
Abstract
BACKGROUND AND AIMS:
Although HCV is one of the major health problems worldwide with the highest prevalence of genotype 4a in Egypt, it is poorly understood because of the limitations of having a robust in vitro model that allows the investigation and understanding of viral pathogenesis and life cycle. Genomic replicons for HCV are widely used and proved to have strong replication efficiency in cell culture, however, they are not able to produce infectious particles to enable the investigation of the whole viral life cycle and they mostly represent few sub-genomic classes for HCV. Hence, Genotype specific replication system is necessary to address specific sub-genomic phenotypes related to Hepatitis C pathogenicity.
METHODS:
In this study we attempt to develop a sustainable co-culture model, which potentially provides essential route of infection for HCV by using HCV-positive sera from infected patients. In this novel in vitro model, we tested the viral replication in co-cultured Huh 7.5 and HepG2 cells in order to sustain full viral replication cycle. We used high viral load serum of HCV-infected patients (10
RESULTS AND CONCLUSIONS
: Viral replication capacity was increased 3–5 folds in the coculture condition compared to the individual cell lines, which indicates an improvement to viral infectivity in vitro.
SIGNIFICANCE STATEMENT
: This novel coculture system represents a new in vitro model that will help study the underlying mechanisms of HCV pathogenicity.
Abbreviations
HCV: Hepatitis C virus; DMEM: Dulbecco’s modified eagle medium; PCR: Polymerase Chain Reaction; HCC: Hepatocellular carcinoma; HuH 7: Human Hepatoma cells, HepG2: Hepatoma G2 cells.
Introduction
According to the recent world health organization report, 70 million people around the world are infected with Hepatitis C Virus (HCV) in 2015; accounting for 1% of the world’s population. European and Eastern Mediterranean regions are the most affected areas [11]. Egypt has the highest prevalence of HCV worldwide with approximately 15% of the population infected with HCV. In Egypt, the most predominant genotype is genotype 4a which is responsible for almost 90% of chronic hepatitis, and hepatocellular carcinoma [20].
The scarcity of having a robust infectious in vitro model for studying HCV replication has hindered scientific research efforts to understand the pathogenesis and life cycle of the virus, many groups attempted to establish a cell culture system to support viral replication to understand the relationships between the virus and its natural host. Many of these attempts were successful in having a cell culture model, however, lacking the virions production was hampering the sustainability of the HCV research in vitro [2, 12, 15, 16, 20, 27].
Recently, a significant HCV culture model was achieved by the subgenomic replicon which enabled consistent HCV replication in vitro. In 2005 Wakita et al. were able to clone HCV genotype 2a JFH1 which then was transfected into Huh-7 cell line, this recorded the first attempt to enhance virions production and replication [18]. Zhong et al. also was able to establish a strong system using infectious virus particles in Huh-7 cell line [12]. However, there are several limitations for the subgenomic replicon system, one is the utilization of the rare clone of HCV genotype 2a JFH1 [18, 25], another is the use of cloned HCV genotype 1a H77-S which was identified to have some adaptive mutations [2]. Other groups have studied HCV replication using positive serum to infect hepatocytes in vitro, using infected serum provides the cocktail which contains the essential factors serving as a nourishing environment for HCV virions biology, dynamics and life cycle [6, 7, 8, 9, 10, 16, 17, 18, 19, 20, 23].
HCV replicon system paved the way to identify potential permissive hepatoma cell lines. the discovery of Huh-7 clones (Huh-7, Huh-7.5, and Huh-7.5.1) introduced a new era in HCV research. these cell lines supported HCV replication, more specifically HuH 7.5 which showed high success in cell-based infection systems. Another hepatocyte cells, the HepG2 cells are derived from human hepatoblastoma, these cells are known to express liver-specific metabolites, and they have the ability to form polarized cell membranes in culture [3, 5, 22]. Naiive HepG2 cells were found to lack CD81; however, these cells observed increasing HCV infection if incorporated with CD81, it is worth highlighting that HepG2 cells are around 700 folds less than Huh-7.5 infective in vitro [25, 26].
Co-culture systems are commonly used to study interactions between different cell populations. Some systems were found to enhance infection of pathogens as described in Tsun-Mei et al. in which they observed that coculturing monocytes with endothelial cells enhances the infection of Chlamydia Pneumoniae [4, 26].
Some cell lines synergize to develop a complementary, additivity or stimulatory relationship. Huh 7.5 supported HCV replication in vitro, this accounts for the lack of antiviral response induced by the Retinoic-Acid Inducible Gene I, however, these cells can’t polarize in culture which is necessary for compartmentalization of HCV receptors, HepG2 cells has the ability to polarize, although they lack CD81 which limits this cell line to support viral entry [7, 13].
Subgenomic replicons, and serum-based cell culture systems are available for genotypes 1, 2 and 3; however, genotype 4a is not well represented in cell culture models [3, 10, 14, 20, 22].
In the current study, we aimed at developing a long-term in vitro cell culture model, this model relies on co-culturing cell lines (Huh7.5-HepG2), and serum from HCV genotype 4a-infected patients as a source of infection.
Methods
Viral inoculation and cell culture
Blood samples were brought fresh from infected patients, serum was isolated on the same day then stored at
Infection protocol
At Day 0: When cells reach 60–80% confluency add 100
Huh 7.5 cells were used to optimize the inoculum volume of positive HCV serum for the infection protocol, 50 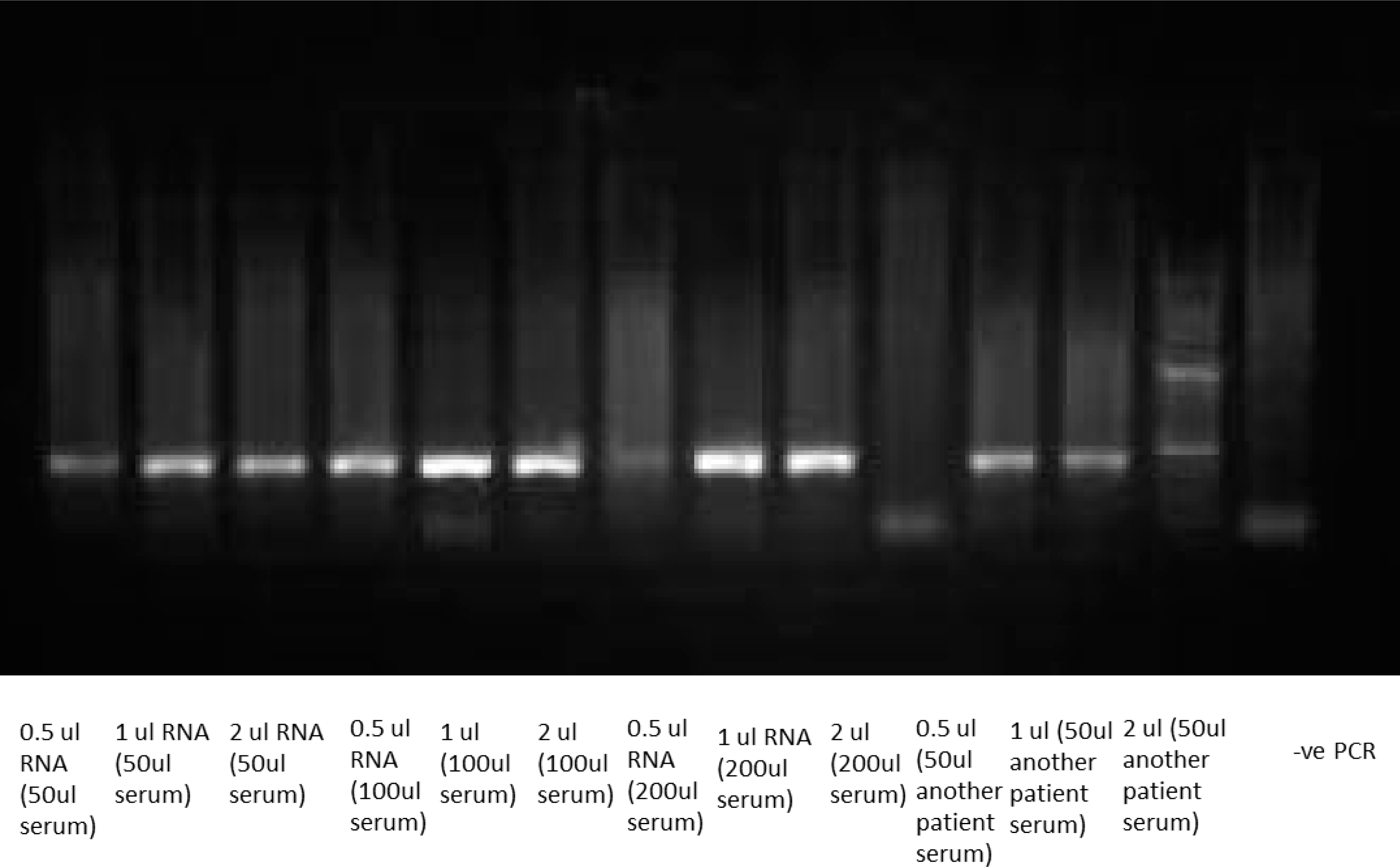
For qualitative studies for the detection of the viral RNA, reverse transcription PCR was performed as described in Lohr et al. mixture volume 25
Anti NS3 was used as an indicator for viral replication in Huh7.5 cells in which positive signal was detected after infection of cells using HCV genotype 4a positive serum.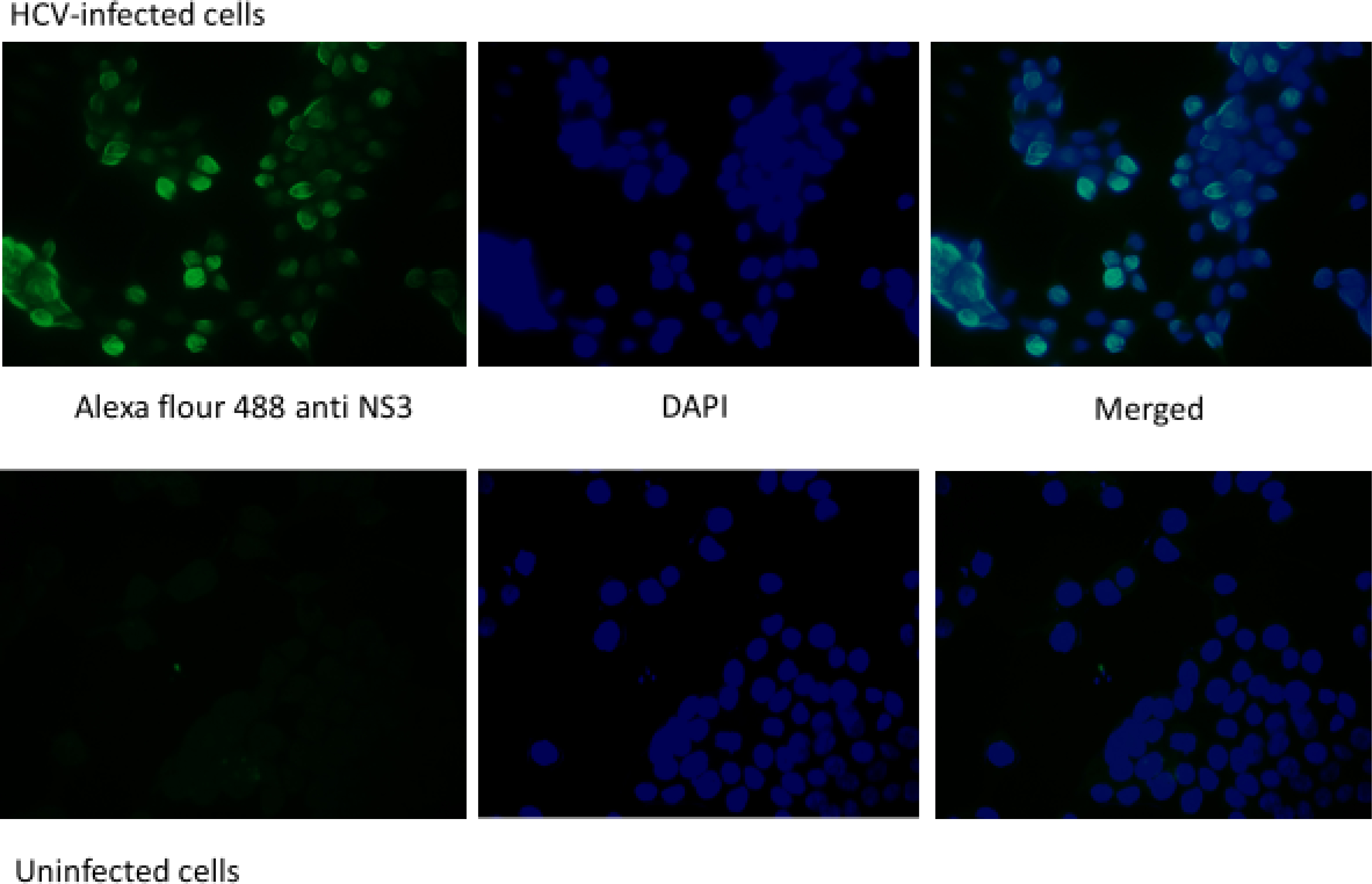
Real Time results for: Huh7.5, coculture of Huh7.5 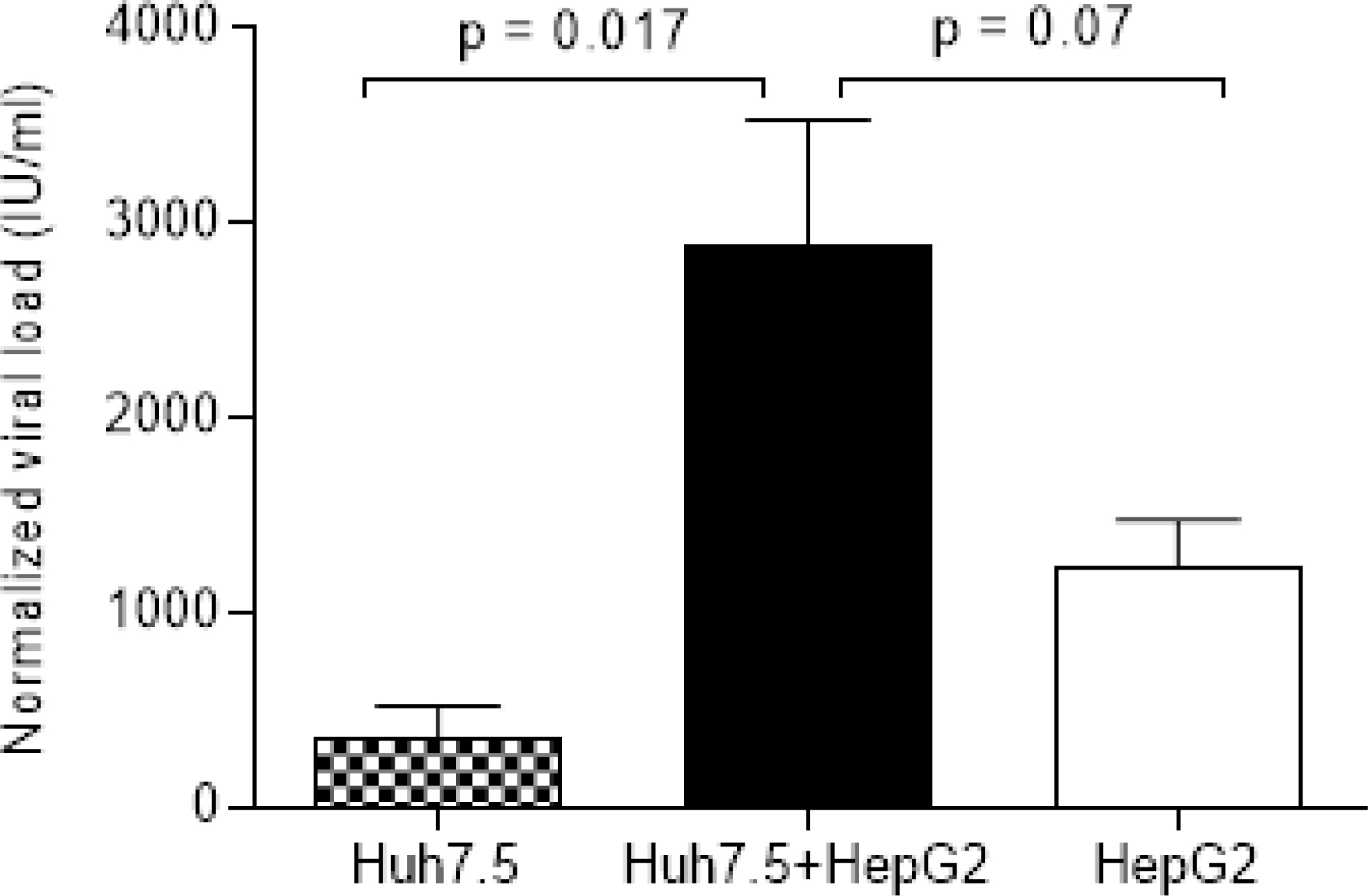
Plate layout for Huh7.5 on the left infected with 100 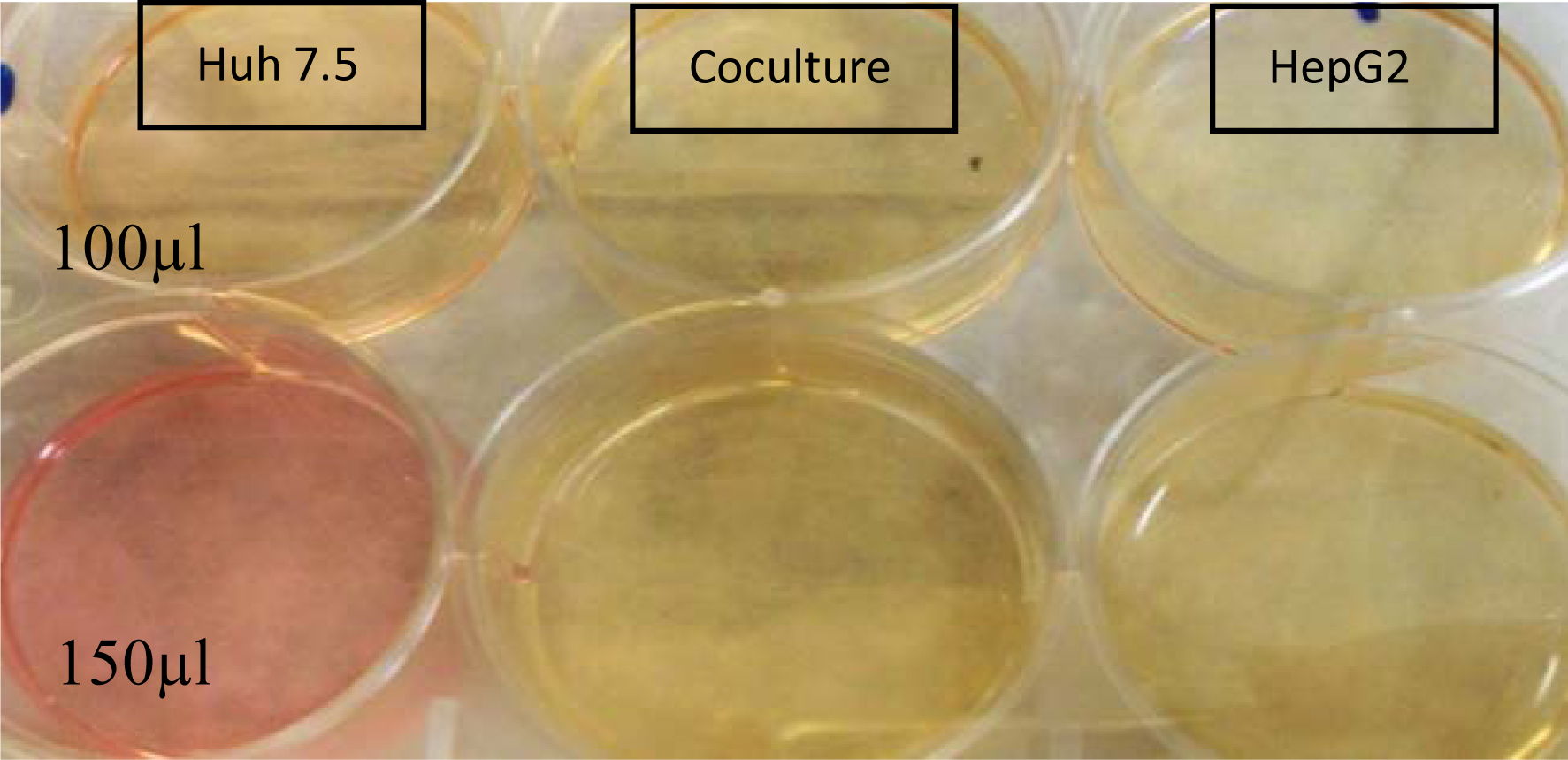
Viral RNA was isolated using the QIAamp Viral RNA Extraction Kit (Qiagen GmbH, Hilden, Germany) according to instructions by the manufacturer. Purified RNA was quantified by using the Artus HCV RT-PCR Quantification Kit (Qiagen GmbH) on Rotor Gene Real-Time PCR equipment following the manufacturer’s protocol.
Immune-staining protocol
For the immunofluorescence, Huh7.5 cells were seeded in a 24-well plate with coverslips into corresponding wells, infection protocol was followed, cells fixation with 4% paraformaldehyde for 10 minutes, washed twice with PBS, then added blocking buffer to reduce nonspecific binding, for 30–45 minutes. Incubated with primary antibody mouse anti human NS3 (invitrogen, US) 1 hour, then washed 3 times gently with PBS and incubated with Alexa Fluor™-(488) goat anti-mouse IgG 1 hour at room temperature. Then cells were washed and mounting media was added with Prolong™. Antifade (Invitrogen) and examined with a confocal Zeiss LSM 780.
Results
Infection optimization
We used positive serum samples with viral load between 10
Real Time results for: Huh7.5 alone, coculture of Huh7.5 
In order to estimate the optimal inoculum for infecting cells, we performed a titration with different infection inoculum (50, 100, 200
On a further validation for the infectivity results, and to confirm infectivity of serum we used NS3 protein as a biomarker for replication of HCV using serum of infected patients as a source of infection, immunostaining results (Fig. 2) shows positive fluorescence of HCV non-structural protein 3 (NS3).
Infection of coculture model
We infected cells with HCV positive serum (100, 150
Quantitative results of RT-PCR performed (Fig. 3) showed significant differences between coculturing Huh7.5 with HepG2 than individual cell lines, the cocultured condition infected with 100 and 150
Discussion
To gain insight into HCV pathogenesis and to facilitate the development of potential drugs against HCV, the establishment of an in vitro infection system which supports HCV replication is required. Many years of research showed that the establishment of an invitro replication model was very challenging, although in vivo infection is more successful as HCV causes chronic infection to 70% to 80% of cases. Ideally serum-derived HCV infection in hepatocytes should be efficient, however until now it is not clear why in vitro infection is not achievable, one of the suggested reasons is that the cells grown outside of their natural environment may produce more cytokines of an antiviral activity that resists infection [6]. Many groups attempted to infect using different cell models: MDBK, HELA, HEK-293 T, HepG2 and Huh-7 have been used to support HCV replication [3, 12, 15, 18] however, Huh-7 derived cell lines, are the most commonly used cells for studying liver associated diseases [1, 12, 14].
HuH-7 is a human hepatoma cell line isolated from a liver tumor in a 57 years old Japanese male in 1982. These cells typically grow in monolayers with slower growth rate than HepG2. HepG2 cells are hepatoma cells derived from a 15 years old male; These cells are also epithelial and are widely used in drug metabolism studies [1, 8, 20, 21, 24].
HepG2 cells are susceptible to HCV infection and supports its replication in vitro. Although Huh7 showed better susceptibility, but HepG2 cells enhance the expression of HCV structural proteins. They are also capable of shedding viral particles into culture media which in turn become infectious to uninfected cells [20].
Subgenomic replicon system is used widely as an infectious cell culture system for HCV-pathogenesis studies [18]. Despite of its effectiveness it cannot support the actual HCV replication cycle. Buck et al. and Molina et al. have infected human primary hepatocytes with HCV positive serum of genotype 1, 2, 3 and 4 and they were able to detect efficient HCV replication [6, 14, 20, 23].
In the present study, we used serum of HCV genotype 4a infected patients to infect Huh-7.5 and HepG2 cell lines. It is necessary to have full length HCV genotype 4a genome and to ensure the presence of essential factors that represent the micro environment for viral replication in the host. Interestingly, the co-culture model showed a significant increase in HCV RNA after 7 days of infection. The co-culture system and infection protocol described in our study differs from systems using JFH 1 virus for infection. It is also worth mentioning that we have tried depleting the serum of infected patients from IgG and Albumin to assess the change in the virus capacity for replication but no difference was observed (data not shown).
Huh7 and HepG2 cells are derived from well-differentiated hepatocellular carcinoma tissues. The results of our coculture system showed the possibility of expressing liver-specific factors that enhance complete HCV life cycle, however further studies are required to understand factors involved in this process. Origin of each cell line and the age of the cell line giver is one of the contributing factors towards the infectivity of the cells, which might contribute to the progression of hepatic cancer as well [1]. The relation between HCV replication capacity and hepatocellular carcinoma progression is not fully understood, although some studies have found that HCV-NS5A directly binds to P53 – a tumor suppressor protein-which in turns induces cellular proliferation. HepG2 cells carry wild type P53 while Huh 7 cells have a P53 point mutation [2, 12, 18]
It is challenging to mimic the liver environment for viral infection, however this coculture model might provide the complexity required for efficient replication. For future studies, it will be interesting to understand whether both cell lines synergize to induce higher replication capacity or one cell line is induced by the other. Synergistic effect is developed through either a complementary, additivity or stimulatory relationship. Although Huh7 cells vary greatly in supporting HCV infection, Huh 7.5 clone showed significant support for HCV replication in vitro, this is mainly accounted for the lack of antiviral response by the Retinoic-Acid Inducible Gene I, however the limitation for this cell line is the lack of polarity which is necessary for compartmentalization of HCV receptors, it is also thought that polarization impacts viral entry and release. HepG2 cells are known to polarize in culture, however they lack CD81 that limits viral entry. CD81 expressing HepG2 cells are proved to be a robust infection model.
Further studies should be performed on the protein level, to validate the expression of nonstructural proteins involved in the replication cycle of the virus by performing protein-based assays as western blot or ELISA, performing proteomics studies to recognize excreted proteins or cytokines from each cell line and in mixture will be beneficial. flow cytometry will help identify cell numbers for each cell line, this will be specifically useful for the co-culture condition in order to identify weather any cell line dominates over the corresponding one.
The established coculture model using Huh 7.5 and HepG2 cells, maintained HCV replication for at least 1 week after infection. This model will be useful in uncovering the interactions between HCV, human hepatocytes and hepatocellular carcinoma, this new invitro model will unravel the cross talking between both cell lines and can provide better understanding towards specific cytokines, growth factors and signaling pathways involved in the replication process of the virus and enhance the investigation of the complex relationship between HCV entry and hepatocellular carcinoma.
Conclusions
These findings report the first coculture model using Huh-7.5 and HepG2 cell lines for the support of HCV genotype 4a replication in vitro.
Authors contributions
M.M. Kamel designed and performed the experiments and manuscript writing; M.K. Ibrahim and R.M. Dawood contributed equally to this work by performing statistical analysis, critical revision to manuscript and data interpretation; All work was performed under supervision of A.B. Barakat and M.K. El-Awady.
Footnotes
Conflict of interest
The authors have no conflict of interest(s) related to this publication.