
Abstract
Atherosclerosis is a major cause of cardiovascular disease, in which vascular smooth muscle cells (VSMCs) proliferation and migration play a vital role. Circular RNAs (circRNAs) have been reported to be correlated with the VSMCs function. Therefore, this study is designed to explore the role and mechanism of circRNA lipase maturation factor 1 (circLMF1) in Human aortic VSMCs (HASMCs). The microarray was used for detecting the expression of circLMF1 in proliferative and quiescent HASMCs. Levels of circLMF1, microRNA-125a-3p (miR-125a-3p), vascular endothelial growth factor A (VEGFA), and fibroblast growth factor 1 (FGF1) were determined by real-time quantitative polymerase chain reaction (RT-qPCR). Cell viability, cell cycle progression, and migration were assessed by Cell Counting Kit-8 (CCK-8), flow cytometry, wound healing, and transwell assays, respectively. Western blot assay determined proliferating cell nuclear antigen (PCNA), Cyclin D1, matrix metalloproteinase (MMP2), osteopontin (OPN), VEGFA, and FGF1 protein levels. The possible interactions between miR-125a-3p and circLMF1, and miR-125a-3p and VEGFA or FGF1 were predicted by circbank or targetscan, and then verified by a dual-luciferase reporter, RNA Immunoprecipitation (RIP), RNA pull-down assays. CircLMF1, VEGFA, and FGF1 were increased, and miR-125a-3p was decreased in platelet-derived growth factor-BB (PDGF-BB)-inducted HASMCs. Functionally, circLMF1 knockdown hindered cell viability, cell cycle progression, and migration in PDGF-BB-treated HASMCs. Mechanically, circLMF1 could regulate VEGFA or FGF1 expression through sponging miR-125a-3p. Our findings revealed that circLMF1 deficiency could inhibit cell viability, cell cycle progression, and migration of PDGF-BB stimulated atherosclerosis model partly through the miR-125a-3p/VEGFA or FGF1 axis, suggesting that targeting circLMF1 can be a feasible therapeutic strategy for atherosclerosis.
Introduction
As a major cause of cardiovascular disease, atherosclerosis is a vascular disorder characterized by the accumulation of fibrous components and lipids in the vascular walls [1]. Currently, the increasing incidence of atherosclerosis has brought great medical and economic burdens to society in China [2, 3]. Intriguingly, abnormal proliferation and migration of vascular smooth muscle cells (VSMCs), the main component of blood vessel walls, have been reported to promote the formation of atherosclerotic plaque [4–6]. Recent studies suggested that platelet-derived growth factor-BB (PDGF-BB) played a vital role in regulating VSMCs proliferation and migration through the activation of intracellular signal transduction pathways [7, 8]. Thus, repressing PDGF-BB-inducted VSMCs proliferation and migration might be an effective way for attenuating atherosclerosis progression.
Up to now, an extensive body of recent research has focused on non-coding RNAs, particularly circular RNAs (circRNAs), which are formed by back-splicing covalently closed-loop structures [9, 10]. Therefore, unlike linear RNA, circRNAs are resistant to the degradation of RNA exonuclease and more stable [11, 12]. It is becoming increasingly apparent that numerous circRNAs, as essential regulators, take part in the modulation of the progression of various diseases, including atherosclerosis [13, 14]. For example, Peng et al. displayed that the downregulation of circRNA circDHCR24 prevented vascular restenosis by alleviating the proliferation and migration of aortic smooth muscle cells via sponging miR-149-5p [15]. Similarly, Ding et al. found that the forced expression of circ_0010283 promoted cell viability and migration of vascular smooth muscle cells by regulating the miR-370-3p/HMGB1 axis in atherosclerosis [16]. Consistently, Yu et al. presented that circRNA_0029589 exacerbated vascular smooth muscle cell injury by regulating IGF12 via miR-424-5p [17]. These findings suggested the important role of circRNAs in atherosclerosis progression.
In this paper, we found that circRNA lipase maturation factor 1 (circLMF1) originated from exons 6, 7, and 8 of the LMF1 gene was increased in proliferating conditions of human aortic SMCs (HASMCs) and PDGF-BB-treated HASMCs. Here, we aimed to explore whether circLMF1 regulated the proliferation and migration of PDGF-BB-stimulated HASMCs in a microRNA (miRNA)-mRNA-dependent manner.
Materials and methods
Microarray analysis
CircRNA Microarray (GSE77280), which has been deposited at Gene Expression Omnibus (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE77278), was utilized for the global profiling of human circRNAs. Microarray analysis was conducted using GEO2R (https://www.ncbi.nlm.nih.gov/geo/geo2r/), which is an online tool for processing data using GEO queries and limma packages [18] in R from the Bioconductor project [19]. And volcano plot filtering was applied to identify the differential expression of circLMF1 with statistical significance.
Cell culture
In a humidified incubator with 5%CO2 at 37°C, Human aortic smooth muscle cells (HASMCs, ScienCell, Carlsbad, CA, USA) was grown in Dulbecco’s modified Eagle’s medium (DMEM; Hyclone, Logan, UT, USA) containing 10%fetal bovine serum (FBS; Hyclone) and 1%penicillin/streptomycin (Hyclone). In this study, quiescent HASMCs were t FBS starved for 24 h, and proliferative HASMCs were acquired from quiescent HASMCs treated with PDGF-BB (10 ng/mL, R&D Systems, Minneapolis, Minnesota, USA).
Real-time quantitative polymerase chain reaction (RT-qPCR)
After treatment with TRIzol reagent (Invitrogen, Carlsbad, CA, USA), extracted total RNAs from cells were subjected to reverse transcription to synthesize cDNA with RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific, Barrington, IL, USA), according to the operation manual. Whereafter, in line with the instruction guidelines of SYBR Green PCR Kit (TaKaRa), RT-qPCR reaction was conducted using Thermal Cycler CFX6 System (Bio-Rad, Hercules, California, USA). And then, the results were normalized to GAPDH (for CircLMF1, VEGFA, and FGF1) and U6 (for miR-125a-3p), and the relative changes in gene expression were calculated. The specific primers were presented in Table 1.
The sequences of primers for RT-qPCR used in this study
The sequences of primers for RT-qPCR used in this study
Actinomycin D and RNase R was used to examine the stability of circLMF1. After the culture medium was treated with Actinomycin D (10 mM, Sigma-Aldrich, St. Louis, MO, USA) for different time points, HASMCs (1×105 cells/well) in 6-well plates were harvested, followed by extraction with TRIzol reagent (Invitrogen). And the assessment of the expression of circLMF1 and GAPDH was performed using RT-qPCR assay. For RNase R digestion, total RNA was incubated with 3 U/mg of RNase R (Epicentre, San Diego, California, USA) at 37°C. After treatment for 15 min, the expression of circLMF1 and GAPDH was detected.
Subcellular fractionation assay
According to the instructions of the Nuclear and Cytoplasmic Extraction Kit (Thermo Scientific), cell nucleus and cytoplasm fraction were separated, followed by purification with TRIzol reagent (Invitrogen). Whereafter, the sample was subjected to RT-qPCR analysis of circLMF1, 18S rRNA (cytoplasm control), and U6 (nucleus control).
Cell transfection
CircLMF1 small interference RNA (si-circLMF1-1 and si-circLMF1-2), miR-125a-3p mimic (miR-125a-3p), miR-125a-3p inhibitor (anti-miR-125a-3p), and their respective controls (si-NC, miR-NC, anti-miR-NC) were designed and synthesized from Ribobio (Guangzhou, China). Moreover, VEGFA or FGF1 overexpression vector was obtained by introducing the sequence of VEGFA or FGF1 into pcDNA vector (Invitrogen). As per the user’s guidebook of Lipofectamine 3000 reagent (Invitrogen), the above oligonucleotides (50 nM) and vectors (200 ng) were transfected into HASMCs at 37°C for 48 h. Then, transfected cells were collected for the subsequent assays.
Cell counting kit (CCK-8) assay
In this assay, HASMCs were seeded into a 96-well plate for 24 h at a density of 2000 cells/well, followed by addition with CCK-8 (10μL, Dojindo, Kumamoto, Japan). After incubation for another 4 h, the absorbance at 450 nm was determined using a microplate reader (Bio-Rad) based on the supplier’s direction.
Cell cycle assay
After washing with PBS (Invitrogen) twice, HASMCs were fixed with 70%ethanol at 4°C overnight, followed by staining with Propidium Iodide (PI) (Beyotime, Nantong, China) for 30 min at room temperature. According to the operation manual of FACSan flow cytometry (Becton Dickinson, San Jose, CA, USA), the percentages of cells in the G0-G1, S, and G2-M phases were analyzed.
Wound healing assay
In this assay, after trypsin and reseeding, HASMCs in 6-well plates were cultured to 90%confluency. Subsequently, a scratch was made with a 200μL sterile pipette tip (record 0 h), creating a clear cell-free zone. After the replacement of the medium with a serum-free medium, the cells were allowed to migrate for 24 h. At last, images of migration were taken using an inverted microscope (Olympus, Tokyo, Japan) (magnification×100), and the gap size was analyzed using Image J software (National Institutes of Health, Bethesda, MD, USA).
Transwell assay
Furthermore, the measurement of HASMCs migration was performed using Transwell chambers (Corning Incorporated, Corning, NY, USA), according to the instructions guidelines. In short, 2×105 HASMCs were cultured in medium with 1%FBS in the upper chamber, while 20%FBS was added to the bottom counterparts as a chemoattractant. After incubation for 24 h, migrated cells on the surface of the bottom counterparts were stained with 0.1%crystal violet (Sigma-Aldrich, St. Louis, MO, USA) for 15 min. Finally, an inverted microscope (Olympus, magnification×100) was carried out for the observation of migrated cells in this assay.
Western blot assay
Total protein from HASMCs was isolated according to the instruction guidelines of pre-cold RIPA buffer (Beyotime, Nantong, China), followed by quantification with BCA Protein Assay Kit (Beyotime). Whereafter, protein samples (30μg) were subjected to 10%SDS-PAGE electrophoresis, which were then transferred to nitrocellulose membranes (Millipore, New York, NY, USA). After blocking with 5%skim milk for 2 h, the immunoblots were incubated with primary antibodies against proliferating cell nuclear antigen (PCNA, 1:1000, ab29, Abcam, Cambridge, MA, USA), Cyclin D1 (1:1000, ab134175, Abcam), matrix metalloproteinase (MMP2, 1:1000, ab37150, Abcam), osteopontin (OPN, 1:1000, ab8448, Abcam), VEGFA (1:1000, ab46154, Abcam), FGF1 (1:1000, ab9588, Abcam), and β-actin (1:5000, ab52614, Abcam) at 4°C overnight, followed by incubation with secondary antibody (ab205178, 1:10000, Abcam). At last, the ECL detection system (GE Healthcare, Piscataway, NJ, USA) was used for visualization, and the Image J software (National Institutes of Health) was employed for analysis.
Dual-luciferase reporter assay
Generally, circLMF1 wild-type (WT) reporter vector (WT-circLMF1) possessing the binding sequences for miR-125a-3p and its mutant-type in complementary (MUT-circLMF1), and VEGFA or FGF1 3’ untranslated region (3’UTR) WT reporter vector (WT-VEGFA or FGF1 3’UTR) containing the binding sites with miR-125a-3p and its mutant-type in the seed region (MUT-VEGFA or FGF1 3’UTR) were synthesized and inserted into psiCHECK vector (Promega, Madison, WI, USA). Whereafter, these reporters (50 ng) were co-transfected into HASMCs with miR-125a-3p or miR-NC (20 nM), followed by analysis using a dual-luciferase reporter assay kit (Promega).
RNA immunoprecipitation (RIP)
According to the user’s guidebook of a commercial Magna RNA immunoprecipitation kit (Millipore, Bedford, MA, USA), RIP assay was employed in HASMCs. In brief, transfected cells in the ice-cold RIP lysis buffer were lysed. Subsequently, the cell extracts were incubated with anti-Argonaute 2 antibody (Anti-Ago2; Millipore) or negative control normal mouse IgG (Millipore) at 4°C for 4 h before treating magnetic beads for 2 h, followed by washing with PBS twice. After digestion with proteinase K (Invitrogen), immunoprecipitated RNAs were isolated and further decoction of RT-qPCR assay.
RNA pull-down assay
In this assay, HASMCs were transfected with 50 nM biotinylated bio-miR-NC, WT-bio-miR-125a-3p, and MUT-bio-miR-125a-3p for 48 h. After washing with PBS, the samples were incubated in lysis buffer, followed by treatment with M-280 streptavidin magnetic beads (Sigma-Aldrich) for 3 h. Finally, these bound RNAs were eluted by TRIzol reagent (Invitrogen) and analyzed by RT-qPCR reaction.
Statistical analysis
Statistical analysis was performed using GraphPad Prism7 software. Results were reported as mean±standard deviation (SD) from at least three biological replicates. The statistical significance of differences was assessed using Student’s t-test or one-way analysis of variance (ANOVA) with Tukey’s tests. If P-value < 0.05, it was deemed as statistically significant.
Results
CircLMF1 expression was increased in PDGF-BB-stimulated HASMCs
Firstly, circLMF1 in proliferative and quiescent HASMCs in the database GSE77280 was analyzed. As shown in Fig. 1A and 1B, circLMF1 was expressed at a high level in proliferating HASMCs compared with that of in quiescent HASMCs. Then, we found that circLMF1 was derived from exon 6, exon 7, and exon 8 of the LMF1 gene (Fig. 1C). Subsequently, we designed divergent primers to amplify circLMF1 and convergent primers to amplify GAPDH mRNA. As shown in Fig. 1D, circLMF1 was only detectable in cDNA but not genomic DNA (gDNA) from HASMCs by RT-qPCR with divergent primers, whereas GAPDH could be amplified in both cDNA and gDNA using convergent primers. Besides, a previous study suggested that PDGF-BB accelerated the dedifferentiation and proliferation of VSMCs, and was increased at the site of vascular injury and atherosclerotic plaque specimens [20]. Therefore, we detected the response of circLMF1 to PDGF-BB in cultured HASMCs. RT-qPCR analysis showed that circLMF1 was upregulated in HASMCs treated with PDGF-BB relative to the control groups (Fig. 1E). To further validate the circular characteristics of circLMF1, we treated HASMCs treated with Actinomycin D (a transcriptional inhibitor) and RNase R. Data suggested that the half-life of circLMF1 transcript exceeded 24 h, while that of GAPDH was less than 12 h (Fig. 1F), implying that circLMF1 is more stable than the linear mRNA transcript in HASMCs. Meanwhile, RNase R digestion results presented that circLMF1, but not housekeeping mRNA GAPDH, was highly resistant to RNase R (Fig. 1G). Furthermore, subcellular fractionation results suggested that circLMF1 was predominantly located in the cytoplasm of HASMCs (Fig. 1H), implying the underlying post-transcriptional regulatory mechanism of circLMF1 in HASMCs. In addition, considering the upregulation of circLMF1 in PDGF-BB-induced HASMCs, we knocked down circLMF1 expression. Results presented that circLMF1 level was apparently downregulated in PDGF-BB-induced HASMCs transfected with si-circLMF1-1 or si-circLMF1-2 (Fig. 1I), suggesting the transfection efficiency was successful for the subsequent loss-of-function analyses. Especially in si-circLMF1-1-transfected HASMCs, thus we selected the transfected cells for subsequent assays. Together, these data indicated that circLMF1, a highly stable circRNA, might be the regulator in PDGF-BB-stimulated HASMCs.

CircLMF1 expression was upregulated in PDGF-BB-induced HASMCs. (A) Volcano plot exhibited that the expression level of circLMF1 in proliferative HASMCs and quiescent HASMCs in the database GSE77280. (B) The expression value of circLMF1 in the database GSE77280 was presented. (C) Three exons from circLMF1 by back splicing from the chromosomal region. (D) Divergent primers detected circular RNAs in cDNA but not gDNA. (E) RT-qPCR assay was applied to assess the expression level of circLMF1 in HASMCs treated with or without PDGF-BB (10 ng/mL) for 24 h. (Student’s t-test). (F) Relative RNA levels of circLMF1 and GAPDH were detected in HASMCs treated with Actinomycin D. (ANOVA with Tukey’s tests). (G) CircLMF1 and GAPDH levels were examined in HASMCs after treatment with RNase R. (Student’s t-test). (H) Subcellular fractionation assay was performed to analyze the cellular localization of circLMF1 in HASMCs. (Student’s t-test). (I) CircLMF1 level was measured in HASMCs transfected with si-NC, si-circLMF1-1, and si-circLMF1-2. (ANOVA with Tukey’s tests). *P < 0.05.
The effect of circLMF1 on the phenotypic changes of PDGF-BB-induced HASMCs was examined by knocking-down the circLMF1. As shown in Fig. 2A, the treatment of PDGF-BB promoted cell viability of HASMCs, while the downregulation of circLMF1 significantly attenuated the effect. Meanwhile, flow cytometry analysis indicated that PDGF-BB resulted in a significant decrease in the proportion of G0/G1-phase and a significant increase in the proportion of S-phase, which was effectively overturned by circLMF1 downregulation in HASMCs (Fig. 2B). That is to say, the transfection of si-circLMF1 could abolish the promotive action of PDGF-BB on cell cycle progression. Furthermore, wound healing assay displayed that enhanced migratory rate caused by PDGF-BB treatment was obviously relieved through circLMF1 silencing in HASMCs (Fig. 2C). Consistent with wound healing results, transwell assay analysis indicated that the knockdown of circLMF1 mitigated the positive effect of PDGF-BB on migration in HASMCs (Fig. 2D). Then, we detected the PCNA and Cyclin D1 (proliferation-related factor), and MMP2 and OPN (migration-associated factor) in HASMCs. As shown in Fig. 2E, PDGF-BB induced an increase of PCNA, Cyclin D1, MMP2, and OPN was reverted after the introduction of si-circLMF1 in HASMCs. All of these data suggested that circLMF1 knockdown could attenuate PDGF-BB-induced proliferation and migration of HASMCs.
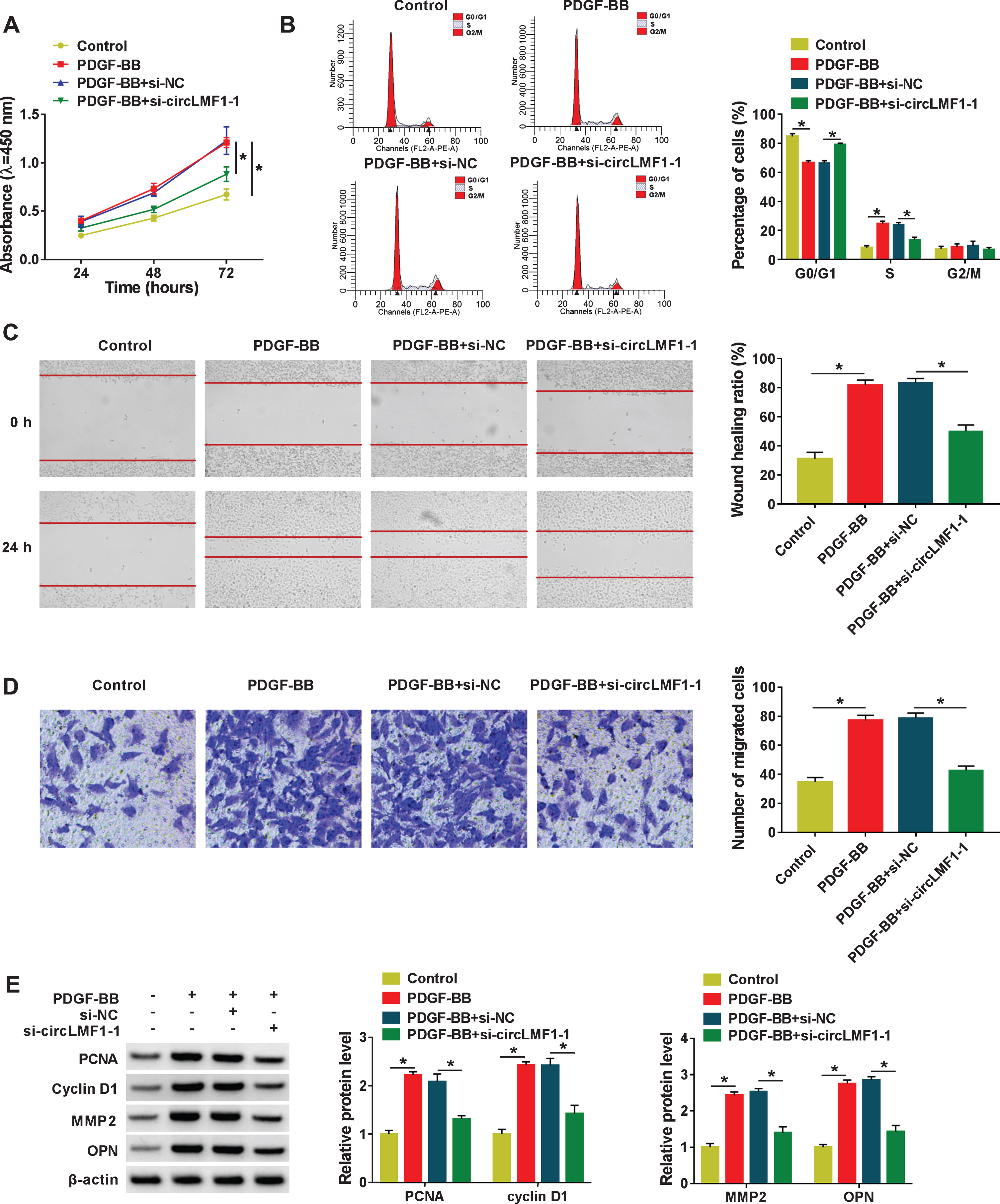
CircLMF1 knockdown suppressed cell viability, cell cycle progression, and migration in PDGF-BB-induced HASMCs. HASMCs were treated with Control, PDGF-BB, PDGF-BB + si-NC, PDGF-BB + si-circLMF1-1. (A) CCK-8 assay was used to assess cell viability in treated HASMCs. (ANOVA with Tukey’s tests). (B) Flow cytometry assay was applied to examine cell cycle distribution in treated HASMCs. (ANOVA with Tukey’s tests). (C) Wound healing assay was conducted to detect migration rate in treated HASMCs. (ANOVA with Tukey’s tests). (D) Migration in treated HASMCs was assessed using transwell assay. (ANOVA with Tukey’s tests). (E) Protein levels of PCNA, Cyclin D1, MMP2, and OPN in treated HASMCs were determined. (ANOVA with Tukey’s tests). *P < 0.05.
In view of the cytoplasmic localization of circLMF1 in HASMCs, we explored whether circLMF1 could exert the function in a miRNA-mRNA-dependent manner. Through circBank software analysis, we found that circLMF1 and miR-125a-3p share some complementary sequences (Fig. 3A). Synchronously, the upregulation or downregulation of efficiency of miR-125a-3p was detected and presented in Fig. 3B and 3C. Subsequently, the dual-luciferase reporter assay was used to examine the possible interaction between circLMF1 and miR-125a-3p in HASMCs. Results exhibited that the luciferase activity in HASMCs transfected with WT-circLMF1 and miR-125a-3p mimic was significantly declined relative to that in cells transfected with WT-circLMF1 and miR-NC, while there was little effect in the cells with MUT-circLMF1 (Fig. 3D). Apart from that, to further verify the mutual effect of circLMF1 and miR-125a-3p at endogenous levels, RIP assay was employed using antibody Ago2, a key component of RISC complex. As shown in Fig. 3E, circLMF1 and miR-125a-3p in HASMCs were significantly enriched in the Ago2 groups in comparison with the IgG groups. Also, compared with the bio-miR-NC and MUT-bio-miR-125a-3p groups, RNA pull-down analysis indicated that circLMF1 enrichment in the WT-bio-miR-125a-3p group was significantly intensified in HASMCs, suggesting the interaction between circLMF1 and miR-125a-3p (Fig. 3F). And the downregulation of miR-125a-3p in PDGF-BB-treated HASMCs was viewed versus the control group (Fig. 3G), implying the involvement of miR-125a-3p in PDGF-BB-induced HASMCs damage. In a word, miR-125a-3p was a direct target of circLMF1 in HASMCs.

MiR-125a-3p was a functional target of circLMF1. (A) CircBank software was utilized to predict the binding site between circLMF1 and miR-125a-3p. (B) Relative miR-125a-3p expression was detected in HASMCs transfected with miR-NC or miR-125a-3p. (Student’s t-test). (C) MiR-125a-3p level was measured in HASMCs transfected with anti-miR-NC or anti-miR-125a-3p. (Student’s t-test). (D) A dual-luciferase reporter assay was used to confirm the binding. (Student’s t-test). (E) RIP assay was performed in HASMCs cell extracts to detect miR-125a-3p endogenously associated with circLMF1. (Student’s t-test). (F) RNA pull-down assay was employed to further verify their interaction. (ANOVA with Tukey’s tests). (G) MiR-125a-3p level was assessed in HASMCs treated with control or PDGF-BB. (Student’s t-test). *P < 0.05.
As mentioned above, circLMF1 played an important role in the proliferation and migration of PDGF-BB-stimulated HASMCs, and miR-125a-3p was a putative target of circLMF1. Therefore, we further tunneled the impact of circLMF1 in the proliferation and migration of HASMCs was related to miR-125a-3p. In PDGF-BB-stimulated HASMCs, the repression of cell viability and cell cycle progression on account of si-circLMF1 was significantly weakened via miR-125a-3p downregulation (Fig. 4A and 4B). Simultaneously, the re-introduction of anti-miR-125a-3p significantly counteracted the suppressive effect of circLMF1 deletion on migration in PDGF-BB-stimulated HASMCs (Fig. 4C and 4D). What’s more, the deficiency of circLMF1 led to a significant reduction in the protein levels of PCNA, Cyclin D1, MMP2, and OPN was significantly recuperated after the transfection of miR-125a-3p inhibitor (Fig. 4E). All of these results suggested that circLMF1 knockdown suppressed PDGF-BB-stimulated proliferation and migration by regulating miR-125a-3p.

Downregulation of miR-125a-3p abrogated the inhibitory action of circLMF1 silencing on proliferation and migration in PDGF-BB-induced HASMCs. HASMCs were treated with PDGF-BB + si-NC, PDGF-BB + si-circLMF1-1, PDGF-BB + si-circLMF1-1 + anti-miR-NC, and PDGF-BB + si-circLMF1-1 + miR-125a-3p. (A and B) Cell viability and cycle progression were determined by CCK-8 and flow cytometry assay in treated HASMCs. (ANOVA with Tukey’s tests). (C and D) Migration was examined by wound healing and Transwell assays in treated HASMCs. (ANOVA with Tukey’s tests). (E) Protein levels of PCNA, Cyclin D1, MMP2, and OPN in treated HASMCs were measured. (ANOVA with Tukey’s tests). *P < 0.05.
It has been confirmed that miRNAs play the biological effect mainly by their target genes. Thus, targetscan software was conducted to search for the possible targets of miR-125a-3p. As shown in Fig. 5A and 5B, miR-125a-3p and VEGFA 3’UTR or FGF1 3’UTR share some complementary sites, respectively. Whereafter, dual-luciferase reporter assays suggested that the forced expression of miR-125a-3p reduced the luciferase activity of WT-VEGFA 3’UTR or WT-FGF1 3’UTR reporter but not that of MUT-VEGFA 3’UTR or MUT-FGF1 3’UTR reporter in HASMCs (Fig. 5C and 5D). Also, RIP assays indicated that miR-125a-3p, VEGFA, and FGF1 in the Ago2 groups were higher than in the IgG groups (Fig. 5E). Besides, consistent with bioinformatics analysis, and the results of the dual-luciferase reporter and RIP assays, the significant augment of VEGFA and FGF1 enrichment was noticed in the WT-bio-miR-125a-3p group in RNA pull-down assays (Fig. 5F). Interestingly, we found that the manipulation of miR-125a-3p expression changed the expression of VEGFA or FGF1, exhibiting that VEGFA or FGF1 expression was reduced by miR-125a-3p upregulation, and VEGFA or FGF1 was enhanced due to miR-125a-3p knockdown in HASMCs (Fig. 5G and 5H). Also, the trend of VEGFA or FGF1 protein expression in HASMCs was in line with the mRNA expression under the same conditions (Fig. 5I and 5J). Additionally, RT-qPCR and western results suggested that the mRNA level and protein level of VEGFA or FGF1 were upregulated in PDGF-BB-induced HASMCs relative to their several controls (Fig. 5K and 5N). Overall, these data discovered that miR-125a-3p interacted with VEGFA or FGF1.
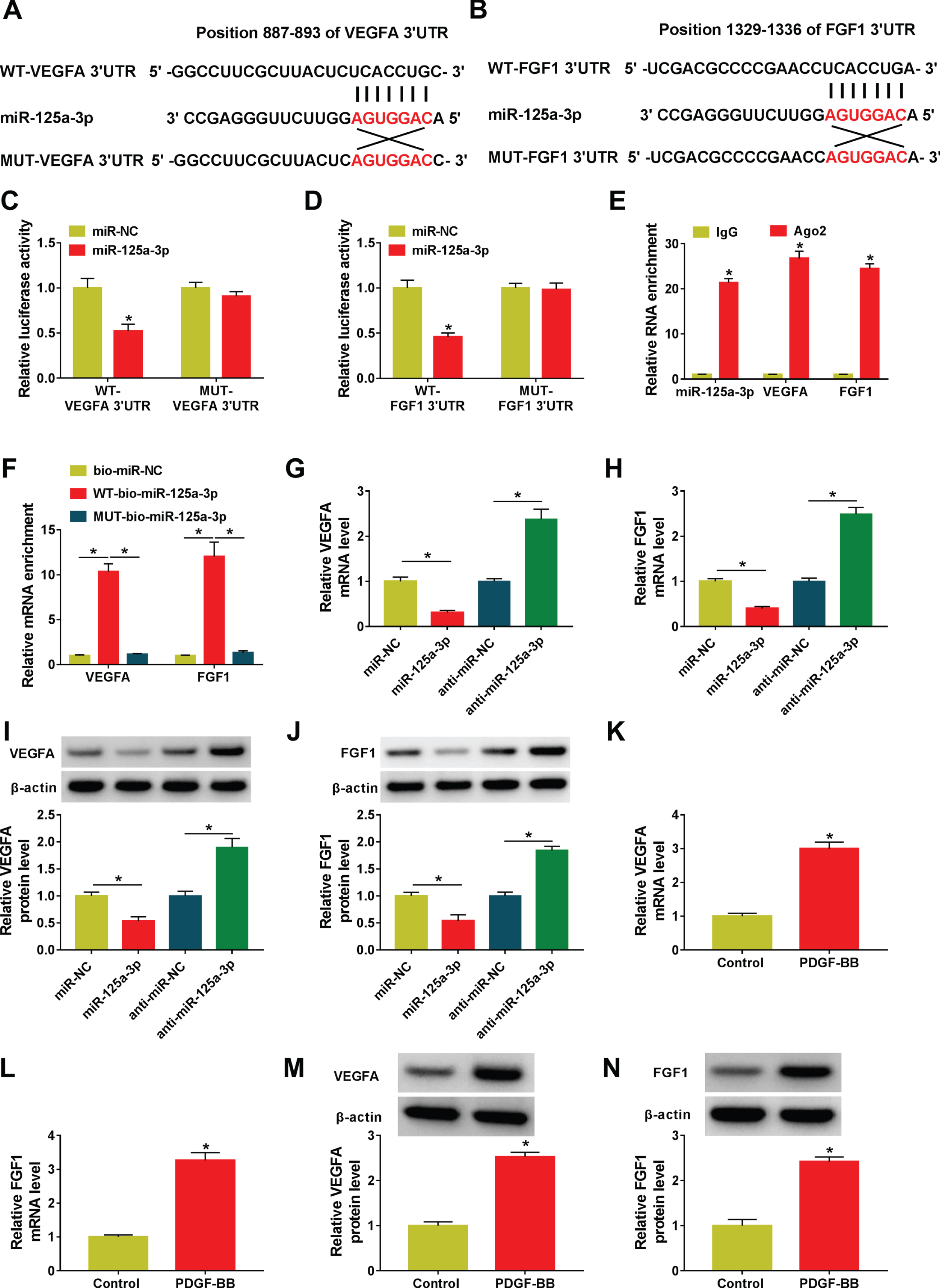
VEGFA and FGF1 served as the targets of miR-125a-3p. (A and B) The latent binding sites between miR-125a-3p and VEGFA 3’UTR or FGF1 3’UTR were predicted using targetscan software. (C and D) The prediction was proved by a dual-luciferase reporter assay. (Student’s t-test). (E and F) The interactions were verified by using RIP (Student’s t-test) and RNA pull-down assay (ANOVA with Tukey’s tests). (G and H) VEGFA or FGF1 level was detected in HASMCs transfected with miR-NC, miR-125a-3p, and anti-miR-NC, and anti-miR-125a-3p. (Student’s t-test). (I and J) Protein levels of VEGFA and FGF1 were measured in miR-125a-3p or anti-miR-125a-3p-transfected HASMCs. (Student’s t-test). (K-N) VEGFA and FGF1 were tested in control or PDGF-BB-treated HASMCs. (Student’s t-test). *P < 0.05.
Considering the regulatory role of miR-125a-3p in VEGFA or FGF1 expression in HASMCs, whether the effect of miR-125a-3p in PDGF-BB-stimulated proliferation and migration of HASMCs was correlative with VEGFA or FGF1 was further investigated. Firstly, the overexpression efficiency of pcDNA-VEGFA or pcDNA-FGF1 was verified by RT-qPCR (Fig. 6A and 6B) and western blot assays (Fig. 6C and 6D). And these transfection efficiencies were successful for the following studies. Functional analysis suggested that VEGFA or FGF1 upregulation weakened the negative action of miR-125a-3p mimic on cell viability and cell cycle progression in PDGF-BB-treated HASMCs (Fig. 6E and 6F). Synchronously, decreased migratory ability due to miR-125a-3p overexpression was significantly abated after the transfection of pcDNA-VEGFA or FGF1 in PDGF-BB-stimulated HASMCs (Fig. 6G and 6H). Consistently, the changes of PCNA, Cyclin D1, MMP2, and OPN under the same conditions further verified the influence of miR-125a-3p in PDGF-BB-stimulated proliferation and migration of HASMCs was mediated by VEGFA or FGF1 (Fig. 6I). These findings indicated that miR-125a-3p regulated PDGF-BB-stimulated proliferation and migration of HASMCs by interacting with VEGFA or FGF1.

MiR-125a-3p alleviated the PDGF-BB-induced proliferation and migration via targeting VEGFA and FGF1. (A-D) Overexpression efficiency of pcDNA-VEGFA or pcDNA-FGF1FGF1 in HASMCs was assessed. (Student’s t-test). (E-I) HASMCs were treated with PDGF-BB + miR-NC, PDGF-BB + miR-125a-3p, PDGF-BB + miR-125a-3p + pcDNA, PDGF-BB + miR-125a-3p + VEGFA, and PDGF-BB + miR-125a-3p + FGF1. (E and F) Cell viability and cell cycle progression were examined in treated HASMCs. (ANOVA with Tukey’s tests). (G and H) Migration was detected in treated HASMCs. (ANOVA with Tukey’s tests). (I) Protein levels of PCNA, Cyclin D1, MMP2, and OPN of treated HASMCs were determined. (ANOVA with Tukey’s tests). *P < 0.05.
Based on the above findings, we speculated that circLMF1 played the carcinogenic role partly by modulating the miR-125a-3p/VEGFA or FGF1 axis in HASMCs. To further verify the inference, we carried out the rescue assays. As shown in Fig. 7A-7D, the deficiency of circLMF1-1 constrained the mRNA level and protein level of VEGFA or FGF1 in HASMCs, whereas the downregulation of miR-125a-3p significantly ameliorated the inhibitory effect of si-circLMF1-1 on VEGFA or FGF1 expression. That was to say, circLMF1 served as a molecular sponge to sequester miR-125a-3p from VEGFA or FGF1.

VEGFA and FGF1 expression were regulated through circLMF1/miR-125a-3p in HASMCs. HASMCs were transfected with si-NC, si-circLMF1-1, si-circLMF1-1 + anti-miR-NC, and si-circLMF1-1 + anti-miR-125a-3p. (A and B) Relative mRNA levels of VEGFA and FGF1 were determined in transfected HASMCs. (ANOVA with Tukey’s tests). (C and D) Protein levels of VEGFA and FGF1 were detected in transfected HASMCs. (ANOVA with Tukey’s tests). *P < 0.05.
There is increasing evidence that circRNAs play a key role in a variety of human diseases, including atherosclerosis [21]. Owing to the high consecration and relative stability, circRNAs have become a hot research area in recent decades [22]. Although more than 100,000 circRNA have been identified, the function of some remains unclear and still needs to be further explored in atherosclerosis. In this study, we found that circLMF1was upregulated in proliferating conditions of HASMCs by using microarray analysis. Since PDGF-BB could rapidly reprogram cells from a differentiated contractile and non-proliferating to a dedifferentiated synthetic and proliferating stage, HASMCs were stimulated by PDGF-BB to establish a cell model in vitro in this study. Our results verified the high expression of circLMF1 in PDGF-BB- stimulated HASMCs, implying that the dysregulation of circLMF1 might be associated with PDGF-BB-stimulated HASMCs phenotype. Furthermore, the results from the present work verified that circLMF1 served as a promoting biomarker due to the stable loop structure and copious cytoplasm. CircLMF1 deficiency was affirmed to hinder PDGF-BB-stimulated proliferation and migration of HASMCs, suggesting that circLMF1 might be involved in the process of atherosclerosis.
It has been widely accepted that circRNAs mainly function as miRNA sponges to participate in a variety of biological activities [23, 24]. In the current study, we first verified that circLMF1 interacted with miR-125a-3p in HASMCs. MiR-125a-3p, a well-known tumor suppressor, has been confirmed to block metastasis of hemangioma endothelial cells [25]. Moreover, a recent study has reported the repression of miR-125a-3p in the growth and migration of VSMC [26]. Consistent with these works, we found that PDGF-BB induced an increase of miR-125a-3p expression in HASMCs. Meanwhile, functional analysis verified that inhibition of miR-125a-3p partly reversed the si-circLMF1-caused decrease in PDGF-BB-stimulated proliferation and migration of HASMCs. MiRNAs regulated cellular effects through interacting with multiple target genes. Recent literature indicated that the targets of miR-125a-3p, such as FUT5 and FUT6, Smad4 and Jak1, and MAPK, have been proved in some diseases [27–29]. In this work, VEGFA and FGF1 were identified as the downstream target genes of miR-125a-3p for the first time. VEGFA, a vascular endothelial growth factor, has been demonstrated to play a significant role in angiogenesis [30]. Synchronously, VEGFA has been pointed out to be related to the proliferation and differentiation of smooth muscle cells [31–33]. In addition, FGF1, also known as a pro-angiogenic factor for wound repair, induced the proliferation and migration of VSMC [33, 34]. In this paper, VEGFA and FGF1 were increased in PDGF-BB-triggered HASMCs, and VEGFA or FGF1 elevation partially counteracted the suppression effect of miR-125a-3p on PDGF-BB-induced the proliferation and migration of HASMCs.
To further verify that circLMF1 as a sponge of miR-125a-3p to effect VEGFA and FGF1 expression, rescue assays were performed in HASMCs. Our results indicated that reduced miR-125a-3p partly overturned circLMF1 deficiency-mediated decrease in VEGFA and FGF1 levels in HASMCs, further supporting the regulatory role of circLMF1/miR-125a-3p/VEGFA and FGF1.
Conclusion
Taken together, these findings discovered that circLMF1 knockdown dampened PDGF-BB-induced proliferation and migration of HASMCs partly by modulating miR-125a-3p/VEGFA and FGF1, which might help to explore the novel underlying therapeutic target for atherosclerosis.
Footnotes
Acknowledgment
None
Disclosure of interest
The authors declare that they have no financial conflicts of interest
Funding
None.