
Abstract
OBJECTIVE:
Endothelial cells play a critical role in maintaining vascular function and kinetic homeostasis, but excessive accumulation of palmitic acid (PA) may lead to endoplasmic reticulum stress and trigger endothelial cell dysfunction. Baicalin (BCL), a natural plant extract, has received widespread attention for its biological activities in anti-inflammation and anti-oxidative stress. However, the mechanism of BCL on PA-induced endothelial cell dysfunction is unclear. Therefore, the aim of this study was to investigate whether BCL could inhibit PA-induced endoplasmic reticulum stress and thus attenuate endothelial cell dysfunction.
METHODS:
Human umbilical vein endothelial cells (HUVECs) were divided into Control, PA, PA + BCL-10 μM, PA + BCL-20 μM, and PA + BCL-50 μM groups. The PA group was treated with PA (200 μM), while the PA + BCL groups were co-treated with different concentrations of BCL (10 μM, 20 μM, 50 μM) for 24 hours. Cell viability was detected by MTT. Cell migration ability was determined by Transwell assay, apoptosis level by flow cytometry, and tube formation ability by tube formation assay. Finally, the levels of apoptosis-related proteins (Bax, Bcl-2, and cleaved caspase-3) and angiogenesis-related proteins (VEGFA and FGF2) were detected by western blot, MMP-9, as well as the protein levels of endoplasmic reticulum stress biomarkers (GRP78, CHOP, PERK, and ATF4).
RESULTS:
The results at the cellular level showed that cell viability, migration ability and tube formation ability of PA-induced HUVECs were significantly reduced, while apoptosis level was significantly increased. However, administration of different concentrations of BCL significantly enhanced PA-induced cell viability, migration ability and tube formation ability of HUVECs while inhibiting apoptosis. The results of protein levels showed that the protein levels of Bax and cleaved caspase-3 were observably up-regulated in the cells of the PA group, while the protein level of Bcl-2 was significantly down-regulated; compared with the PA group, the protein levels of Bax and cleaved caspase-3 were much lower and the Bcl-2 protein level was much higher in the PA + BCL group. Additionally, the protein levels of VEGFA, FGF2 and MMP-9 were raised and those of GRP78, CHOP, PERK and ATF4 were lowered in the PA + BCL group of cells in a concentration-dependent manner.
CONCLUSION:
BCL significantly attenuates PA-induced endothelial cell dysfunction by inhibiting endoplasmic reticulum stress.
Introduction
In recent years, the incidence of cardiovascular diseases has steadily climbed and grown to be a major global public health concern. Endothelial cells are the main cell type that constitutes the inner lining of the blood vessels, and they are crucial for maintaining the normal function and structure of blood vessels [1, 2]. Endothelial cells play a key role in regulating vascular tone, anti-inflammation, anticoagulation, and anti-platelet aggregation [3]. However, some exogenous factors, such as hyperlipidemia, hyperglycemia, and inflammation, can lead to endothelial cell dysfunction, which in turn induces the onset and development of cardiovascular diseases [4–8]. Among these factors, excessive accumulation of fatty acids, especially palmitic acid (PA), is widely recognized as one of the important factors responsible for endothelial cell dysfunction [9, 10]. Gu and Luo et al. demonstrated that PA treatment induced endoplasmic reticulum stress (ERS), dysfunction of the endoplasmic reticulum, apoptosis, cardiovascular diastole, inflammation of the vessel wall, and an increased risk of diseases such as atherosclerosis [11–13]. Therefore, elucidating the stress response mechanism of endothelial cells responding to excessive lipid load and searching for effective interventions are important for the prevention and treatment of cardiovascular diseases.
Baicalin (BCL) is a natural plant compound extracted from Scutellaria baicalensis, which is widely discussed in traditional Chinese medicine research [14]. It has been shown that BCL possesses a variety of biological activities including antioxidant, anti-inflammatory, and anti-apoptotic activities [15, 16]. Xu et al. pointed out [17] that BCL can regulate macrophage polarization, reduce cellular response pressure, and attenuate endothelial cell injury and myocardial ischemia/reperfusion injury by inhibiting the JAK/STAT pathway. In the report of Feng et al. [18], BCL regulated TLR4/Iκ Bα/NFκB signaling pathway to reduce adriamycin-associated cardiotoxicity and inflammatory response. Collectively, BCL has an important role in cardiovascular disease, but its inhibition of ERS in endothelial cells as well as its effect on endothelial cell function have not been explored in depth.
The aim of this study was to systematically investigate whether BCL can inhibit PA-induced ERS and its potential protective effect on endothelial cell function through experiments. Human umbilical vein endothelial cells (HUVECs) were selected as the study subjects, and an ERS model was established by treating the cells with PA. The effects of BCL on cell viability, migration ability, apoptosis level, and tube formation ability were analyzed. In addition, the levels of key proteins were detected by western blot in order to delve into the regulatory mechanism of BCL on endothelial cell function. By comprehensively investigating the role of BCL in ERS of endothelial cells, this study aimed to provide a scientific basis for the development of new therapeutic strategies for cardiovascular diseases and experimental support for the application of BCL in the cardiovascular field.
Materials and methods
Cell culture and grouping
HUVECs were purchased from ATCC (No. AC337632). The cells were cultured in a Dulbecco’s Modified Eagle Medium (DMEM, GIBCO, XB01, USA) + 10% fetal bovine serum (FBS, GIBCO, 10270-106, USA) + 1% antibiotics (100 U/ml penicillin and 100 μg/ml streptomycin, GIBCO, 15240062, USA), and they were placed in an incubator with 5% CO2 at 37°C to ensure the normal growth of the cells.
Cell culture and group treatment
The cells were divided into the following five groups for the experiments: the Control group, HUVECs without any special treatment; the PA group, HUVECs treated with PA (200 μM) for 24 h; the PA + BCL-10 μM group, HUVECs treated with a combination of PA and BCL (10 μM) for 24 h; the PA + BCL-20 μM group, HUVECs treated with a combination of PA and BCL (20 μM) for 24 h; PA + BCL-50 μM group, HUVECs treated with a combination of PA and BCL (50 μM) for 24 h.
MTT assay for cell viability
MTT assay was used to assess the endothelial cell viability under different treatment conditions [19]. Firstly, the treated HUVECs from each group were inoculated into 96-well plates containing about 1×104 cells per well, and the cells were cultured for 24, 48, and 72 h, respectively. To each well, 10 μL of MTT solution (5 mg/mL) was added and the cells were incubated in an incubator at 37°C for 4 h. Then the medium was removed and 150 μL of DMSO was added to dissolve the formaldehyde crystals. Next, the plate was gently shaken to ensure complete dissolution. Lastly, the absorbance values were measured using a microplate reader (Thermo Fisher, USA) with the wavelength set at 570 nm.
Flow cytometry for apoptosis level
Apoptosis was detected by flow cytometry [20]. Specifically, the cell samples were collected from each group and washed twice with pre-cooled PBS to remove residual medium. Then they were added and mixed well with 500 μL of binding buffer, and centrifuged at 1000 rpm for 5 min. Upon washing the cells, 5 μL of Annexin V-FITC and 5 μL of PI (Elabscience, E-CK-A211, USA) were added. After mixing them gently, the cells were incubated for 15 min at room temperature away from light, and then centrifuged twice before washing. Finally, 100 μL of binding buffer were supplemented and mixed gently again. Using a flow cytometer (BECKMAN, CytoFLEX, USA), the apoptosis level of each group of cells was detected by Flowjo software.
Transwell assay for cell migration ability
Transwell assay was used to detect the migratory capacity of the cells [21]. A suspension of cells to be tested was collected from each group and 5×104 cells were added to each well of the culture plate in the upper chamber. Later, the upper chamber was inserted into the lower chamber and then they were isolated by means of a porous membrane (SigmaAldrich., AXYPCR16C, USA 8 μm) to prevent direct cell contact. Cell culture solution was added to the upper chamber, and the culture plate containing cells was placed in a thermostatic incubator. Following that, the cells were incubated in the incubator at 37°C and 5% CO2 for 48 hours to allow them to migrate from the upper chamber through the porous membrane into the lower chamber. At the end of the incubation time, the culture solution in the upper chamber was poured out and the cells in each Transwell were washed with cold PBS to stop cell migration. Next, the washed cells were fixed with 4% paraformaldehyde, filled in each well and placed at room temperature for 15 minutes. After the fixation, the fixative was carefully removed and each well was washed with cold PBS again. Subsequently, the migrated cells were stained with 0.5% crystal violet, and the number of the migrated cells was observed counted under a microscope (Leica, IVESTA3, Germany).
Tube formation assay
A 96-well plate was coated with 50 μl of matrix gel (3432-005-01, R& D systems, USA), which was polymerized at 37°C for 1 h. HUVECs were grown on the plate at a density of 1.5×104 cells per well, and vascular network formation was visible for 12 h of incubation [22]. Images were subsequently taken at 10× magnification from three random fields of view per well under a microscope (Leica, IVESTA3, Germany). The number of junctions per well and total branch length were analyzed using ImageJ software.
Western bolt for protein expression level
The cells in each group were washed on ice, and then added with RAPI protein extraction buffer (Beyotime, Shanghai, China) for lysis. The cell debris was removed by a 10-min centrifugation at 1500 rpm, the supernatant was collected, and the protein concentration was determined by a BCA kit (Beyotime, Shanghai, China). Subsequently, the protein samples were subjected to a SDS-PAGE electrophoresis (Beyotime, Shanghai, China). The resulting proteins were transferred onto a PVDF membrane (Thermo Fisher, USA). Later, the membrane was blocked with 5% skim milk powder at room temperature to prevent non-specific binding. Then it was incubated overnight at 4°C with specific primary antibodies against Bax (abcam, No. ab32503, 1 : 1000), Bcl-2 (abcam, No. ab141523, 1 : 2000), cleaved caspase-3 (abcam, No. ab32042, 1 : 1500), VEGFA (abcam (abcam, No. ab52917, 1 : 2000), FGF2 (abcam, No. ab208687, 1 : 2000), MMP-9 (abcam, No. ab141579, 1 : 1000), GRP78 (abcam, No. ab21685, 1 : 1000), CHOP (abcam, No. ab11419, 1 : 1500), PERK (abcam, No. ab229912, 1 : 2000), ATF4 (abcam, No. ab270980, 1 : 2000), and β-actin (abcam, No. ab8226, 1 : 5000). Afterwards, the unbound primary antibodies were removed by washing the membrane with PBS-Tween. HRP-labeled Anti-Rabbit and Anti-Mouse secondary antibodies (abcam, No. ab8226 and ab8226, 1 : 5000) were added for further incubation. Again, PBS-Tween was used to remove the unbound secondary antibodies. Protein bands were visualized using Enhanced Chemiluminescence Substrate (Thermo Fisher, USA) and acquired by Chemi-Image System (Tanon, 5200CE, USA).
Statistical analysis
Statistical analysis was carried out using SPSS 26.0 software. For the measurement data that conformed to normal distribution, they were expressed as the mean±standard deviation, while for the counting data, they were expressed as percentages (%). χ2 test was used to compare two groups, and one-way ANOVA to compare multiple groups. p < 0.05 means the difference is statistically significant.
Results
Baicalin increases viability and migration capacity of palmitic acid-induced endothelial cells
The results of MTT experiments showed significantly lower endothelial cell viability in the PA group relative to the Control group (p < 0.01). Compared with the PA group, the viability of cells in the PA + BCL-10 μM, PA + BCL-20 μM and PA + BCL-50 μM groups displayed a significant increase (p < 0.01), and this increase was time- and concentration-dependent. This indicated that BCL had a significant protective effect against PA-induced endothelial cell injury (Fig. 1A). In addition, the results of the Transwell assay revealed a significant decrease in the migration ability of endothelial cells in the PA group relative to the Control group (p < 0.01). Compared with the PA group, the migration ability of cells in the PA + BCL-10 μM group, PA + BCL-20 μM group and PA + BCL-50 μM group was significantly elevated, and this elevation showed a concentration-dependent pattern (p < 0.01). Therefore, BCL could also significantly enhance the migration ability of the PA-induced endothelial cells (Fig. 1B).

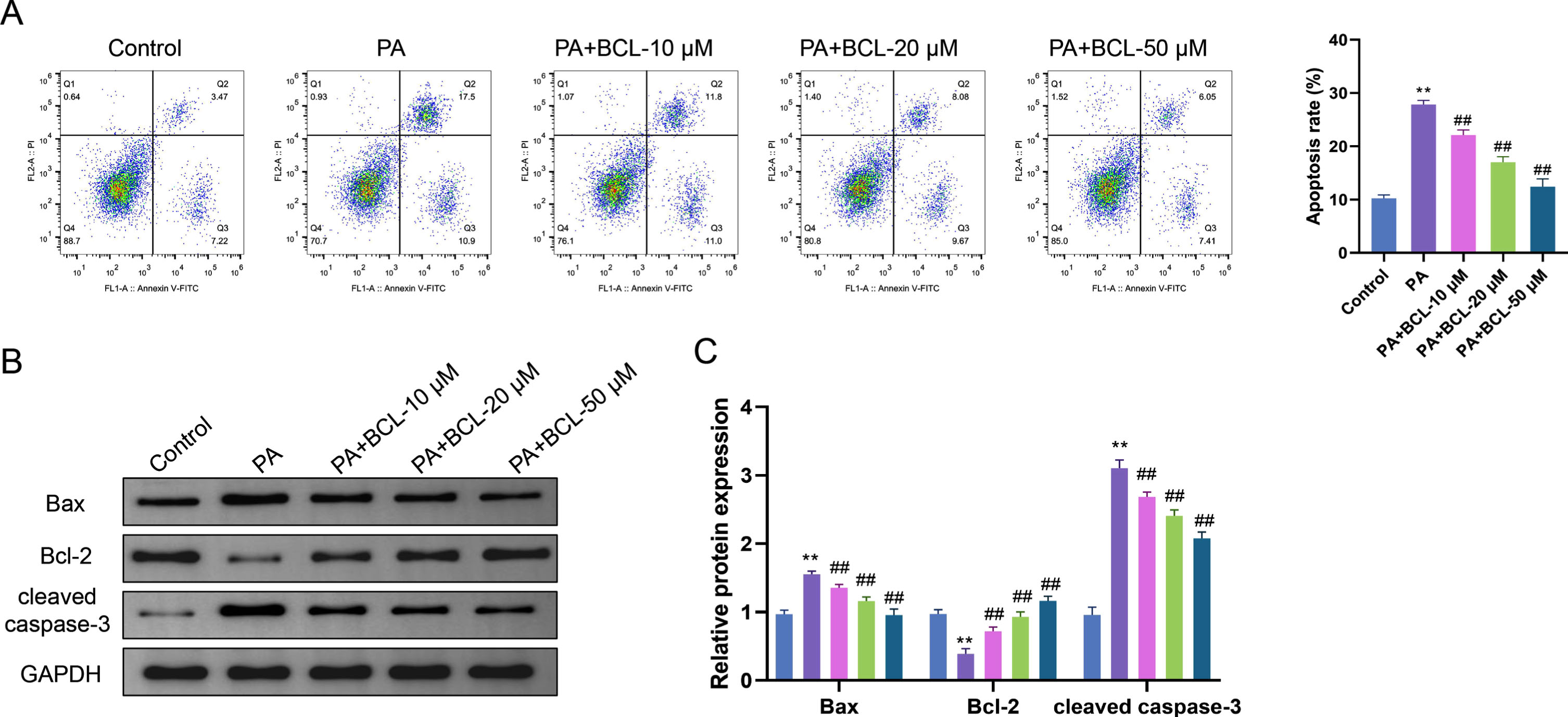
Flow cytometry results showed much higher apoptosis levels of endothelial cells in the PA group compared with the Control group (p < 0.05). The apoptosis levels of cells in the PA + BCL-10 μM group, PA + BCL-20 μM group and PA + BCL-50 μM group were significantly reduced in a concentration-dependent manner compared with the PA group (p < 0.01) (Fig. 2A). The results of protein level analysis showed that the protein levels of Bax and cleaved caspase-3 were considerably increased in endothelial cells in the PA group compared with the Control group, while the protein level of Bcl-2 was decreased (p < 0.01). This suggests that PA induced apoptosis in endothelial cells and contributed to changes in the expression levels of apoptosis-related proteins. Compared with the PA group, the protein levels of Bax and cleaved caspase-3 were obviously reduced in endothelial cells in the PA + BCL-10 μM group, PA + BCL-20 μM group, and PA + BCL-50 μM group, whereas the protein level of Bcl-2 was significantly elevated (p < 0.01), and these changes were concentration-dependent. It further confirms that BCL exerts a protective effect against endothelial cell apoptosis by regulating the protein expression of Bax, cleaved caspase-3, and Bcl-2 (Fig. 2B, C).
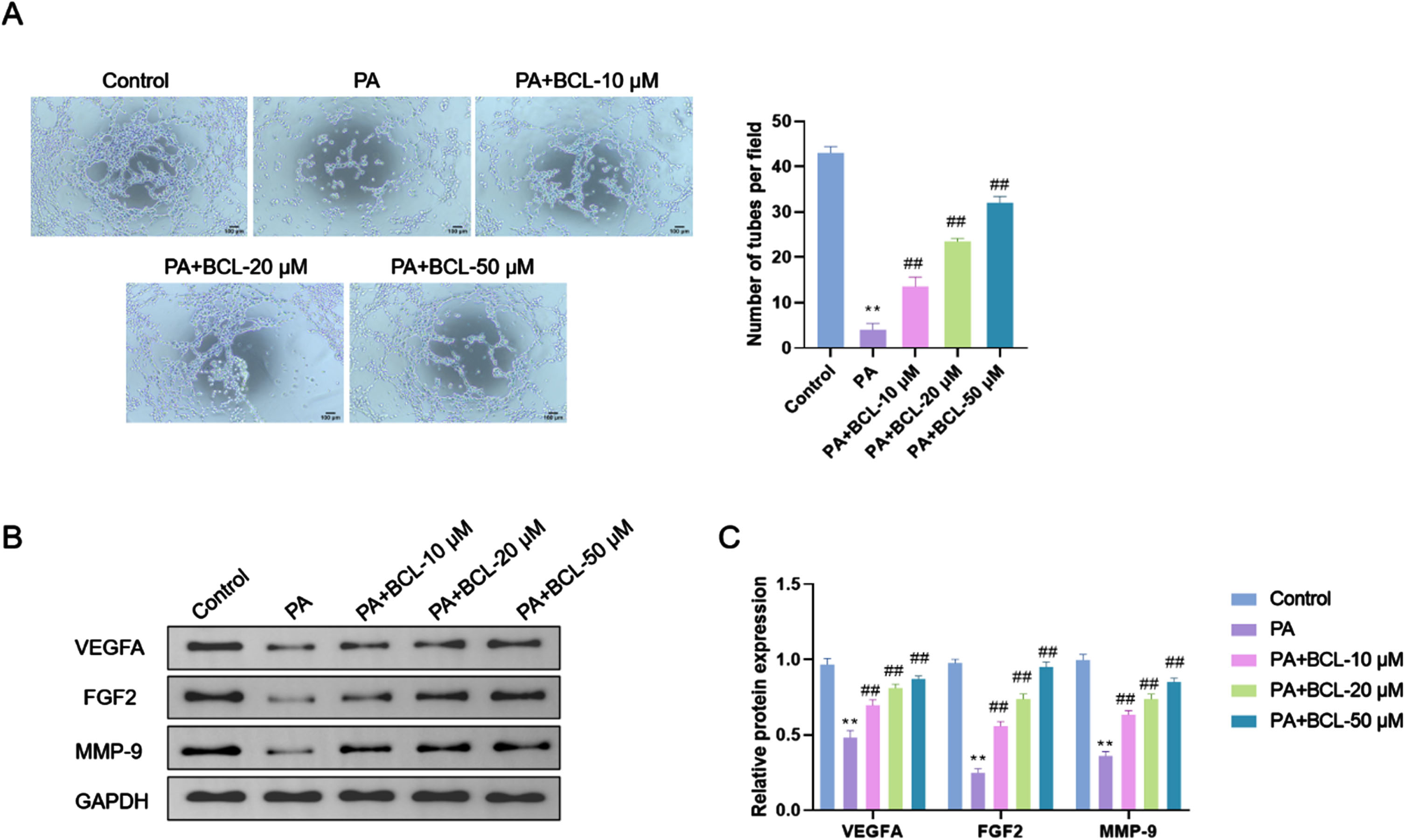
The results of the tube formation assay showed a significant reduction in the tube formation ability of endothelial cells in the PA group relative to the Control group (p < 0.01). The tube formation capacity of cells in the PA + BCL-10 μM, PA + BCL-20 μM, and PA + BCL-50 μM groups was significantly enhanced compared with that in the PA group, and this enhancement exhibited a concentration dependence (p < 0.01) (Fig. 3A). Protein level analysis results revealed much lower protein levels of VEGFA, FGF2, and MMP-9 in endothelial cells in the PA group than the Control group (p < 0.01). This indicated that PA treatment resulted in an obvious down-regulation of angiogenesis- and cell migration-related proteins. Compared with the PA group, the protein levels of VEGFA, FGF2, and MMP-9 were significantly elevated in endothelial cells in the PA + BCL-10 μM group, PA + BCL-20 μM group, and PA + BCL-50 μM group, and this elevation showed concentration-dependence (p < 0.01). BCL may promote angiogenesis and cell migration of endothelial cells(Fig. 3B, C).
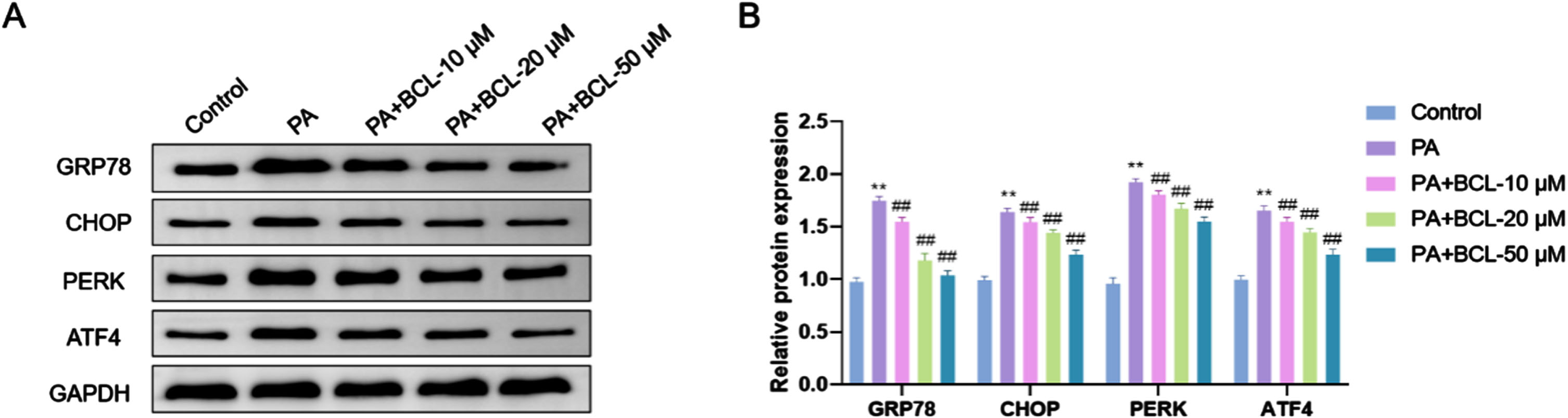
The protein levels of GRP78, CHOP, PERK, and ATF4 in endothelial cells were much higher in the PA group compared to the Control group (p < 0.01). This suggests that PA induced ERS response in endothelial cells, which led to a significant up-regulation of ERS-related proteins. Compared with the PA group, the protein levels of GRP78, CHOP, PERK, and ATF4 were significantly lowered in endothelial cells in the PA + BCL-10 μM group, the PA + BCL-20 μM group, and the PA + BCL-50 μM group, and this reduction was concentration-dependent (p < 0.01). Accordingly, BCL may alleviate endothelial cellular stress via regulating ERS pathway (Fig. 4A, B).
BCL is a flavonoid with rich medicinal value and multiple biological activities [23–25]. Ren et al. [26] uncovered that BCL exerted a protective effect in a variety of diseases such as diabetic nephropathy by inhibiting inflammation, oxidative stress and rapoptosis through the SphK1/S1P/NF-κB signaling pathway. The unique molecular structure of BCL makes it a potent antioxidant that aids in scavenging free radicals in the body and reducing cellular damage caused by oxidative stress. BCL has also been shown to have various pharmacological effects such as anti-inflammation, antibacteria, and antitumor [27]. In this study, we found that the expression of apoptotic proteins was significantly reduced after BCL treatment, suggesting a progressive decrease in the level of apoptosis. The reduction may imply the regulating effect of BCL on intracellular metabolic homeostasis and its improvement effect on the cellular adaptation to the internal and external environments. Furthermore, the concentration-dependent effects of BCL provide a direction for further research, which could help identify the optimal therapeutic concentration range of BCL. This not only contributes to a deeper comprehension of the regulatory mechanism of BCL on ERS, but also provides a scientific basis for its potential application in the treatment of endothelial cell function-related diseases.
ERS has been reported as a protective mechanism for cells to cope with external stresses, but the excessive stress may lead to cellular dysfunction [28–30]. For example, Nasoni et al. showed [31] that oxidized sterol-induced endothelial dysfunction was exacerbated when nitrosative stress attacked the endoplasmic reticulum. It is evident that the effects of ERS on cellular function are dual. PA, as a lipid component ubiquitously present in living organisms, is a member of the fatty acid family and is commonly found in a wide variety of foods, especially animal fats and vegetable oils. Excessive PA intake is closely associated with the development of ERS [32]. The ERS will triggers major dysfunctions, including abnormal protein folding, repair, and transport disorders, and dysfunctions of lipid synthesis and metabolism, all of which have non-negligible effects on cells [32, 33]. Vascular endothelial cells are considered the primary target of fatty acids which may induce endothelial dysfunction [34, 35]. Multiple reports indicate that in vitro, moderate to high concentrations of fatty acids (0.5–0.7 mM and higher) induce endothelial dysfunction and compromise endothelial cell viability via oxidative stress, inflammation, and apoptosis [36–38].
When there is an ERS, PA may induce a series of negative effects on endothelial cells [36]. Zeng et al. [39] observed that oleic acid could inhibit PA-induced ERS, and reduce the expression levels of ERS marker GRP78 and CHOP. Chu demonstrated [40] that excessive intake of PA may lead to the ERS, affecting the normal activities of endothelial cells, such as cell proliferation, migration, apoptosis, angiogenesis and immunoregulation. In light of this, it is of great theoretical and clinical significance to investigate the regulatory mechanisms of ERS in endothelial cells and the potential protective agents. In the present study, we discovered that BCL lowered the level of apoptosis in endothelial cells by modulating the core proteins of the ERS pathway, including Bax, Bcl-2 and cleaved caspase-3. In addition, BCL plays an active role in endothelial function by regulating proteins associated with cell migration and angiogenic proteins, such as VEGFA, FGF2 and MMP-9. In terms of potential molecular mechanisms, BCL significantly reduced the expression levels of key proteins in the ERS pathway, like GRP78, CHOP, PERK, and ATF4, thereby attenuating the ERS in endothelial cells. These results suggest that PA can induce toxic effects due to cellular lipid loading, which are consistent with previous findings of Zeng et al. that PA can trigger apoptosis [39]. Moreover, we found that BCL can attenuate this cytotoxic effect. It was hypothesized that BCL may decrease the ERS to inhibit apoptosis, promote angiogenesis and cell migration, improve the protective capacity of cells responding to various external and internal stresses, and finally maintain intracellular homeostasis.
There are certain limitations to this study, despite the fact that it has investigated in great detail how BCL regulates endothelial cell activity under ERS. First, the molecular mechanism of BCL was not elaborated in this study. Only the ERS pathway proteins that might be involved were mentioned, and a more specific in-depth analysis at the molecular level was absent. Second, only one endothelial cell line was used in our experiments, and we neglected to take into account the differential response of endothelial cells of different sources and types to BCL. In addition, the metabolic kinetics and pharmacokinetics of BCL in vivo were not systematically examined in this study, making it impossible to gain a comprehensive understanding of its bioavailability. These limitations need to be explored more concretely and comprehensively in future studies to further establish the potential application value of BCL in the treatment of related diseases.
Conclusion
In conclusion, the present study revealed the multifaceted regulatory effects of BCL on endothelial cell function under ERS, and initially outlined its potential mechanism of action at the molecular level, which provides a strong theoretical support for further research on the application of BCL in the treatment of related diseases.
Ethics approval and consent to participate
Not applicable.
Funding
This study did not receive any funding in any form.
Conflict of interest
The authors declare that they have no competing interests.
Data availability
The data used to support the findings of this study are available from the corresponding author upon request.
Author contributions
JC conceptualized and designed the study, drafted the initial manuscript. FC, CL and SY collected the data and carried out the initial analyses. JC critically reviewed the manuscript for important intellectual content. All authors approved the final manuscript as submitted and agree to be accountable for all aspects of the work.