
Abstract
Circular RNAs (circRNAs) are associated with the pathogenesis of human diseases, including atherosclerosis. Here, we undertook to investigate the biological role and mechanism of circRNA E3 ubiquitin-protein ligase (circ-CHFR) in atherosclerosis. The expression levels of circ-CHFR, miR-214-3p, and pregnancy-associated plasma protein A (PAPPA) were measured by real-time quantitative polymerase chain reaction (RT-qPCR) and western blot in human aorta vascular smooth muscle cells (HA-VSMCs) exposed to oxidized low-density lipoprotein (ox-LDL). Cell proliferation, migration, and invasion capabilities were assessed by 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl-2H-tetrazol-3-ium bromide (MTT), and transwell assays, respectively. The relationship between miR-214-3p and circ-CHFR or PAPPA was confirmed by dual-luciferase reporter and RNA immunoprecipitation (RIP) assays. Our data showed that circ-CHFR was upregulated in HA-VSMCs after stimulation with ox-LDL. Downregulation of circ-CHFR inhibited the proliferation, migration, and invasion of HA-VSMCs exposed to ox-LDL. Mechanistically, circ-CHFR acted as a miR-214-3p sponge, and miR-214-3p was a molecular mediator of circ-CHFR regulation in ox-LDL-stimulated HA-VSMCs. PAPPA was a miR-214-3p target, and circ-CHFR regulated the expression of PAPPA by sponging miR-214-3p. Moreover, overexpression of miR-214-3p repressed the proliferation, migration, and invasion of ox-LDL-induced HA-VSMCs by decreasing PAPPA expression. Our findings suggest that the circ-CHFR/miR-214-3p/PAPPA axis regulates ox-LDL-induced proliferation, migration, and invasion in HA-VSMCs.
List of abbreviations
pregnancy-associated plasma protein A
human aorta vascular smooth muscle cells
oxidized low density lipoprotein
RNA immunoprecipitation
Introduction
Atherosclerosis is an inflammatory disease with high morbidity and mortality [1, 2]. Proliferation of human aorta vascular smooth muscle cells (HA-VSMCs) is associated with the formation of collagen fibers and lipid-depository in atherosclerosis [3, 4]. Abnormal migration and invasion of HA-VSMCs play critical roles in the vascular remodeling in vasculopathy [5]. Furthermore, oxidized low-density lipoprotein (ox-LDL) functions as a common provocative for atherosclerosis by inducing vascular endothelium and smooth muscle injury and other pathological events [6].
Circular RNAs (circRNAs), a novel subgroup of noncoding RNAs [7], have been implicated in the pathogenesis of numerous human diseases, including atherosclerosis [8]. Some circRNAs have been identified as sponges of miRNAs, functionally liberating other transcripts targeted by that set of miRNAs [9]. For instance, circ-0044073 is upregulated in atherosclerosis and it enhances the growth and migration of HA-VSMCs by sponging miR-107 and related signaling pathways [10]. Circ-CHFR (also called hsa_circ_0029589), generated by the back-splicing of the exons of E3 ubiquitin-protein ligase (CHFR) located on chr12 (133428203-133430159), has been found to be upregulated in ox-LDL-stimulated HA-VSMCs and its silencing can effectively impede the proliferation and migration of HA-VSMCs in vitro [11]. Nonetheless, our understanding of the molecular basis underlying circ-CHFR regulation is still limited.
MiRNAs are small and conserved noncoding RNAs with ∼22 nucleotides in length that control gene expression in a post-transcriptional manner by pairing to the 3’untranslated regions (UTRs) of target mRNAs [12, 13]. Downregulation of miR-214 is required for the proliferation and invasion of tumor cells, including endometrial carcinoma cells [14], and hepatocellular carcinoma cells [15], and glioma cells [16]. Moreover, miR-214-3p is associated with the pathogenesis of atherosclerosis [17–20]. Pregnancy-associated plasma protein A (PAPPA), a zinc metalloproteinase, has been shown to play an important role in cardiovascular disease [21]. PAPPA may work as a biomarker for vulnerable plaques in patients with coronary artery disease [22].
In this study, we undertook to explore the in-depth molecular mechanism of circ-CHFR activity in ox-LDL-stimulated HA-VSMCs, with the hope that such a mechanism might provide a novel insight for atherosclerosis pathogenesis.
Materials and methods
Cell line
HA-VSMCs were purchased from the American Type Culture Collection (Rockville, MD, USA) and propagated in RPMI-1640 medium (Wisent, Shanghai, China) enriched with 10%(v/v) fetal bovine serum (FBS; GIBCO BRL, Grand Island, NY, USA), 100 U/mL penicillin, and 100 U/mL streptomycin (GIBCO BRL) at 37°C and 5%CO2. HA-VSMCs were treated with various concentrations (0, 25, 50, and 100μg/mL) of ox-LDL (Biomedical Technologies, Shanghai, China) for 48 h or 50μg/mL ox-LDL for different time frames (0, 6, 12, 24, 48 h) or 50μg/mL ox-LDL for 48 h.
RNA isolation and real-time quantitative polymerase chain reaction (RT-qPCR)
The TriQuick Reagent (Solarbio, Beijing, China) was used for total RNA extraction from HA-VSMCs. A Cytoplasmic and Nuclear RNA Purification Kit (Norgen Biotek, Thorold, Canada) was applied to extract cytoplasmic and nuclear RNA. Complementary DNA (cDNA) was prepared from 2.0μg of total RNA using a High-capacity Reverse Transcription kit (Takara, Dalian, China) for circ-CHFR and mRNA or 50 ng of total RNA using miRNA specific stem-loop primers with TaqMan Reverse Transcription kit (Applied Biosystems, Foster City, CA, USA) for miR-214-3p. RT-qPCR assay was conducted with SYBR Green Real-Time PCR Master Mix (Takara) with specific primers (Sangon, Shanghai, China) on ABI 7500 HT system (Applied Biosystems). Relative expression levels were calculated based on the 2–ΔΔCt method and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or small nuclear RNA U6. The primers were displayed as follows: circ-CHFR (F-5’-CTGTGCACCTGCTGCTTC-3’; R-5’-CGCCTGCCTTCTGTACTCAG-3’); miR-214-3p (F-5’-GCCGAGACAGCAGGCACAG-3’; R-5’-CAGTGCGTGTCGTGGAGT-3’); PAPPA (F-5’-GCCCACACTGCTCTACCTC-3’; R-5’-GGCTGGCGGAAACTTGAGA-3’); GAPDH (F-5’-AGCTCACTGGCATGGCCTTC-3’; R-5’-CGCCTGCTTCACCACCTTCT-3’); U6 (F-5’-AACGCTTCACGAATTTGCGT-3’; R-5’-CTCGCTTCGGCAGCACA-3’).
Cell transfection
Transient transfection was performed using Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA) in compliance with the manufacturer’s guidelines. Small interfering RNA (siRNA) targeting circ-CHFR (si-circ-CHFR) and control siRNA (si-NC), circ-CHFR overexpression vector (circ-CHFR), PAPPA overexpression vector (PAPPA) and empty vector (pcDNA) were designed and constructed by RiboBio Corporation (Guangzhou, China). MiR-214-3p mimic (miR-214-3p) and miR-NC control, and miR-214-3p inhibitor (anti-miR-214-3p) and anti-miR-NC were obtained from Sangon. HA-VSMCs were cultured for 48 h after various transfections for further analyses. The sequences of si-circ-CHFR were: si-circ-CHFR sense: 5’-UAUGGCUGGACUGCUGAGGGG-3’; antisense, 5’- CCUCAGCAGUCCAGCCAUACG-3’.
3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl-2H-tetrazol-3-ium bromide (MTT) assay
Cell proliferation was analyzed through MTT assay. In brief, HA-VSMCs were plated into 96-well culture plates (3×103 cells/well). After 48 h of transfection, 10μL of MTT solution (Dojindo Molecular Technologies, Kumamoto, Japan) was used. Following a 4-h incubation with MTT solution, 150μL of dimethyl sulfoxide (DMSO) was added into per well to dissolve formazan crystal. The proliferation capability of HA-VSMCs was determined by detecting optical density at a wavelength of 490 nm using a microplate reader (Applied Biosystems).
Transwell assay
For transwell migration analysis, 200μL of cell suspensions (1×106 cells/mL) in FBS-free media were seeded into the upper chambers of transwell chambers. The lower chamber was filled with RPMI-1640 containing 10%FBS to induce cell migration. Following the incubation at 37°C for 24 h, the cells that did not migrate were wiped out with a cotton swab, and the migratory cells were fixed and stained. The number of migratory cells was calculated in at least three randomly selected fields using a microscope (Olympus, Tokyo, Japan). For Matrigel invasion analysis, the protocols were similar to migration analysis, and Matrigel invasion analysis was conducted using transwell chambers precoated with Matrigel (BD Biosciences, San Jose, CA, USA).
Western blot
Total protein was separated by 10%sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and then electroblotted onto polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA). Membranes were blocked in 5%skim milk solution for 1 h at room temperature and incubated with the indicated primary antibodies overnight at 4°C. Subsequently, IgG secondary antibody (ab150077; 1:3000 dilution; Abcam, Cambridge, MA, USA) was used to incubate the membranes for 1 h at room temperature. Immunoreactive bands were visualized with Clarity™ Western ECL Substrate Kit (Bio-Rad, Hercules, CA, USA) and analyzed using Image LabTM Software (Bio-Rad), with GAPDH as the internal control. The primary antibodies were: anti-GAPDH (ab181602; 1:2000 dilution; Abcam), anti-ki67 (ab92742; 1:1000 dilution; Abcam), anti-CyclinD1 (ab16663; 1:1000 dilution; Abcam), anti-N-cadherin (ab18230; 1:1000 dilution; Abcam), and anti-PAPPA (ab174314; 1:1000 dilution; Abcam).
Dual-luciferase reporter assay
The miRNA-binding sites to circ-CHFR and human 3’UTRs were predicted by the bioinformatics database starBase v2.0 (http://starbase.sysu.edu.cn/). To construct WT-circ-CHFR and PAPPA 3’UTR-WT report vectors, the partial segments of circ-CHFR and PAPPA 3’UTR containing the miR-214-3p binding sites were amplified and cloned into the pMIR-REPORTTM vector (Realgene, Nanjing, China). The mutant-type luciferase reporters MUT-circ-CHFR and PAPPA 3’UTR-MUT were generated by the mutagenesis kit (Toyobo, Osaka, Japan). HA-VSMCs were seeded in 24-well plates at 5×104 cells per well and allowed to adhere. After that, 40 ng of individual luciferase reporter and 40 pmol of miR-214-3p mimic or miR-NC were co-transfected into HA-VSMCs. After 48 h, the cells were collected for luciferase activity assay using Dual-Luciferase Reporter Assay System (GeneCopoeia, Rockville, MD, USA).
RNA immunoprecipitation (RIP) assay
The Imprint® RNA immunoprecipitation kit (Sigma, St. Louis, MO, USA) was applied for the RIP assay. HA-VSMCs (2×107) were lysed by radio Immunoprecipitation (RIPA) buffer supplemented with RNase inhibitor. The lysates were incubated with magnetic beads-conjugated with Argonaute-2 (Ago2, Millipore) or IgG (Millipore) antibody at 4°C overnight. The co-precipitated RNAs were treated with proteinase K buffer and then subjected to RT-qPCR assay.
Statistical analysis
Data were displayed as the mean±SD from at least three independent experiments. All statistical analyses were performed using the SPSS 21.0 software (IBM, Somers, NY, USA), and P < 0.05 was considered significant. Significant differences between two groups were assessed by a Student’s t-test, and one-way analysis of variance was applied for the comparison of multiple groups.
Results
Circ-CHFR expression was increased and miR-214-3p expression was decreased in ox-LDL-stimulated HA-VSMCs
As demonstrated by RT-qPCR, ox-LDL stimulation induced circ-CHFR expression in HA-VSMCs in dose- and time-dependent manners (Fig. 1A-1B). Conversely, ox-LDL reduced miR-214-3p expression in HA-VSMCs in dose- and time-dependent manners (Fig. 1C-1D). Additionally, to examine the localization of circ-CHFR, we adopted subcellular fractionation assays. Circ-CHFR was mainly located at the cytoplasm of HA-VSMCs (Fig. 1E).

The expression levels of circ-CHFR and miR-214-3p in HA-VSMCs exposed to ox-LDL. (A-B) The expression level of circ-CHFR in HA-VSMCs treated with ox-LDL (different dose and time) was measured by RT-qPCR analysis. (C-D) RT-qPCR was used to analyze miR-214-3p level in HA-VSMCs exposed to ox-LDL. (E) The expression level of circ-CHFR in nucleus and cytoplasm was detected by RT-qPCR analysis. N = 3, *P < 0.05.
To direct elucidate the role of circ-CHFR, we performed in vitro loss-of-function experiments in ox-LDL-stimulated HA-VSMCs. Transient transfection of si-circ-CHFR, but not si-NC controls, remarkably decreased circ-CHFR expression in HA-VSMCs exposed to ox-LDL (Fig. 2A). Notably, knockdown of circ-CHFR reduced cell proliferation induced by ox-LDL in HA-VSMCs (Fig. 2B). Moreover, silencing of circ-CHFR reversed ox-LDL-induced migration and invasion (Fig. 2C-2D). Additionally, ox-LDL stimulation led to enhanced expression levels of ki67, CyclinD1, and N-cadherin in HA-VSMCs, and knockdown of circ-CHFR abrogated the enhancement effects (Fig. 2E). Collectively, the data demonstrated that knockdown of circ-CHFR reduced ox-LDL-induced proliferation, migration, and invasion in HA-VSMCs.
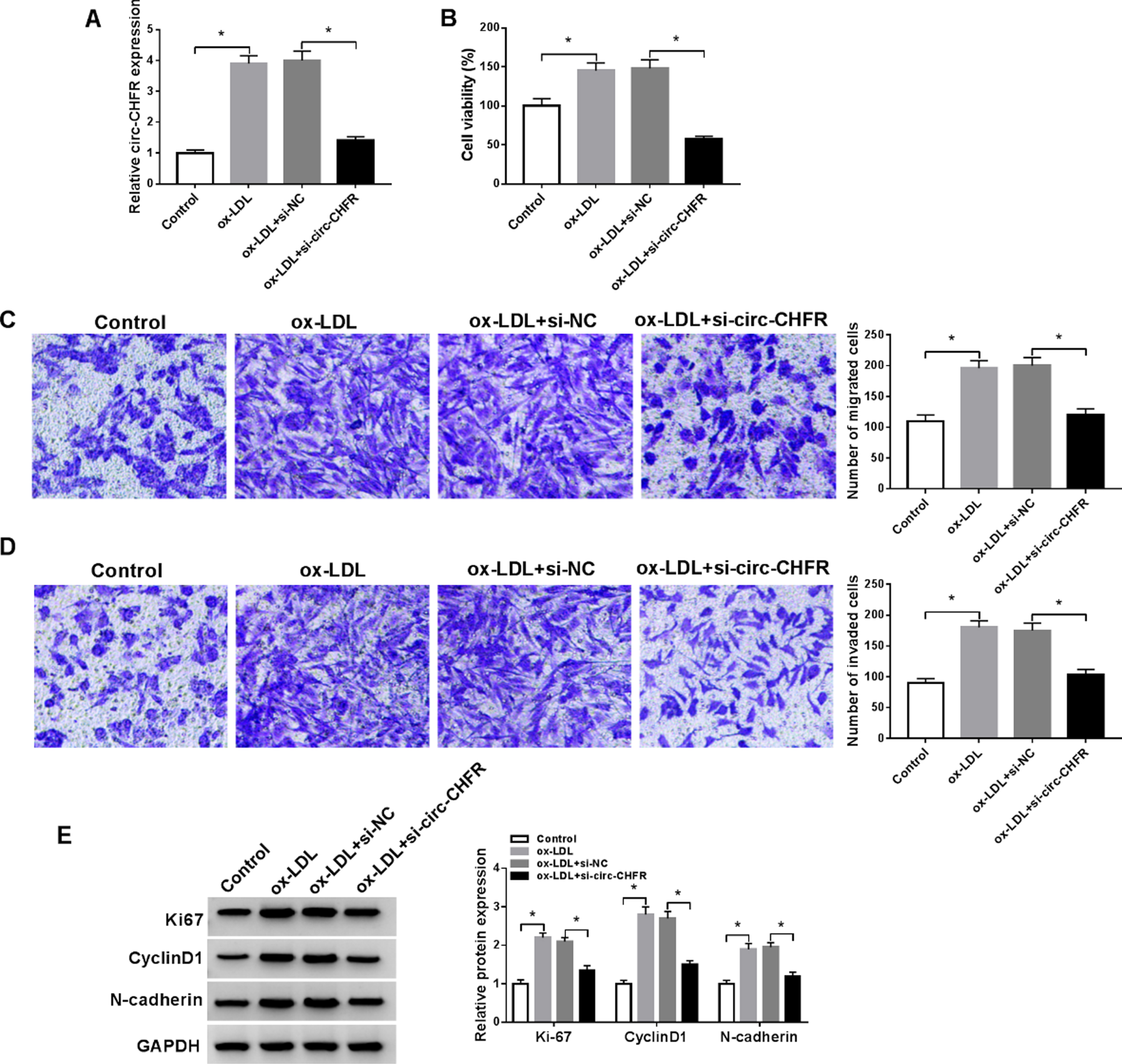
Knockdown of circ-CHFR reduced cell proliferation, migration, and invasion of HA-VSMCs caused by ox-LDL. (A-E) HA-VSMCs were separated into four groups: Control, ox-LDL, ox-LDL+si-NC, and ox-LDL+si-circ-CHFR. (A) The expression level of circ-CHFR was evaluated in HA-VSMCs by RT-qPCR assay. (B) The proliferation capability of HA-VSMCs was measured by MTT assay. (C-D) Representative pictures and migration and invasion abilities of HA-VSMCs were presented by transwell assay. (E) Western blot analysis was implemented to test ki67, CyclinD1, and N-cadherin levels in HA-VSMCs. N = 3, *P < 0.05.
We then determined the mechanism by which circ-CHFR regulated the proliferation, migration, and invasion in ox-LDL-induced HA-VSMCs. As shown in Fig. 3A, bioinformatic analysis showed that circ-CHFR had a putative binding region for miR-214-3p. Co-transfection of the wild-type luciferase reporter (WT-circ-CHFR) and miR-214-3p mimic into HA-VSMCs generated lower luciferase activity than cells co-transfected with miR-NC controls but mutation of the seed region (MUT-circ-CHFR) abolished the repression of miR-214-3p (Fig. 3B). RIP assays revealed that circ-CHFR and miR-214-3p were synchronously enriched by an anti-Ago2 antibody compared with IgG controls (Fig. 3C). Next, we evaluated whether circ-CHFR could affect miR-214-3p expression. Transfection of circ-CHFR expressing plasmid into HA-VSMCs elevated circ-CHFR expression in ox-LDL-stimulated HA-VSMCs (Fig. 3D). Importantly, in ox-LDL-stimulated HA-VSMCs, silencing of circ-CHFR by si-circ-CHFR transfection resulted in increased expression of miR-214-3p, while overexpression of circ-CHFR inhibited miR-214-3p expression (Fig. 3E).
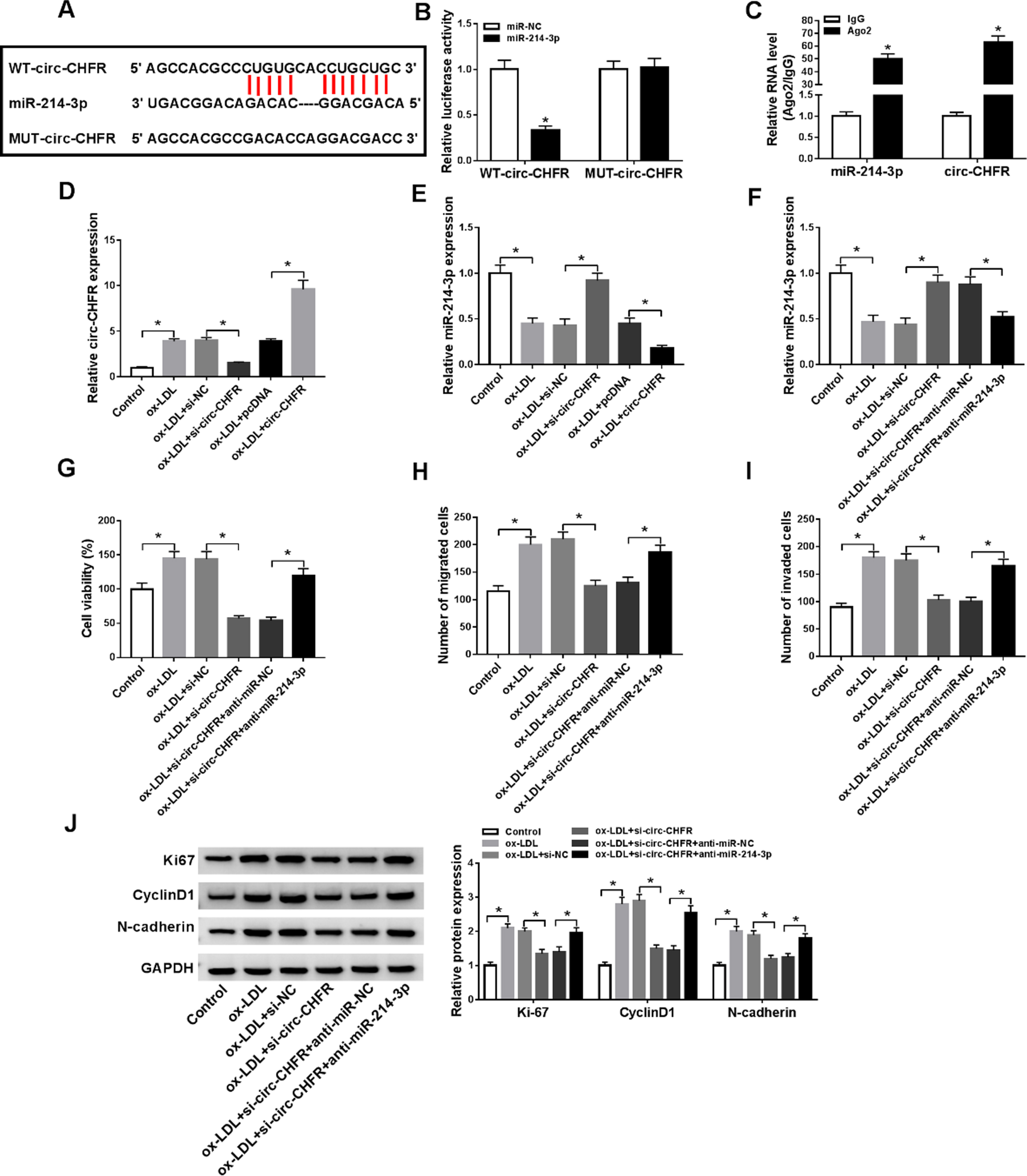
Circ-CHFR regulated the proliferation, migration, and invasion of ox-LDL-induced HA-VSMCs by miR-214-3p. (A) Binding regions between miR-214-3p and circ-CHFR, and the mutant form of circ-CHFR were shown. (B) The relative luciferase activities of WT-circ-CHFR and MUT-circ-CHFR were examined by dual-luciferase reporter assay. (C) The abundances of circ-CHFR and miR-214-3p in Ago2 and IgG groups were measured by RT-qPCR assay following RIP assay. (D-E) The expression levels of circ-CHFR and miR-214-3p were quantified by RT-qPCR assay in HA-VSMCs treated with ox-LDL, ox-LDL+si-NC, ox-LDL+si-circ-CHFR, ox-LDL+pcDNA, or ox-LDL+circ-CHFR, with untreated cells as Control. (F-J) HA-VSMCs were divided into six groups: Control, ox-LDL, ox-LDL+si-NC, ox-LDL+si-circ-CHFR, ox-LDL+si-circ-CHFR+anti-miR-NC, ox-LDL+si-circ-CHFR+anti-miR-214-3p. (F) RT-qPCR assay was used to measure miR-214-3p level in HA-VSMCs. (G) MTT assay was conducted to evaluate the cell viability of HA-VSMCs. (H-I) Transwell migration and invasion assays were performed in HA-VSMCs. (J) The protein expression levels of ki67, CyclinD1, and N-cadherin in HA-VSMCs were evaluated by western blot assay. N = 3, *P < 0.05.
To directly test whether the effects of circ-CHFR silencing were due to the upregulation of miR-214-3p, we suppressed miR-214-3p expression in si-circ-CHFR-transfected HA-VSMCs under ox-LDL (Fig. 3F). Remarkably, knockdown of miR-214-3p rescued circ-CHFR silencing-imposed cell viability defect in ox-LDL-induced HA-VSMCs (Fig. 3G). Consistently, transwell migration and invasion assays indicated that knockdown of miR-214-3p abrogated circ-CHFR silencing-driven inhibition of migration and invasion of ox-LDL-induced HA-VSMCs (Fig. 3H-3I). Moreover, knockdown of miR-214-3p reversed the suppression effects of circ-CHFR downregulation on ki67, CyclinD1, and N-cadherin expression levels in ox-LDL-induced HA-VSMCs (Fig. 3J). Together, miR-214-3p seemed to be a downstream mediator of circ-CHFR regulation in ox-LDL-induced HA-VSMCs.
Using starbase software, PAPPA was predicted to have a putative complementary site for miR-214-3p (Fig. 4A). Transfection of miR-214-3p mimic decreased the luciferase activity of PAPPA 3’UTR luciferase reporter (PAPPA 3’UTR-WT) but not the mutation in the seed region (PAPPA 3’UTR-MUT) (Fig. 4B), indicating that PAPPA was a miR-214-3p target. RIP experiments revealed that PAPPA mRNA and miR-214-3p were significantly enriched in the anti-Ago2 antibody group compared with the control group (Fig. 4C). Importantly, ox-LDL induced the expression of PAPPA at both mRNA and protein in HA-VSMCs in dose- and time-dependent manners (Fig. 4D-4G). We then assessed the ability of miR-214-3p to regulate PAPPA expression. The transfection efficiencies of miR-214-3p mimic and anti-miR-214-3p were validated by RT-qPCR (Fig. 4H). As expected, overexpression of miR-214-3p inhibited PAPPA expression at both mRNA and protein in ox-LDL-induced HA-VSMCs; conversely, knockdown of miR-214-3p led to elevated levels of PAPPA mRNA and protein (Fig. 4I-4J). All these data indicated that miR-214-3p negatively regulated PAPPA expression in HA-VSMCs exposed to ox-LDL.

MiR-214-3p targeted PAPPA in HA-VSMCs exposed to ox-LDL. (A) By bioinformatics database prediction, miR-214-3p had the binding region in 3’UTR of PAPPA. (B) The association between miR-214-3p and PAPPA was analyzed by dual-luciferase reporter assay. (C) RIP assay was conducted to assess the level of miR-214-3p and PAPPA in Ago2 and IgG group from cell lysates of HA-VSMC. (D-G) The expression levels of PAPPA were determined by RT-qPCR and western blot assays in HA-VSMCs treated with ox-LDL for different doses or times. (H) RT-qPCR was utilized to show the miR-214-3p level in HA-VSMCs treated with ox-LDL, ox-LDL+miR-NC, ox-LDL+miR-214-3p, ox-LDL+anti-miR-NC, ox-LDL+anti-miR-214-3p, with untreated cells as Control. (I-J) The mRNA and protein expression levels of PAPPA were measured by RT-qPCR and western blot assays in HA-VSMCs treated with ox-LDL, ox-LDL+miR-NC, ox-LDL+miR-214-3p, ox-LDL+anti-miR-NC, ox-LDL+anti-miR-214-3p, with untreated cells as Control. N = 3, *P < 0.05.
As shown in Fig. 5A, co-transfection of PAPPA expressing plasmid significantly abrogated miR-214-3p overexpression-mediated inhibition of PAPPA expression in ox-LDL-induced HA-VSMCs. Moreover, miR-214-3p overexpression suppressed cell viability in ox-LDL-induced HA-VSMCs, and this effect was abolished by restoration of PAPPA (Fig. 5B). Overexpression of miR-214-3p led to repressed migration and invasion rates in ox-LDL-induced HA-VSMCs, which was abolished by restoration of PAPPA (Fig. 5C-5D). Furthermore, ox-LDL-induced HA-VSMCs expressing miR-214-3p exhibited lower levels of Ki-67, CyclinD1, and N-cadherin compared with controls, and restoration of PAPPA reversed these impacts (Fig. 5E). In summary, miR-214-3p inhibited the proliferation, migration, and invasion in ox-LDL-induced HA-VSMCs by repressing PAPPA expression.
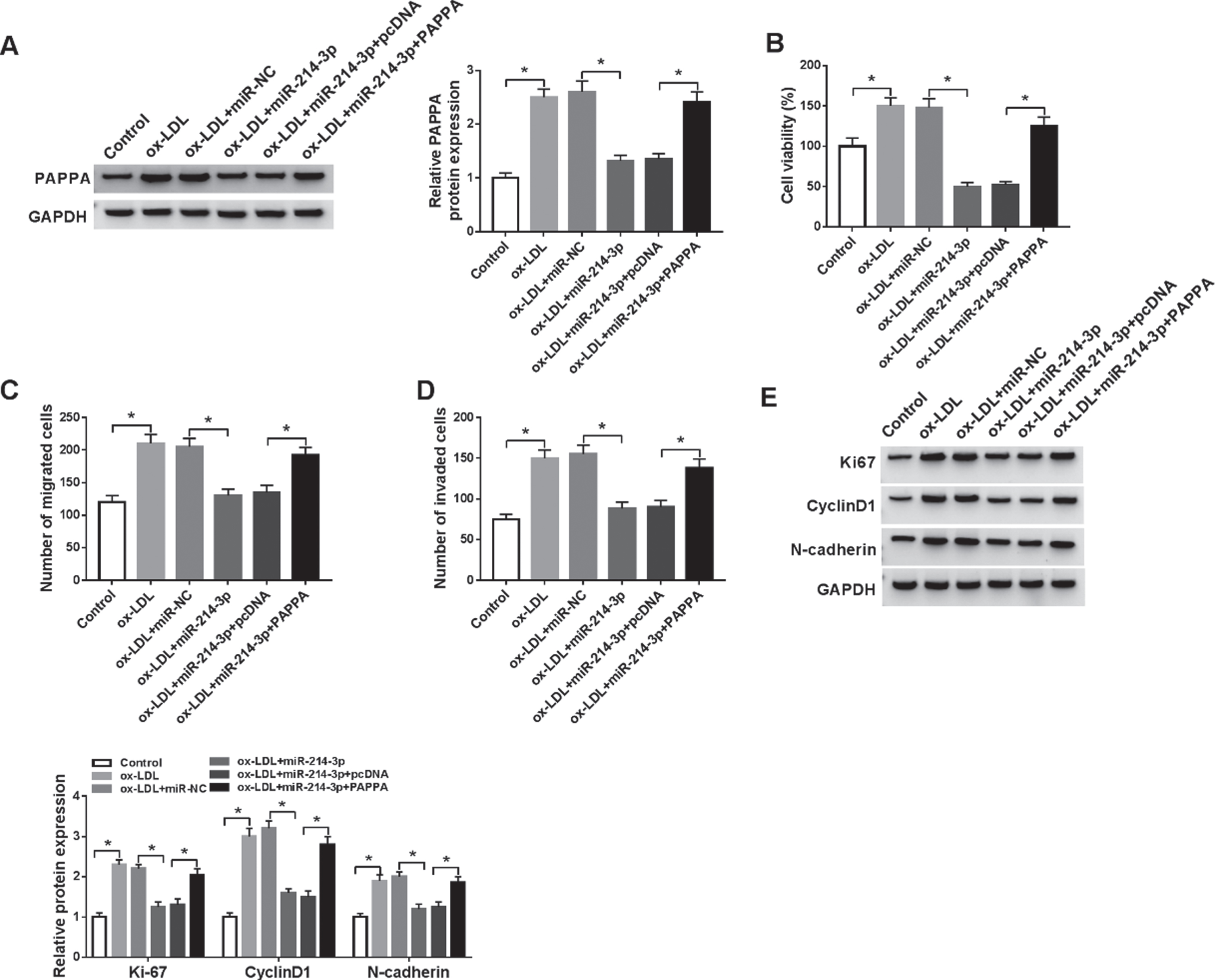
MiR-214-3p regulated proliferation, migration, and invasion in ox-LDL-induced HA-VSMCs by affecting PAPPA. (A-E) HA-VSMCs were treated with ox-LDL, ox-LDL+miR-NC, ox-LDL+miR-214-3p, ox-LDL+miR-214-3p+pcDNA, ox-LDL+miR-214-3p+PAPPA, with untreated cells as Control. (A) The western blot assay was carried out to test PAPPA level in HA-VSMCs. (B) Cells viability of HA-VSMCs was measured by MTT assay. (C-D) Transwell analysis was introduced to show the migration and invasion abilities of HA-VSMCs. (E) The protein expression levels of ki67, CyclinD1, and N-cadherin were quantified by western blot assay in HA-VSMCs. N = 3, *P < 0.05.
Having established the direct relationship between miR-214-3p and circ-CHFR or PAPPA, we further determined whether circ-CHFR could affect PAPPA expression via miR-214-3p. Intriguingly, knockdown of circ-CHFR resulted in reduced expression of PAPPA at both mRNA and protein in ox-LDL-induced HA-VSMCs, and silencing of miR-214-3p reversed the effect (Fig. 6A-6B). Conversely, overexpression of circ-CHFR enhanced the expression of PAPPA in ox-LDL-induced HA-VSMCs, which was abrogated by overexpression of miR-214-3p (Fig. 6C-6D). These data together suggested that circ-CHFR regulated PAPPA expression by miR-214-3p.

Circ-CHFR regulated PAPPA expression through miR-214-3p. (A-B) The mRNA and protein expression levels of PAPPA were quantified by RT-qPCR and western blot assays in HA-VSMCs treated with ox-LDL, ox-LDL+si-NC, ox-LDL+si-circ-CHFR, ox-LDL+si-circ-CHFR+anti-miR-NC, ox-LDL+si-circ-CHFR+anti-miR-214-3p, with untreated cells as Control. (C-D) The expression levels of PAPPA were measured by RT-qPCR and western blot assays in HA-VSMCs treated with ox-LDL, ox-LDL+pcDNA, ox-LDL+circ-CHFR, ox-LDL+circ-CHFR+miR-NC, ox-LDL+circ-CHFR+miR-214-3p, with untreated cells as Control. N = 3, *P < 0.05.
In this study, we validated that circ-CHFR expression was elevated and miR-214-3p expression was declined in HA-VSMCs exposed to ox-LDL. Functionally, knockdown of circ-CHFR could attenuate ox-LDL-induced proliferation, migration, and invasion of HA-VSMCs, in line with a previous report [11]. Mechanically, circ-CHFR acted as a molecule sponge of miR-214-3p, and miR-214-3p targeted and inhibited PAPPA. Our findings uncovered the vital role of the circ-CHFR/miR-214-3p/PAPPA axis in HA-VSMCs treated with ox-LDL.
Ox-LDL has been considered as a stimulus for atherosclerosis through multiple mechanisms, including the regulation of HA-VSMC behavior [23]. Moreover, HA-VSMCs are highly associated with vascular wall repair after injury and pro-atherosclerotic switch [24]. Consistent with previous findings [25], we also confirmed that ox-LDL enhanced the proliferation of HA-VSMCs. Ki67, a nuclear antigen, is closely associated with cell division and proliferation; the abnormal expression of Ki67 can reflect cell proliferation alteration in tumor cells [26]. Not surprisingly, the expression of Ki67 was higher in ox-LDL-induced HA-VSMCs than control cells. Considering cell proliferation is related to mitosis, the expression of CyclinD1 was measured. CyclinD1 can bind to cyclin-dependent kinase to form a complex, thereby regulating cell cycle [27]. The previous report suggested that Isoliquiritigenin suppressed the proliferation of HA-VSMCs and arrested HA-VSMCs in the G1/S phase by decreasing CyclinD1 expression [28]. The migration of HA-VSMCs is significantly correlated with the occurrence and development of intimal hyperplasia and atherosclerosis [29]. The upregulation of N-cadherin is conducive to the migration of HA-VSMCs during atherosclerosis [30]. We unveiled that the expression of N-cadherin was higher in ox-LDL-induced HA-VSMCs than in control cells.
Circ-CHFR is upregulated in HA-VSMCs treated with ox-LDL and it can enhance cell proliferation and migration in vitro [11], implying that circ-CHFR may be involved in the development of atherosclerosis. Accumulating evidence shows that circRNAs play key roles in the pathogenesis of atherosclerosis by sponging miRNAs [31, 32]. In this study, we first demonstrated that circ-CHFR regulated miR-214-3p expression by binding to miR-214-3p. MiR-214 has been identified as a potential diagnosis and therapeutic target for cardiovascular diseases [33]. Liu et al. confirmed that knockdown of LINC00341 suppressed the growth and invasion abilities of VSMCs by regulating the miR-214/Forkhead box O4 axis [34]. We first verified that miR-214-3p expression was decreased in ox-LDL-induced HA-VSMCs. Gain-of-function experiments further showed that upregulation of miR-214-3p resulted in the dramatic suppression of proliferation, migration, and invasion in ox-LDL-induced HA-VSMCs. Analogously, Afzal et al. reported that upregulation of miR-214 decreased the proliferation and migration of VSMCs via the regulation of NCK-associated protein 1 [35]. Importantly, we first uncovered that knockdown of circ-CHFR impeded the proliferation, proliferation and mobility of ox-LDL-stimulated HA-VSMCs by increasing miR-214-3p expression.
MiRNAs silence gene expression by binding to the 3’UTRs of target mRNAs [36]. PAPPA is able to promote the growth of VSMCs and the development of atherosclerosis [37]. Here, we first established that miR-214-3p targeted and inhibited PAPPA. Additionally, we further validated the increase of PAPPA expression in HA-VSMCs exposed to ox-LDL. Analogously, miR-141 might also be involved in the regulation of atherosclerosis by targeting PAPPA [38]. More importantly, we first ascertained that circ-CHFR regulated PAPPA expression through miR-214-3p, suggesting the implication of the circ-CHFR/miR-214-3p/PAPPA axis in ox-LDL-treated HA-VSMCs. Similarly, Yang et al. reported that circ-CHFR promoted the growth and motility of ox-LDL-treated HA-VSMCs depending on the modulation of the miR-370/FOXO1 axis [11]. Therefore, we hypothesized that the circ-CHFR/miR-214-3p/PAPPA and circ-CHFR/miR-370/FOXO1 axes might be two interactional or paralleled networks in the regulation of ox-LDL-treated HA-VSMCs. PAPPA is considered as a detrimental factor in atherosclerosis, and the in-depth mechanism by which PAPPA contributes to atherosclerosis pathogenesis is worth exploring in the further study [39].
Conclusions
Collectively, knockdown of circ-CHFR could reduce ox-LDL-induced growth, motility and invasion of HA-VSMCs. Mechanistically, circ-CHFR worked as a sponge for miR-214-3p to upregulate PAPPA in ox-LDL-stimulated HA-VSMCs. Our findings suggest that circ-CHFR affects the proliferation, migration, and invasion of ox-LDL-induced HA-VSMCs via the miR-214-3p/PAPPA axis.
Footnotes
Acknowledgments
None
Disclosure of interest
The authors declare that they have no financial conflicts of interest
Funding
There is no funding support.
Ethical approval
The conducted research is not related to either human or animal use.