
Abstract
BACKGROUND:
Previous data have shown that circ_0033596 is involved in the pathogenesis of atherosclerosis (AS). The study aims to reveal the detailed mechanism of circ_0033596 in AS.
METHODS:
Human umbilical vein endothelial cells (HUVECs) were treated with oxidized low-density lipoprotein (ox-LDL) to establish an AS cell model. Quantitative real-time polymerase chain reaction and western blot were implemented to detect the expression of circ_0033596, miR-637, growth factor receptor bound protein2 (GRB2), BCL2-associated x protein (Bax) and B-cell lymphoma-2 (Bcl-2). Cell viability, proliferation, apoptosis and tube formation were investigated by cell counting kit-8, EdU assay, flow cytometry and tube formation assay, respectively. The production of interleukin (IL-6) and tumor necrosis factor-α (TNF-α) was evaluated by enzyme-linked immunosorbent assay. Oxidative stress was evaluated by lipid peroxidation malondialdehyde assay kit and superoxide dismutase activity assay kit. Dual-luciferase reporter assay, RNA pull-down assay and RIP assay were performed to identify the associations among circ_0033596, miR-637 and GRB2.
RESULTS:
The expression of circ_0033596 and GRB2 was significantly increased, while miR-637 was decreased in the blood of AS patients and ox-LDL-induced HUVECs compared with controls. Ox-LDL treatment inhibited HUVEC viability, proliferation and angiogenic ability and induced cell apoptosis, inflammation and oxidative stress, while these effects were attenuated after circ_0033596 knockdown. Circ_0033596 interacted with miR-637 and regulated ox-LDL-induced HUVEC damage by targeting miR-637. In addition, GRB2, a target gene of miR-637, participated in ox-LDL-induced HUVEC injury by combining with miR-637. Importantly, circ_0033596 activated GRB2 by interacting with miR-637.
CONCLUSION:
Circ_0033596 depletion protected against ox-LDL-induced HUVEC injury by miR-637/GRB2 pathway, providing a therapeutic target for AS.
Keywords
Introduction
Atherosclerosis (AS) mainly includes coronary heart disease, stroke and peripheral arterial disorder, featured by lipid accumulation as well as fibrous cap formation [1]. AS is a life-long illness and contributes to the occurrence of cardiovascular disease [2]. Atherosclerotic lesion formation involves different types of cells like endothelial cells and macrophages [3]. As reported, AS occurrence involves endothelial damage, inflammatory reaction, cell apoptosis and oxidative stress [4, 5]. Exposure to AS risk factors stimulates the endothelium to express adhesion molecules and cytokines such as monocyte chemoattractant protein-1, further recruiting circulating monocytes to the lesion and differentiate into macrophages, which in turn scavenge lipids to form foam cells [6]. Oxidized low-density lipoprotein (ox-LDL) has pro-inflammatory and immune-stimulatory properties and can activate endothelial cells, thus playing an important role in AS [7]. In addition, previous data have demonstrated that ox-LDL treatment can activate human umbilical vein endothelial cells (HUVECs) in AS [8]. Thus, the study of the detailed mechanism underlying the abnormal biological behaviors of HUVECs in AS is important for AS therapy.
In recent years, experimental data have suggested that noncoding RNAs combine with coding RNAs to regulate endothelial cell injury during AS [9–11]. CircRNA is a RNA generated through head-to-tail splicing [12]. As reported, circRNA is responsible for multiple biological processes, and its dysregulation is involved in complicated diseases such as cardiovascular disease [13] and cancers [14]. Recently, some researchers reported that circRNAs also participated in AS occurrence. For example, circ_102541 was upregulated in AS specimens and combined with miR-296-5p to promote HUVEC proliferation in vitro [15]. Additionally, circPTPRA depletion repressed vascular smooth muscle cell (VSMC) growth by combining with the miR-636/Sp1 transcription factor (SP1) pathway in AS [16]. Another circRNA, circ_0033596, has been reported to aggravate ox-LDL-induced endothelial cell damage in AS [17]; however, the regulatory mechanism of the circRNA in atherosclerosis remains to be largely explored.
MicroRNA (miRNA) can inhibit gene expression by inducing the degradation of mRNA or the inhibition of the translation process [18]. It has been accepted that endothelial cell dysfunction is an initial induction factor of AS occurrence [19]. MiRNA, as it influents endothelial cell function, must be considered to be associated with the impacts of the monolayer [20]. Considerable studies have revealed that miRNA affects endothelial cell dysfunction by exacerbating endothelial cell senescence [21]. Some miRNAs, such as miR-576 [22], miR-21 [23] and miR-217 [24], have been reported to participate in regulating mechanisms related to AS. Another miRNA, miR-637, has a high value in diagnosing AS [25]. Moreover, miR-637 introduction alleviated AS-like vascular smooth muscle cell injury induced by ox-LDL [26].
The competitive endogenous RNA hypothesis proposes transcripts can compete for shared miRNAs to regulate each other and link the functions of circRNA, miRNA and mRNA [27]. Through the prediction of online databases, we found that circ_0033596 contained the complementary sites of miR-637. Thus, we hypothesized that circ_0033596 combined with miR-637 to function in AS. In this study, we validated whether the circ_0033596/miR-637 axis was involved in atherosclerosis occurrence using ox-LDL-stimulated HUVECs. In addition, we searched for miR-637 and circ_0033596-associated mRNA(s) in ox-LDL-treated HUVECs
Materials and methods
Study population
25 AS patients and 25 healthy volunteers were recruited from the First People’s Hospital of Lianyungang, the First Affiliated Hospital of Kangda College of Nanjing Medical University for the study. The blood of these individuals was collected and stored at –80°C. This research protocol was approved by the Ethics Committee of the First People’s Hospital of Lianyungang, the First Affiliated Hospital of Kangda College of Nanjing Medical University. The written informed consents were acquired from all participants before the study. These AS patients did not receive any treatment before the study. The healthy controls were registered according to the published standards [28].
Cell culture and treatment
HUVECs were purchased from Otwo Biotech (Shenzhen, China) and maintained in DMEM (Biosun, Shanghai, China) plus 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin at 37°C in an atmosphere of 5% CO2. In addition, HUVECs were treated with various concentrations of ox-LDL (Yeasen Biotech, Shanghai, China; 0, 25, 50 and 75μg/mL) for 24 h to induce AS-like cell injury. HUVECs from passages 1 to 5 were used in the study.
Cell transfection
As per the guidebook, plasmids and oligonucleotides were transfected into HUVECs at 50–75% confluence with FuGENE6 (Roche, Basel, Switzerland). Further study was performed after 48 h of cell transfection. Ribobio Co., Ltd. (Guangzhou, China) provided the small interfering RNA targeting circ_0033596 (si-circ_0033596, 5’-CAGCATGGCTGAGGTTGTGCA-3’), the mimics and inhibitors of miR-637 (miR-637, 5’-ACUGGGGGCUUUCGGGCUCUGCGU-3’ and anti-miR-637, 5’-ACGCAGAGCCCGAAAGCCCCCAGU-3’) as well as controls (si-NC, miR-NC, and anti-miR-NC). The overexpression plasmid of growth factor receptor bound protein2 (GRB2) was built by Geneseed Co., Ltd. (Guangzhou, China) using pcDNA 3.0 (+) vector.
Quantitative real-time polymerase chain reaction (qRT-PCR)
RNA isolation from blood samples and cells was performed using TsingZol (Tsingke, Shanghai, China). cDNA synthesis was conducted according to the instruction of FastKing RT reagents (Tiangen, Beijing, China) or cDNA Synthesis Kit (Tsingke). Then, Fast qPCR Mix (Tsingke) was utilized to quantify the expression of circRNA/miRNA/mRNA with a qRT-PCR machine (Bio-Rad, Hercules, CA, USA). Data were analyzed by the 2–ΔΔCt method with normalization to GAPDH or U6. Primer sequences are listed in Table 1.
Primer sequences used in qRT-PCR
Primer sequences used in qRT-PCR
The circular characteristic of circ_0033596 was identified using RNase R, oligo(dT)18 primer and random primer based on the reporter methods [29]. qRT-PCR was applied to examine circ_0033596 and GAPDH expression. For circ_0033596 localization, Nuclear/Cytosol isolation reagents (Amyjet, Wuhan, China) were used to extract cytoplasmic and nuclear RNA of HUVECs, and qRT-PCR was used to measure the nucleocytoplasmic circ_0033596 expression.
Cell counting kit-8 (CCK-8) assay
HUVECs were plated in 96-well culture microplates and were cultured to 50–75% confluence. Then, the cells were incubated with test compounds including ox-LDL (0, 25, 50 and 75μg/mL), si-circ_0033596, si-NC, anti-miR-637, anti-miR-NC, miR-637, miR-NC, GRB2 and pcDNA. After 48 h of transfection, the cells were incubated with CCK-8 solution (Solarbio, Beijing, China) at 37°C for 3 h. Finally, enzyme immunoassay analyzer (Bio-Rad) was employed to detect the absorbance at 450 nm. Mock HUVECs were chosen as controls.
5-Ethynyl-29-deoxyuridine (EdU) assay
HUVECs with different treatments were cultured in 6-well plates for 48 h. Then, the cells in the logarithmic growth phase were passed into 96-well plates and incubated with EdU to analyze DNA synthesis as per the user’s manual of EdU staining kit (Ribobio). At last, EdU-positive HUVECs were analyzed with a fluorescent microscope (Olympus, Tokyo, Japan).
Flow cytometry analysis
We adopted Annexin V-FITC apoptosis detection kit (Solarbio) to analyze HUVEC apoptosis. Briefly, HUVECs in the logarithmic growth phase were harvested and centrifuged at 300 g for 12 min. Then, the cells were suspended in Binding Buffer and incubated with Annexin V-FITC and propidium iodide (PI) in the dark. After 15 min, flow cytometry was applied to analyze apoptotic cells.
Western blot analysis
Total proteins were extracted after lysing blood and cell samples using NP-40 lysis buffer containing phenylmethanesulfonyl fluoride. After denaturation at 95°C for 12 min, the proteins were separated using a Midi-Cell Electrophoresis System. The aspecific signals on nitrocellulose membranes were blocked, and the following primary antibodies including BCL2-associated x protein (Bax) (Cat# PA5-11378; 1:2000; Thermo Fisher, Waltham, MA, USA), B-cell lymphoma-2 (Bcl-2) (Cat# PA5-20068; 1:1000; Thermo Fisher), GRB2 (Cat# PA5-27151; 1:1500; Thermo Fisher) and GAPDH (Cat# PA1-988; 1:2500; Thermo Fisher) were used to incubate the membranes. Then, the membranes were probed by secondary antibodies (Thermo Fisher). Analysis for the gray values of blots was performed using image J software.
Analysis of angiogenic ability
The tube formation of HUVECs was analyzed by calculating capillary-like branches. In short, HUVECs in the logarithmic growth phase were inoculated in 35 mm petri dishes, which were pre-coated with Matrigel (Qcbio science, Shanghai, China). Forty-eight hours later, capillary-like branches from six microscope fields were analyzed under a DP71 microscope (Olympus).
Enzyme-linked immunosorbent assay (ELISA)
After 48 h of various treatments, cell supernatant was collected by centrifugation. Commercial ELISA kits (Beyotime, Shanghai, China) were applied to analyze IL-6 and TNF-α levels in accordance with the user’s manuals. In brief, the supernatants of HUVECs at the layer confluent were harvested by centrifugated at 500 g for 5 min and added to each well of 96-well plates. The cell supernatants were incubated with antibodies for 1-2 h and horseradish peroxidase-marked streptavidin for 20 min. At last, IL-6 and TNF-α levels were detected using a microplate reader (Thermo Fisher).
Analysis of oxidative stress
The assay used to analyze oxidative stress was conducted by detecting malondialdehyde (MDA) level and superoxide dismutase (SOD) activity using commercial assay kits (Beyotime). For detecting MDA level, HUVECs at the layer confluent were harvested and then mixed with MDA lysis buffer for 2 min. Cell supernatant was collected by centrifugation at 13000 g for 10 min. Afterward, thiobarbituric acid regent was added into each well for 1 h. At last, the samples were measured immediately using a microplate reader (Thermo Fisher). For the SOD activity assay, HUVECs at the layer confluent were harvested and lysed using Tris/HCl. Then, cell supernatant was collected and incubated with Enzyme Working Solution. After 30 min, the samples were analyzed using a microplate reader.
Dual-luciferase reporter assay
Online databases including Circular RNA interactome and Targetscan were applied to search for the targets of circ_0033596 and miR-637. The sequences of circ_0033596 and GRB2 3’-untranslated region (3’UTR) containing miR-637-binding sites were introduced into pmirGLO vector to build WT-circ_0033596 and WT-GRB2 3’UTR. The potential miR-637-binding sites in circ_0033596 and GRB2 3’UTR were mutated with site mutagenesis reagents (Yeasen) to build the mutant (MUT) plasmids including MUT-circ_0033596 and MUT-GRB2 3’UTR. Subsequently, HUVECs in the logarithmic growth phase were transfected with the above plasmids, miR-637 mimics or miR-NC using FuGENE6 (Roche). After 48 h, luciferase activity was determined with Dual-Lucy Assay Kit (Solarbio).
RNA pull-down assay
Biotinylated miR-637 (bio-miR-637) and control (bio-miR-NC) were synthesized by GenePharma Company (Shanghai, China). HUVECs at 70–80% confluence were transfected with bio-miR-637 and bio-miR-NC. After 48 h of culture, the HUVECs were exposed to lysis buffer and incubated with streptavidin-coupled beads (Invitrogen, Carlsbad, CA, USA). At last, circ_0033596 and GRB2 expression were subjected to qRT-PCR analysis.
RNA immunoprecipitation (RIP) assay
We applied Magna RIP kit (Millipore, Bradford, MA, USA), Ago2 antibody (Abcam, Cambridge, MA, USA) and IgG antibody (Abcam) to determine whether miR-637 interacted with circ_0033596 and GRB2 as per the guidebook. Briefly, the lysates of HUVEC at the layer confluent were prepared using RIP lysis buffer (Millipore) and then incubated with magnetic beads conjugated with the above antibodies. After 24 h, RNA was purified and analyzed by qRT-PCR.
Statistical analysis
All data from 3 independent duplicate tests were analyzed on GraphPad Prism and shown as means±standard deviations. Significant differences were compared with two-tailed Student’s t-tests, Wilcoxon-Mann-Whitney test or one-way analysis of variance (ANOVA). P < 0.05 indicated statistical significance.
Results
Ox-LDL-induced AS-like HUVEC injuries
To mimic AS-like HUVEC injuries, we simulated HUVECs using various concentrations of ox-LDL (0, 25, 50 and 75μg/mL). As presented in Fig. 1A, ox-LDL treatment concentration-dependently repressed HUVEC viability. 50μg/mL ox-LDL was chosen for the following study as the 50% cell viability under this condition. Subsequently, we found that 50μg/mL ox-LDL inhibited HUVEC proliferation and induced cell apoptosis, accompanied by an increase of Bax expression and a decrease of Bcl-2 expression (Fig. 1B–E). As shown in Fig. 1F and G, tube formation of HUVECs was also repressed after ox-LDL treatment. Moreover, the results showed that ox-LDL treatment increased the production of IL-6 and TNF-α (Fig. 1H). Further, ox-LDL stimulation led to decreased SOD activity and increased MDA level (Fig. 1I and J). Thus, these results suggested that ox-LDL could induce AS-like HUVEC injuries.
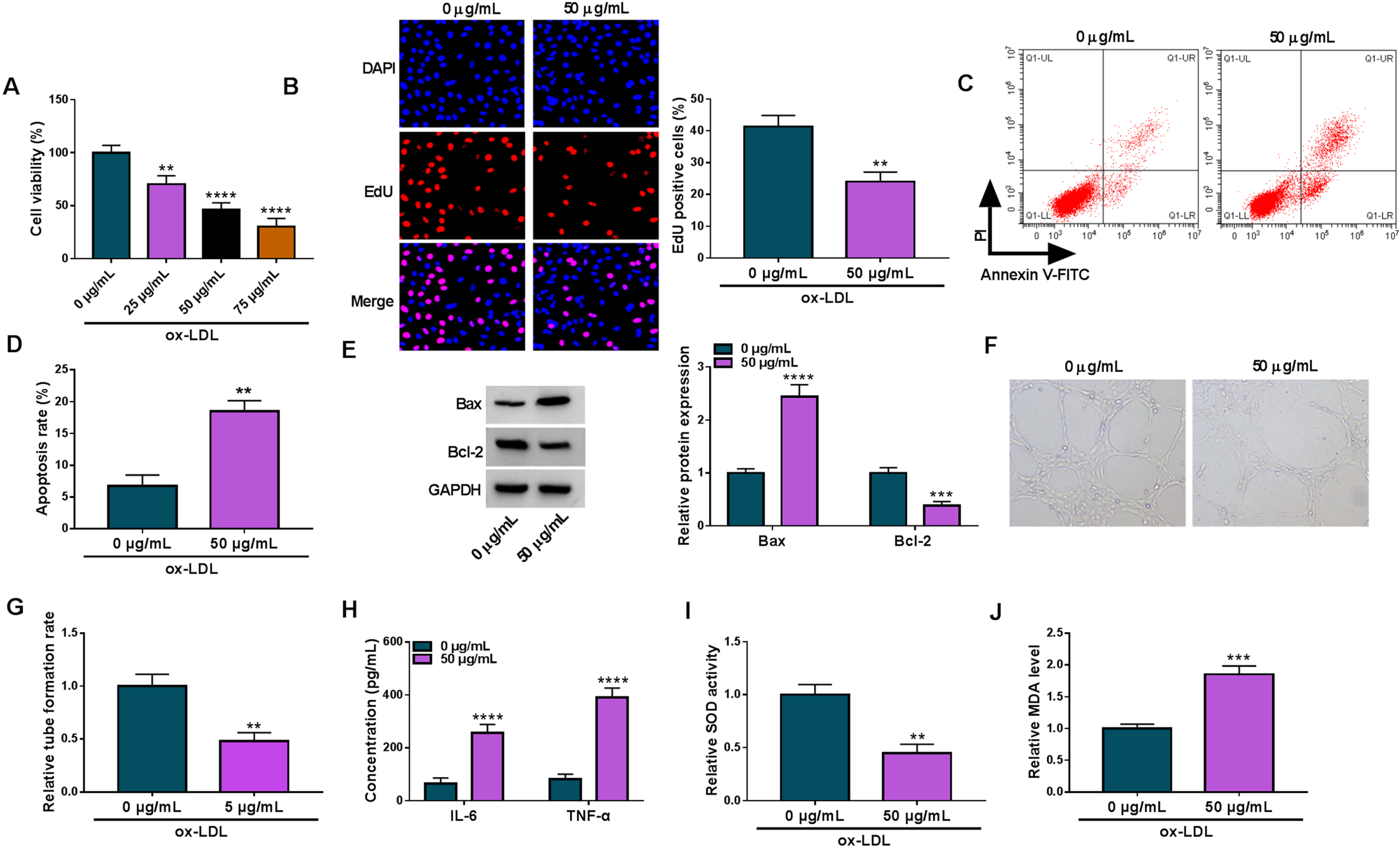
Ox-LDL-induced AS-like HUVEC injuries. (A) The effects of various concentrations of ox-LDL (0, 25, 50 and 75μg/mL) on HUVEC viability were assessed by CCK-8 assay. HUVECs were treated with ox-LDL (0 or 50μg/mL), and cell proliferation was analyzed by EdU assay (B), cell apoptosis by flow cytometry (C and D), the protein expression of Bax and Bcl-2 by Western blot (E), tube formation by tube formation assay (F and G), the production of IL-6 and TNF-α by ELISA (H), SOD activity by SOD activity assay kit (I) and MDA level by lipid peroxidation MDA assay kit (J). **P < 0.01, ***P < 0.001 and ****P < 0.0001.
Then, we analyzed circ_0033596 expression in the blood of AS patients. As shown in Fig. 2A, circ_0033596 expression was dramatically increased in the blood samples of AS patients when compared with the blood samples of healthy volunteers. Subsequently, the results showed the value of circ_0033596 was high in distinguishing AS from healthy controls with the AUC of 0.928 (Fig. 2B). We also analyzed the effects of ox-LDL treatment on circ_0033596 expression using qRT-PCR, and the results revealed that ox-LDL treatment concentration-dependently increased circ_0033596 expression (Fig. 2C). In addition, we confirmed the circular structure of circ_0033596 using RNase R, oligo(dT)18 primer and random primer. For instance, circ_0033596 was resistant to RNase R digestion when compared with liner GAPDH (Fig. 2D). Meanwhile, compared with using random primers, circ_0033596 expression was significantly decreased in HUVECs when using the oligo(dT)18 primer, while GAPDH was not (Fig. 2E). Further, the results showed that circ_0033596 was mainly expressed in the cytoplasm of HUVECs (Fig. 2F). Therefore, these results demonstrated that circ_0033596 might be involved in AS occurrence.
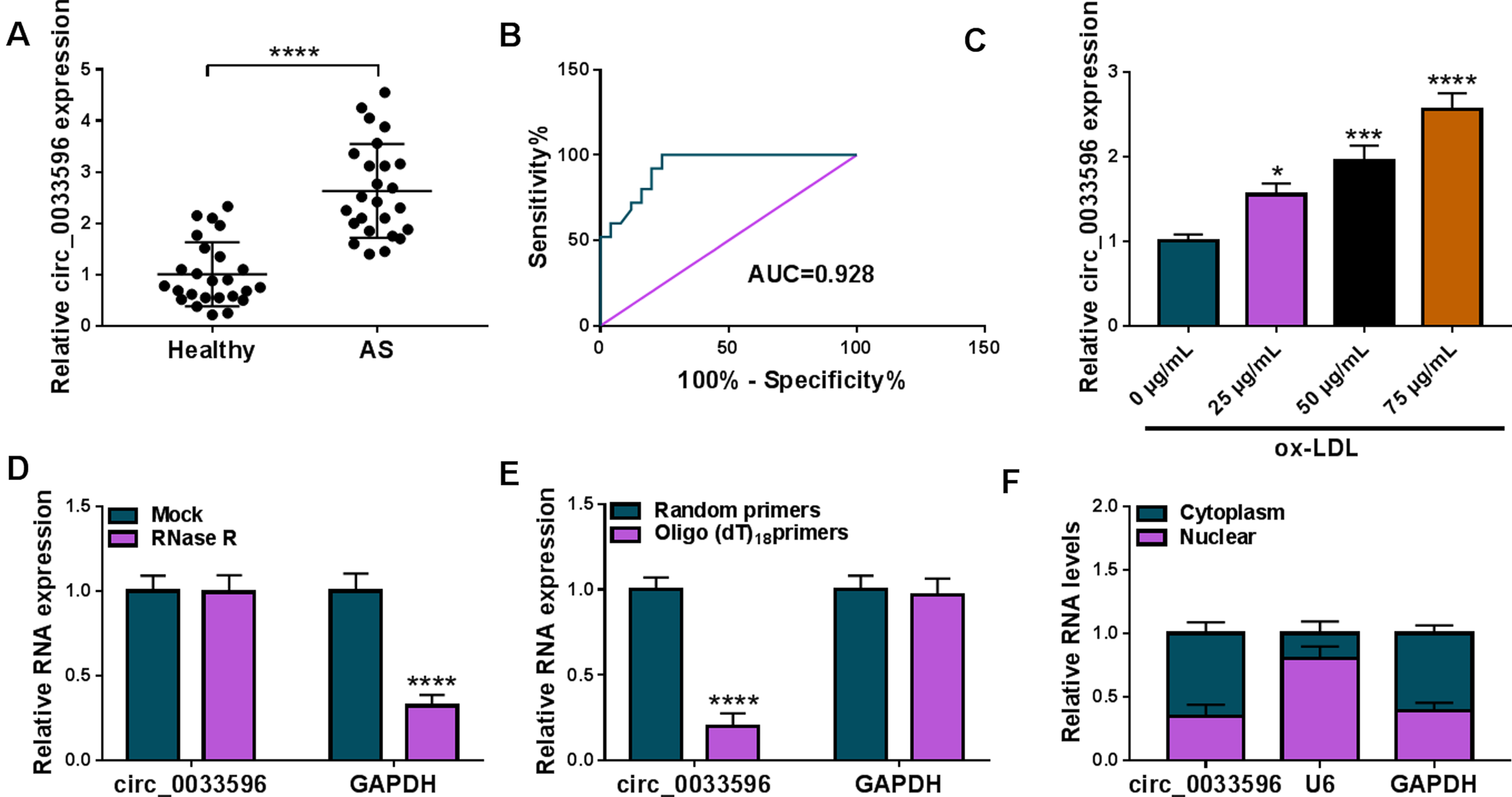
Circ_0033596 expression was increased in the blood of AS patients and ox-LDL-induced HUVECs. (A) Circ_0033596 expression was detected by qRT-PCR in the blood of AS patients and healthy volunteers. (B) ROC curve and areas under the curve were used to determine the diagnostic value of circ_0033596 in AS. (C) The effects of various concentrations of ox-LDL treatment on circ_0033596 expression were analyzed by qRT-PCR. (D and E) The circular structure of circ_0033596 was analyzed using RNase R, oligo(dT)18 primer and random primer. (F) Subcellular fractionation assay was used to assess the location of circ_0033596 in HUVECs. *P < 0.05, ***P < 0.001 and ****P < 0.0001.
We then silenced circ_0033596 expression in ox-LDL-stimulated HUVECs to determine the consequent effects on cell biological behaviors. The results first showed that ox-LDL treatment increased circ_0033596 expression, whereas the effect was relieved after transfection with si-circ_0033596 (Fig. 3A). Then, ox-LDL stimulation led to decreased HUVEC viability and proliferation; however, these effects were attenuated when circ_0033596 expression was reduced (Fig. 3B and C). As shown in Fig. 3D and E, the increased cell apoptosis and Bax protein expression and the decreased Bcl-2 expression induced by ox-LDL were restored by reducing circ_0033596 expression. Moreover, ox-LDL treatment repressed HUVEC tube formation and promoted IL-6 and TNF-α promotion, whereas these effects were restored after circ_0033596 depletion (Fig. 3F and G). Further, ox-LDL stimulation inhibited SOD activity and increased MDA level, but these effects were remitted when circ_0033596 expression was decreased (Fig. 3H and I). The above data manifested that circ_0033596 depletion ameliorated ox-LDL-induced HUVEC damage.

Circ_0033596 knockdown ameliorated ox-LDL-induced HUVEC damage. HUVECs were divided into control group (mock HUVECs), ox-LDL group, ox-LDL+si-NC group and ox-LDL+si-circ_0033596 group, and circ_0033596 expression was analyzed by qRT-PCR (A), cell viability by CCK-8 assay (B), cell proliferation by EdU assay (C), cell apoptosis by flow cytometry (D), the protein expression of Bax and Bcl-2 by Western blot (E), tube formation by tube formation assay (F), the production of IL-6 and TNF-α by ELISA (G), SOD activity by SOD activity assay kit (H) and MDA level by lipid peroxidation MDA assay kit (I). **P < 0.01, ***P < 0.001 and ****P < 0.0001.
Circular RNA interactome was used to predict the target miRNAs of circ_0033596. MiR-637, a candidate, was found to potentially bind to circ_0033596 (Fig. 4A). Subsequently, we conducted a series of experiments to validate the association between circ_0033596 and miR-637. The high efficiency of miR-637 overexpression in HUVECs was presented in Fig. 4B. As shown in Fig. 4C, miR-637 overexpression significantly repressed the luciferase activity of WT-circ_0033596 but not that of MUT-circ_0033596. Moreover, biotin-labeled miR-637 could dramatically enrich circ_0033596 when compared with biotin-labeled miR-NC (Fig. 4D). In addition, both miR-637 and circ_0033596 expression were higher in the co-precipitated RNAs induced by the Ago2 antibody than in the co-precipitated RNAs induced by the IgG antibody (Fig. 4E). The data from Fig. 4F and G showed that miR-637 expression was downregulated and negatively correlated with circ_0033596 expression in the blood of AS patients. Further, the results showed that ox-LDL treatment concentration-dependently decreased miR-637 expression in HUVECs (Fig. 4H). Collectively, these data demonstrated that circ_0033596 bound to miR-637 in HUVECs.

Circ_0033596 bound to miR-637 in HUVECs. (A) The schematic illustration showed the potential binding sites of circ_0033596 for miR-637. (B) The efficiency of miR-637 overexpression was analyzed by qRT-PCR in HUVECs. (C-E) Dual-luciferase reporter assay, RNA pull-down assay and RIP assay were conducted to identify the association of circ_0033596 and miR-637 in HUVECs. (F) MiR-637 expression was analyzed by qRT-PCR in the blood of AS patients and healthy volunteers by qRT-PCR. (G) Circ_0033596 expression was negatively correlated with miR-637 expression in the blood of AS patients. (H) The effects of various concentrations of ox-LDL on miR-637 expression were analyzed by qRT-PCR in HUVECs. **P < 0.01, ***P < 0.001 and ****P < 0.0001.
Given the association of circ_0033596 with miR-637 in HUVECs, we wondered whether miR-637 participated in the regulation of circ_0033596 on ox-LDL-induced HUVEC injury. Firstly, the results showed that circ_0033596 depletion increased miR-637 expression under ox-LDL treatment, whereas the effect was remitted after miR-637 expression was decreased (Fig. 5A). Subsequently, circ_0033596 knockdown promoted the viability and proliferation of ox-LDL-induced HUVECs, but these effects were rescued when miR-637 expression was decreased (Fig. 5B and C). The results of Fig. 5D and E showed that circ_0033596 absence inhibited HUVEC apoptosis, decreased Bax expression and increased Bcl-2 expression under ox-LDL treatment; however, these effects were relieved after transfection with miR-637 inhibitors. In addition, circ_0033596 silencing promoted HUVEC tube formation and repressed the promotion of IL-6 and TNF-α in the presence of ox-LDL, while these effects were remitted by reducing miR-637 expression (Fig. 5F and G). Further, the increased SOD activity and decreased MDA production induced by circ_0033596 depletion were relieved after transfection with miR-637 inhibitors in ox-LDL-induced HUVECs (Fig. 5H and I). Collectively, these data manifested that the circ_0033596/miR-637 pathway regulated ox-LDL-induced HUVEC injuries.
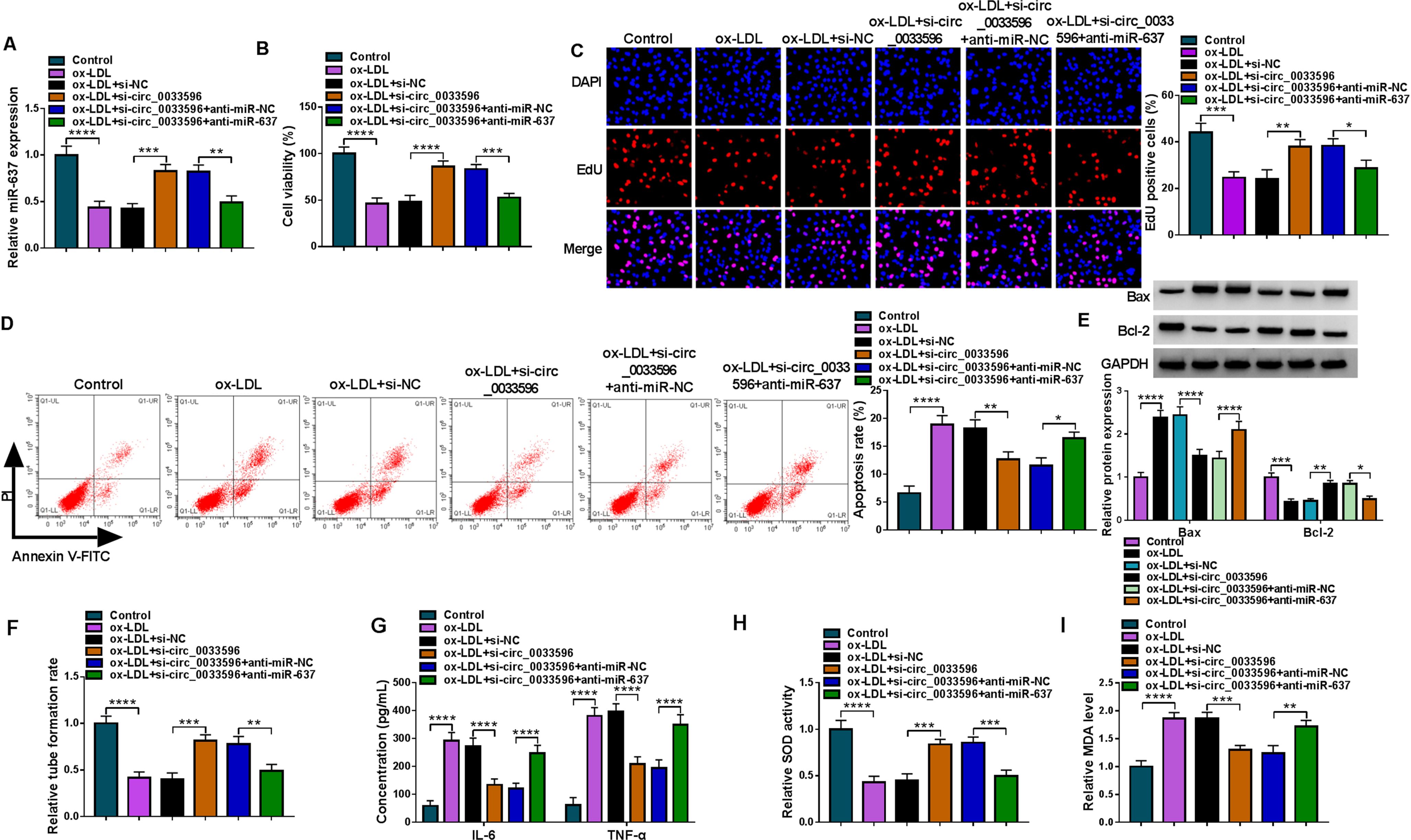
The circ_0033596/miR-637 pathway regulated ox-LDL-induced HUVEC injuries. HUVECs were divided into 6 groups, including control group, ox-LDL group, ox-LDL+si-NC group, ox-LDL+si-circ_0033596 group, ox-LDL+si-circ_0033596+anti-miR-NC group and ox-LDL+si-circ_0033596+anti-miR-637 group, and miR-637 expression was analyzed by qRT-PCR (A), cell viability by CCK-8 assay (B), cell proliferation by EdU assay (C), cell apoptosis by flow cytometry (D), the protein expression of Bax and Bcl-2 by Western blot (E), tube formation by tube formation assay (F), the production of IL-6 and TNF-α by ELISA (G), SOD activity by SOD activity assay kit (H) and MDA level by lipid peroxidation MDA assay kit (I). *P < 0.05, **P < 0.01, ***P < 0.001 and ****P < 0.0001.
We continued to search for the targeting genes of miR-637 using Targetscan online database. As shown in Fig. 6A, miR-637 contained the complementary sites of GRB2, suggesting the potential binding relationship of miR-637 for GRB2. Subsequently, dual-luciferase reporter assay revealed that miR-637 introduction repressed the luciferase activity of wild-type reporter plasmid of GRB2 3’UTR rather than that of mutant reporter plasmid of GRB2 3’UTR (Fig. 6B). Also, GRB2 expression was higher in the bio-miR-637 group than in the bio-miR-NC group (Fig. 6C). In addition, as revealed by the RIP assay, miR-637 and GRB2 expression were significantly enriched in the Ago2 antibody group when compared with the IgG antibody group (Fig. 6D). As expected, GRB2 expression was dramatically increased and negatively correlated with miR-637 in the blood samples of AS patients (Fig. 6E and F). Comparatively, ox-LDL treatment upregulated GRB2 expression in HUVECs in a concentration-dependent manner (Fig. 6G). Thus, these results showed that miR-637 targeted GRB2 in HUVECs.
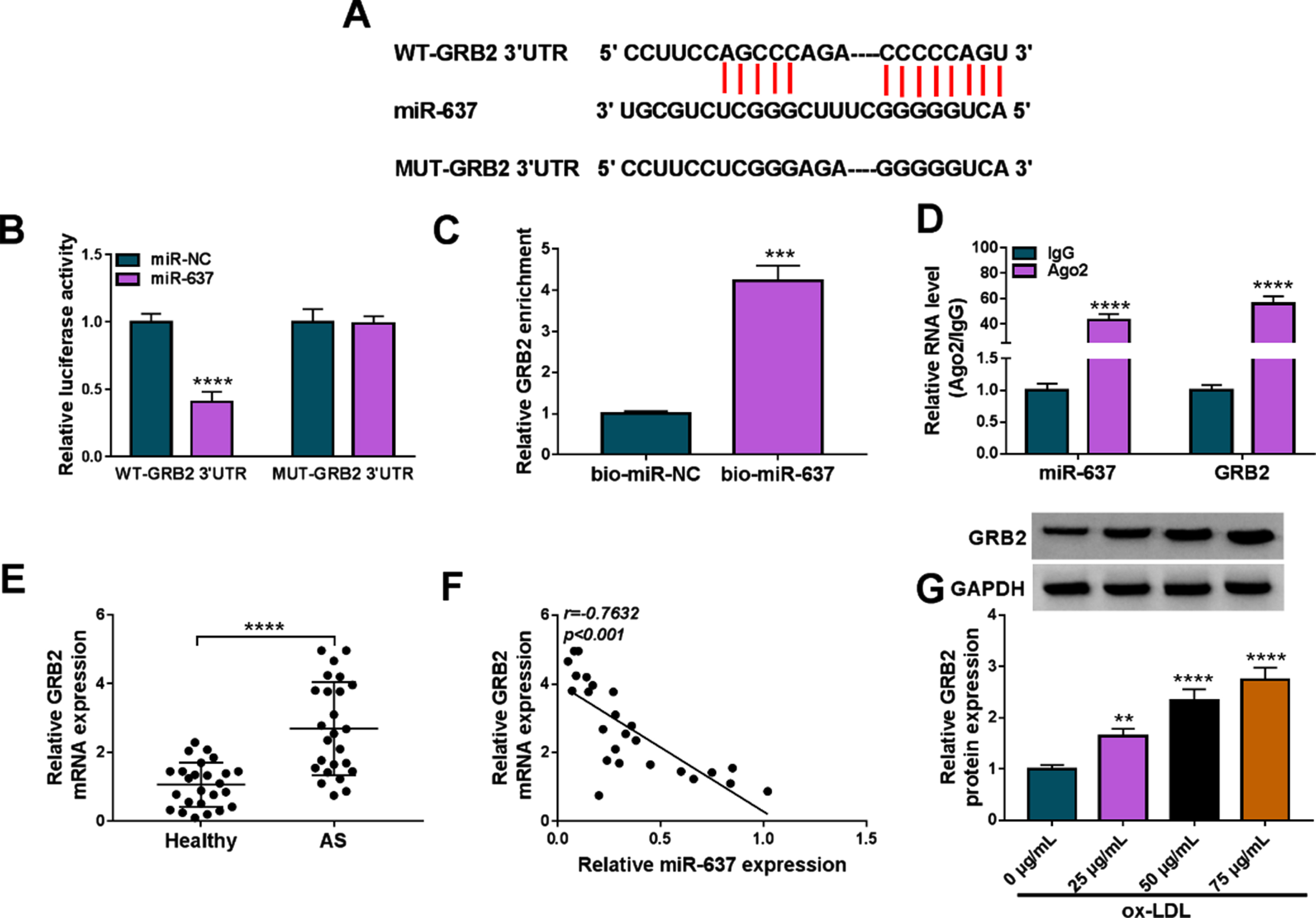
MiR-637 targeted GRB2 in HUVECs. (A) The schematic illustration showed the binding sites of miR-637 for GRB2. (B-D) Dual-luciferase reporter assay, RNA pull-down assay and RIP assay were conducted to identify the association of miR-637 and GRB2 in HUVECs. (E) GRB2 expression was analyzed by qRT-PCR in the blood of AS patients and healthy volunteers by qRT-PCR. (F) MiR-637 expression was negatively correlated with GRB2 expression in the blood of AS patients. (G) The effects of various concentrations of ox-LDL on GRB2 protein expression were analyzed by Western blot in HUVECs. **P < 0.01, ***P < 0.001 and ****P < 0.0001.
We then overexpressed miR-637 and GRB2 in ox-LDL-stimulated HUVECs to explore the consequent effects on cell proliferation, apoptosis, inflammation and oxidative stress. As presented in Fig. 7A, miR-637 overexpression repressed GRB2 protein expression in the cells, whereas the effect was relieved when GRB2 expression was increased. Subsequently, miR-637 introduction led to increased cell viability and proliferation, decreased cell apoptosis and dysregulated Bax and Bcl-2 expression under ox-LDL treatment; however, these effects were restored after transfection with GRB2 overexpression plasmid (Fig. 7B–E). In addition, miR-637 mimics promoted HUVEC tube formation and decreased IL-6 and TNF-α production, while these effects were relieved by GRB2 overexpression (Fig. 7F and G). Further, the increased SOD activity and decreased MDA level by increasing miR-637 expression were rescued after GRB2 expression was increased in ox-LDL-stimulated HUVECs (Fig. 7H and I). Therefore, the above data demonstrated that the miR-637/GRB2 axis regulated ox-LDL-caused HUVEC injuries.
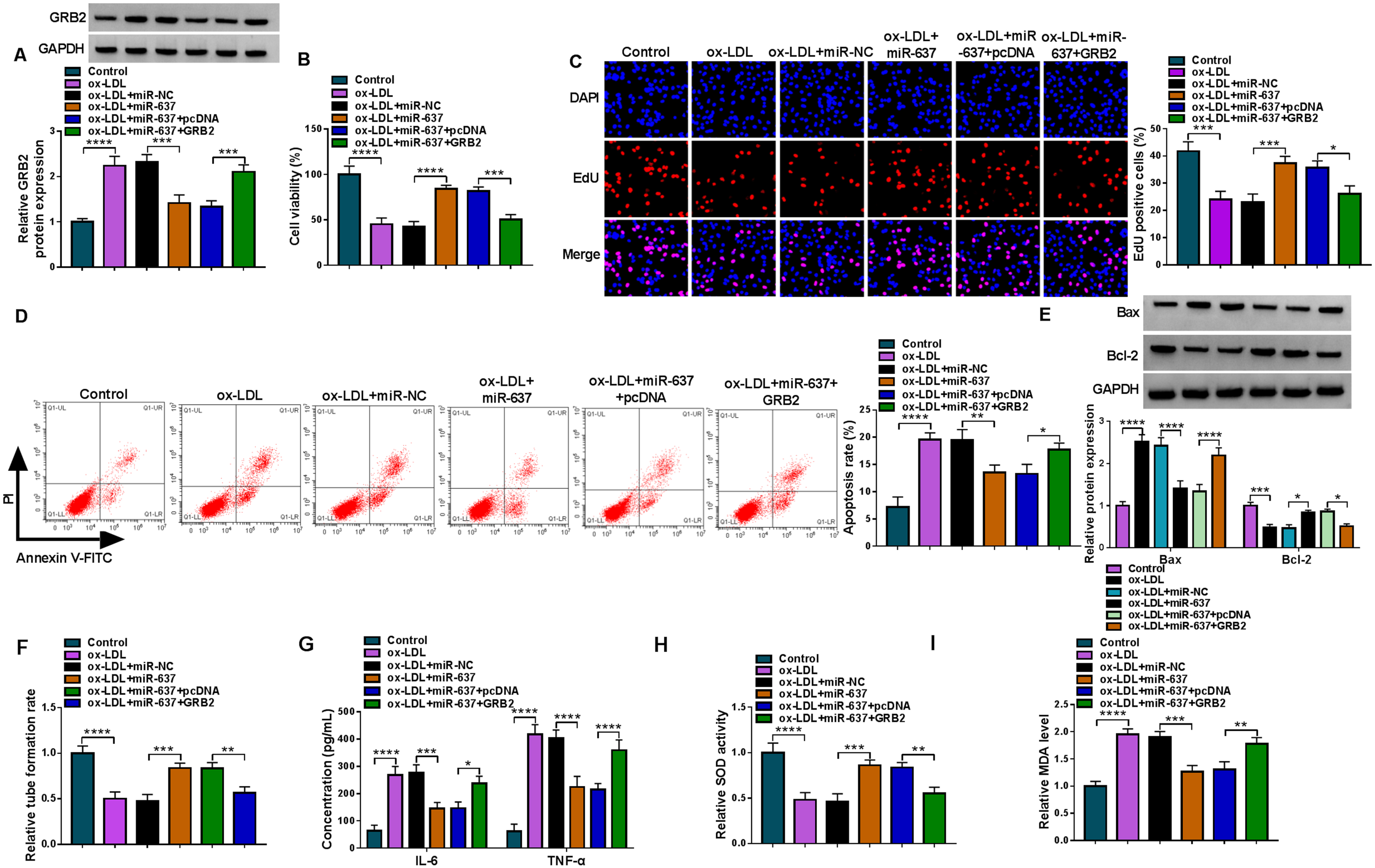
The miR-637/GRB2 axis regulated ox-LDL-caused HUVEC injuries. HUVECs were grouped into 6 groups, including control group, ox-LDL group, ox-LDL+miR-NC group, ox-LDL+miR-637 group, ox-LDL+miR-637+pcDNA and ox-LDL+miR-637+GRB2 group, and GRB2 protein expression was analyzed by Western blot (A), cell viability by CCK-8 assay (B), cell proliferation by EdU assay (C), cell apoptosis by flow cytometry (D), the protein expression of Bax and Bcl-2 by Western blot (E), tube formation by tube formation assay (F), the production of IL-6 and TNF-α by ELISA (G), SOD activity by SOD activity assay kit (H) and MDA level by lipid peroxidation MDA assay kit (I). *P < 0.05, **P < 0.01, ***P < 0.001 and ****P < 0.0001.
The study further assessed the association of GRB2 with circ_0033596 and miR-637 in ox-LDL-induced HUVECs. To this end, we silenced circ_0033596 and miR-637 expression in ox-LDL-induced HUVECs and then detected GRB2 expression by qRT-PCR and Western blot. As presented in Fig. 8A, circ_0033596 depletion reduced GRB2 mRNA expression in the cells, whereas the effect was relieved after transfection with miR-637 inhibitors. Meanwhile, the decreased GRB2 protein expression induced by circ_0033596 knockdown was attenuated after miR-637 downregulation in the cells (Fig. 8B). Thus, these data demonstrated that circ_0033596 mediated GRB2 expression by regulating miR-637 in ox-LDL-induced HUVECs.

Circ_0033596 mediated GRB2 expression by regulating miR-637 in ox-LDL-induced HUVECs. (A and B) The effects of circ_0033596 depletion and miR-637 silencing on GRB2 expression were determined by qRT-PCR and Western blot in ox-LDL-induced HUVECs. **P < 0.01, ***P < 0.001 and ****P < 0.0001.
AS contributes to the development of cardiovascular diseases, and its induction factors include hypertension, diabetes, hyperlipidemia as well as smoking [30, 31]. Vascular endothelium cells, essential components of the blood vessel, show the vasoprotective endothelial phenotype and its dysfunction leads to the earliest detectable change of atherosclerotic lesions [32, 33]. As revealed by both human and animal models, endothelial permeability after endothelial cell activation involves intimal lipid accumulation [34]. Herein, we explored the mechanism of circ_0033596 during AS using ox-LDL-induced HUVECs and found circ_0033596 depletion ameliorated AS-like cell injuries through the miR-637/GRB2 pathway.
Interacting genetic and epigenetic programs maintain homeostasis in the cardiovascular system, and the disequilibrium of which is responsible for AS occurrence [35, 36]. CircRNA, a noncoding RNA, is functional, as it regulates specific physiological processes. CircRNA is required for blood vessel development and thus is involved in cardiovascular diseases [13]. In addition, circRNA participates in the molecular mechanism of endothelial cell dysregulation during AS [30]. In our work, circ_0033596 had a diagnostic value in AS. We also found that circ_0033596 was upregulated in ox-LDL-stimulated cells. HUVEC proliferation inhibition and apoptosis promotion induced by ox-LDL were ameliorated after circ_0033596 depletion, which was consistent with the previous report [17]. We also demonstrated that the inhibitory effect of ox-LDL on HUVEC tube formation and the promoting effects on inflammation and oxidative stress involved circ_0033596 overexpression for the first time. Thus, our data suggested that circ_0033596 knockdown exhibited vasoprotective properties during AS.
MiRNA has received most of the attention for its regulatory roles in the pathophysiology of atherosclerosis [37]. Known as a noncoding RNA, miRNA can combine with circRNA to participate in many biological processes [38]. The present study identified circ_0033596 targeted miR-637. Previous studies have explained the diagnostic values of miR-637 in patients with AS [25] or carotid artery stenosis [39]. The work of Yang et al. explained that miR-637 depletion negatively regulated insulin-like growth factor-2 to promote VSMC proliferation and migration in AS [40]. Moreover, miR-637 could promote endothelial cell proliferation and migration after ox-LDL stimulation by the circUSP36/miR-637/Wnt family member 4 (WNT4) pathway, as revealed by an in vitro AS cell model [41]. In this work, miR-637 expression was reduced in the blood of AS patients and ox-LDL-treated HUVECs. Importantly, the reduced expression of circ_0033596 protected against ox-LDL-induced HUVEC damage through interaction with miR-637.
To analyze the detailed mechanism of miR-637 in ox-LDL-triggered HUVEC injury, we applied the Targetscan database to predict the target gene of miR-637. Subsequent assays confirmed that miR-637 targeted GRB2. GRB2, a cytoplasmic protein, contains a single SH2 domain and is associated with basic cellular functions [42]. As reported, GRB2 can sense growth factor receptor activation through its SH2 domain. The SH3 domain of GRB2 interacts with son-of-sevenless (SOS) to promote stable complex formation. GRB2 participates in cell proliferation, cell motility, epithelial morphogenesis and angiogenesis [43]. In addition, a recent study demonstrated that ox-LDL-induced foam cell formation involved the regulation of GRB2, as revealed using Grb2+/–mice [44]. Xi et al. proved that the generation of NADPH oxidase 4-derived ROS during AS involved GRB2 [45]. Previous work has revealed that GRB2 introduction exhibited inhibitory effects on miR-153-3p-mediated cell inflammation, apoptosis and oxidative stress after ox-LDL stimulation [46]. The above evidence suggested that GRB2 was important for AS development. The present study demonstrated that GRB2 combined with miR-637 to regulate ox-LDL-induced cell damage. Further, we found circ_0033596 induced GRB2 production by trapping miR-637. Therefore, GRB2 participated in the circ_0033596/miR-637 axis-mediated regulation on AS-like cell injuries.
In summary, the pathogenesis of AS involved circ_0033596 overexpression. In terms of mechanism, ectopic circ_0033596 expression induced GRB2 production by trapping miR-637 to induce vascular endothelium cell inflammation, apoptosis and oxidative stress during AS. Therefore, these novel findings might contribute to the investigation into therapeutic targets for AS. Nevertheless, in vivo experiments should be conducted to validate the regulatory mechanism that circ_0033596 combined with miR-637 and GRB2 to mediate AS development.
Footnotes
Acknowledgment
None.
Conflict of interest
The authors declare that they have no conflicts of interest.
Funding
None.
Authors’ contributions
Yanling Teng and Fei Ren conceived and designed the experiments; Yanan Wang, Hua Xu, Hejian Song performed the experiments; Yanling Teng and Yanan Wang analyzed and interpreted the data; Yanan Wang, Hua Xu, Hejian Song contributed reagents, materials, analysis tools or data; Yanling Teng and Fei Ren wrote the paper.
Availability of data and material
Not applicable.
Code availability
Not applicable.
Ethics approval and consent to participate
Written informed consents were obtained from all participants and this study was permitted by the Ethics Committee of the First People’s Hospital of Lianyungang, the First Affiliated Hospital of Kangda College of Nanjing Medical University.
Consent for publication
Not applicable.