhat similar results when examination was made later in the infection (18 hr) [48]. The combination of DIBI and IMI had the strongest dampening effect.
Reduction but not full suppression of leukocyte activation might be considered beneficial during infection/sepsis since the goal for a sepsis treatment is to appropriately dampen the leukocyte hyperactivation but without inducing immunosuppression. Given that infection might be ongoing, an appropriate level of immune cell activation would be clearly needed for adequate host defence.
Capillary perfusion: Microcirculatory dysfunction of the intestines is a major pathophysiologic aspect of sepsis [64]. The mucosa layer is more sensitive to damage due to a more demanding metabolic activity. This layer receives approximately 80% of the total intramural blood flow whereas the muscle layer receives just 20% [65]. Therefore, we measured capillary blood flow in both the intestinal muscle and mucosa layers using IVM. The results for capillary perfusion were consistent with our findings for leukocyte adhesion. For the untreated CASP mice, FCD was decreased in both intestinal muscle and mucosa layers 8.5 hr after the initiation of infection. DIBI and IMI both improved the mucosa FCD. The IMI + DIBI combination further improved mucosa FCD. Consistent with our findings with FCD was the relative degree of tissue damage also observed in histological examination of infected control mice and the corresponding reduction of this damage by the various treatments.
Previous CASP studies had also shown reduction in capillary perfusion [48, 63]. Treatment with DIBI and/or IMI improved capillary perfusion in the mucosa layer, but not in the muscle layer. Al-Banna et al. [61], had not observed any effects of imipenem on the capillary blood flow as assessed by IVM but this had been assessed 16 hr post CASP surgery and only 1 hr post treatment administration. However, Islam et al. [48], 16 hr after CASP surgery and 16 hr post treatment administration found no effect of DIBI and/or IMI treatments on the muscle layer, yet significant improvement of IMI and the combination IMI + DIBI in the mucosa layer. Other models assessing DIBI’s effects on capillary blood flow show yielded conflicting results: sometimes no effects or sometimes improvements [41–44, 66]. The fact that DIBI (+/–imipenem) impacts the two intestinal layers differently might be explained by the higher sensitivity of the mucosa layer to damage but also its response to treatment. CASP-induced abdominal sepsis with and without treatments produced major changes in the mucosa blood flow compared to the muscle layer. Moreover, it is important to note that impairment of capillary blood flow during sepsis is multifactorial and would be affected by various contributing factors such as capillary leakage, coagulopathy, increased leukocyte-endothelial interaction, endothelial cell dysfunction, systemic redistribution of blood flow and tissue edema, externally compressing vessels [64, 67]. It seems reasonable that while in some circumstances a DIBI-related decrease of leukocyte-endothelial interaction could be sufficient to significantly improve capillary perfusion, the various other factors mentioned could also mask any improvement provided by DIBI and/or IMI. Although sepsis pathology is increasingly better understood, the controversy in interpreting capillary blood flow results highlights the likely complexity of damage to the microvasculature caused by sepsis and thus, the need for additional studies.
Cytokine release: Cytokines such as TNF-α, IL-6, IL-10 and ICAM-1 are critical signaling molecules of the immune response while excess cytokine release is a major pathophysiological aspect of sepsis. Upon recognizing a pathogen or being activated by tissue damage, immune cells such as macrophages, neutrophils, lymphocytes and helper T cells start releasing cytokines to initiate diverse inflammatory and healing processes [68–70]. During sepsis there is an exaggerated systemic release of cytokines resulting in the so-called “cytokine storm” that may induce vascular dysfunction, hypo-perfusion, organ dysfunction, and ultimately death [71]. Interestingly, sepsis related dysregulated aspects of this response are not restricted to bacterial infection as the cytokine storm and sepsis responses have also been recently highlighted with the more serious (fatal) cases of SARS-CoV-2 (COVID-19) viral infection [72, Jung,].
Plasma concentrations of key inflammatory cytokines were measured 8.75 hr after CASP induction and we observed early substantial increases of these mediators in the untreated infected mice. DIBI and IMI, and especially their combination treatments, provided reductions in the levels of the pro-inflammatory cytokines, TNF-α and IL-6, the anti-inflammatory cytokine, IL-10 and the adhesion molecule sICAM-1. However, at this early time point during infection these reductions were significant only in the cases of IMI and for IMI + DIBI treatments. In comparing our results to other CASP studies, we noted some discrepancies and some other authors had reported contradictory results [48]. A DIBI only treated group showed significant reduction of TNF-α, IL-1β, IL-6, and IFN-γ plasma levels whereas IMI only and IMI + DIBI impacted only IL-6 plasma levels [48]. A study assessing various cytokines found no significant effects of a single 80 mg/kg DIBI dose on TNF-α, IL-6, IL-12p70, IL-10, IFN-γ, IL-13, IL-17, and sICAM-1 in a CASP-induced sepsis [42]. However, differences as to the effects of DIBI and IMI on cytokines levels might be explained by (i) route of administration (i.v. vs i.p. vs s.c); (ii) differences in the timing and total dose of DIBI and/or imipenem administered and time during the infection-sepsis course when cytokine measurements were taken. Additional studies of DIBI dose response and the timing and frequency of its administration as well as cytokine measurement at different time points during the infection-sepsis course would be useful to better assess DIBI’s full impact on the kinetics of dampening excess cytokine release.
CASP-I infection and effects of treatment
In this study we utilized an important modification to the CASP model that included a surgical repair of the perforated colon after an initial 5 hr period of infection, i.e., continuous fecal leakage to the peritoneal cavity. This CASP-I model therefore should better mimic a likely clinical treatment for a bowel rupture where the source of the infection is surgically controlled, the peritoneal cavity is washed and anti-infectives are administrated.
This model was robust given that all CASP-I infected control mice suffered a rapidly progressing infection and sepsis resulting in 100% mortality by around 25 hr, despite the early repair surgery. Importantly, we demonstrated that the infection rapidly spread systemically from the peritoneal cavity as evidenced by a quickly developing bacteremia. This systemic spread was also primarily caused by facultative aerobic/anaerobic Gram-negative bacteria that would have originated from the fecal input to the peritoneal cavity.
Imipenem is a broad-spectrum antibiotic and one of the main therapeutic choices for polybacterial sepsis. However, with this antibiotic as well as with most other antibiotic choices, there are increasing concerns about the emergence of antibiotic resistant bacteria strains [75].
DIBI is not an antibiotic but rather denies microbial pathogens of an irreplaceably needed micronutrient iron and development of resistance to DIBI has been shown not to occur [76]. When DIBI was administered in combination with imipenem, the infection was further suppressed, and the systemic infection was totally cleared in the surviving mice. In addition, mice receiving both IMI and DIBI appeared clinically improved as compared to those receiving only IMI. IMI + DIBI treated mice were more active, breathing normally and had smoother coats as compared to mice treated with IMI alone.
It is important to keep in mind that our treatments were given just once after the initial surgery and while IMI probably was rapidly distributed systemically based on its known pharmokinetic properties, DIBI being a 9 kDa polymer may have been at least partially retained in the peritoneal cavity at 5 hr post administration. Therefore, a portion of the initial DIBI dose may have been lost by removal with the peritoneal lavage performed at the repair surgery. Interestingly, a second dose of only DIBI, given after the lavage and repair surgery (IMI + DIBI2) significantly improved survival and this may have replenished any DIBI that had been lost through lavage. As well, in an actual clinical setting, antibiotics and/or other treatments including DIBI would likely be given more frequently, for example 2 times a day and for several days. A lower specific dose of DIBI but with repeat administrations over several days both alone and with IMI would be interesting to investigate in this regard.
Our overall findings reveal a potential beneficial impact of DIBI both on its own and as an adjuvant therapy with imipenem and possibly with other antibiotic choices.
The present study does have some limitations, e.g., experiments were performed on healthy young male mice. Although men seem to be more at risk for sepsis and dying from it [77], female patients also suffer from the condition and should be included in future studies.
Conclusion
The dual roles of iron both in bacterial growth and in the immune response and its dysregulation during infection and sepsis represent an important potential therapeutic target for sepsis treatment. The present study investigated the effects of DIBI, an iron sequestering agent with both antimicrobial and anti-inflammatory activities in a clinically relevant sepsis model that features both infection and sepsis inflammation. DIBI demonstrated useful anti-inflammatory properties including reducing the leukocyte-endothelial interactions in collecting and post capillary venules, improving capillary blood flow of the intestinal mucosa layer and it also reduced the excess release of key cytokines. DIBI also reduced and slowed systemic infection on its own improving survival times of infected mice and it greatly improved imipenem efficacy. Survival was substantially improved, and surviving mice treated with IMI and DIBI were found to be free of systemic infection. Our results indicate DIBI has promising potential for sepsis treatment and for use as an adjunct with antibiotics.
Funding & conflicts of interest
BEH has a beneficial interest in Chelation Partners Inc. who funded a portion of this study. The other authors declare no conflicts of interest. This research received funding from the Natural Sciences and Engineering Council (NSERC) of Canada through its CRD program.
Footnotes
Acknowledgments
From Dalhousie University, Halifax, NS, Canada, we would like to thank Dr. David Hoskin (Departments of Pathology and Microbiology and Immunology), Nong Xu (Department of Microbiology and Immunology), Dr. Ashley Stueck and Pat Colp (Department of Pathology), Tanya Myers and Kiyana Kamali (Department of Pharmacology) for their support.