Recent studies have highlighted the role of natural elements in reduction of cancer cell growth and apoptosis. Koenimbine, a natural product isolated from Murraya koenigii (L) Spreng is a substance with cytotoxic effects on cancer cells.
AIM:
The effects of koenimbine on HT-29 and SW48 colon cancer cells were evaluated by MTT and Annexin V assays. Expression levels of Wnt/-catenin pathway genes were quantified by real time PCR.
RESULTS:
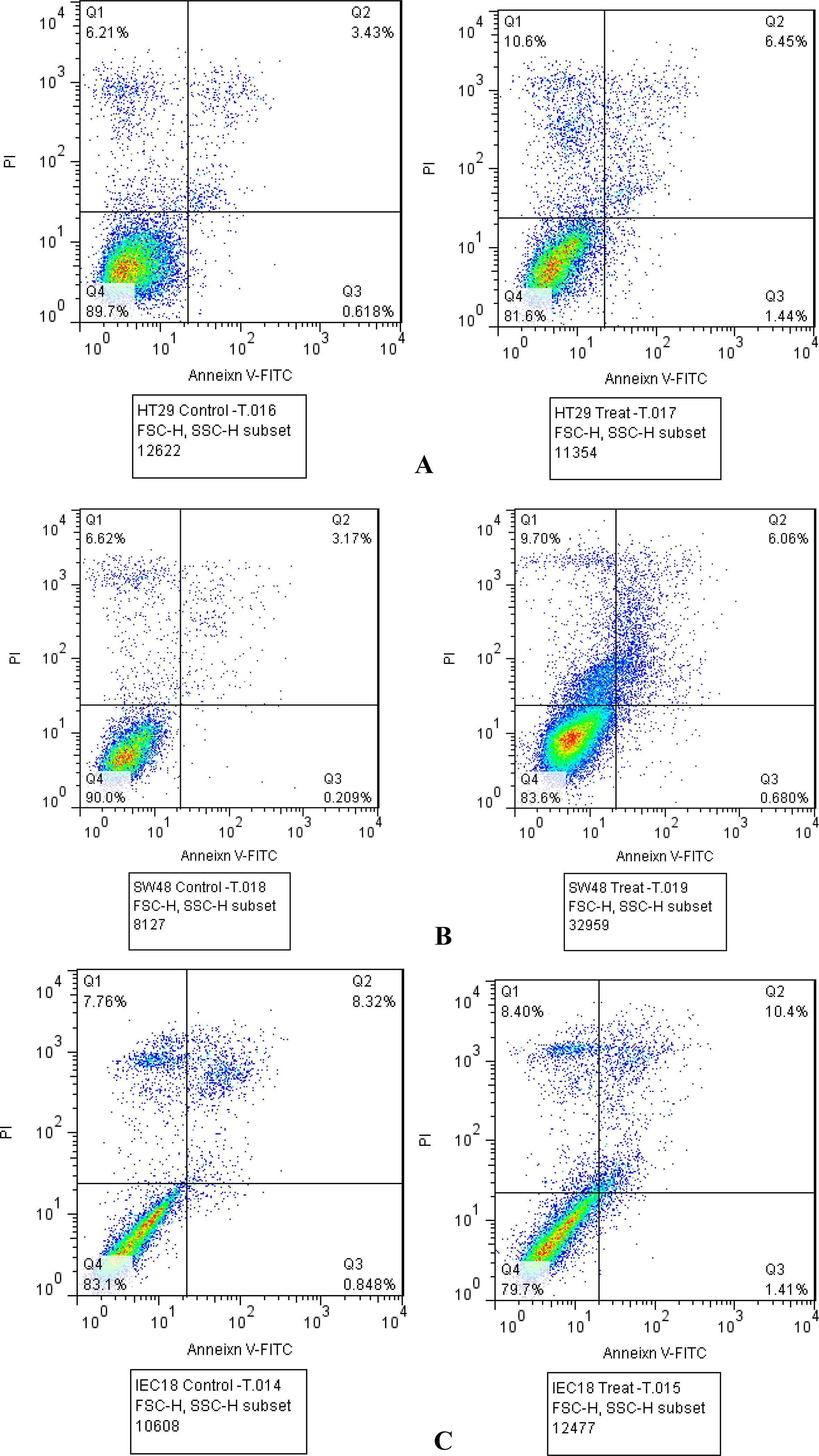
The IC50 values of koenimbine in HT-29 and SW48 was calculated to be 50 g/ml based on the results of MTT assay. This value was 75 g/ml in IEC-18 cells which were used as normal control. Annexin V assays revealed induction of cell apoptosis and necrosis in HT-29 and SW48 cells but not IEG18 cells by koenimbine. Koenimbin treatment resulted in significant down-regulation of CYCLD1 expression in SW48 cell line, but up-regulation of this gene in HT29 cell line. Expression of TBLR1, DKK1, GSK3B and -catenin was significantly decreased after koenimbin treatment in HT-19 cell line. Moreover, expression of DKK1 and GSK3B was significantly decreased after koenimbin treatment in SW-40 cell line. TCF4 expression was not detected in any of cell lines either before or after treatment with koenimbin.
CONCLUSION:
The current in vitro study showed the cytotoxic effects of koenimbin on two colon cancer cell lines and the effects of this substance on expression of selected genes from Wnt- catenin pathway. Future in vivo studies are needed before suggestion of this substance as an anti-cancer drug.
The Wnt signaling pathway has crucial roles in cell proliferation and survival and its over-activation has been demonstrated in almost all colorectal cancers [1]. This pathway modulates expression of -catenin via phosphorylation and ubiquitin-mediated destruction steps. The main components of the cytoplasmic -catenin degradation complex are AXIN, adenomatous polyposis coli (APC), casein kinase 1 (CK1), and glycogen synthase kinase 3 (GSK3). CK1 and GSK3 have roles in phosphorylation of -catenin and this process is accomplished only when Wnt is not present. Subsequent ubiquitination of -catenin leads to its destruction in the proteasomes. However, when Wnt ligand binds to the receptors Frizzled (FZD) and low-density lipoprotein-related protein 5/6 (LRP5/6), the -catenin degradation complex translocates to the membrane, and -TrCP is detached from the other components [2].
Several genes in this pathway including APC, -catenin, and/or AXIN2 are recurrently mutated in colorectal cancers [3]. Moreover, epigenetic inactivation of other regulators of this pathway such as SFRP1–5, WIF1, DKK1, and DKK3 has been reported in this kind of malignancy [2]. Taken together, Wnt pathway is regarded as a therapeutic target in colorectal cancer.
Koenimbine is an alkaloid extracted from leaves and fruits of Murraya koenigii (L) Spreng [4]. This substance has been shown to induce intrinsic apoptotic pathway in prostate and breast cancer cells [4, 5]. However, its effects on colon cancer cells have not been assessed yet. Based on the crucial role of Wnt pathway in the pathogenesis of colon cancer, in the current study, we evaluated the effects of koenimbine on viability of colon cancer cells and expression of a number of genes implicated in Wnt pathway. DKK1 was chosen based on the frequent epigenetic-mediated inactivation in colorectal cancer [2]. TBLR1 was selected according to its fundamental role in Wnt--catenin-mediated transcription and carcinogenesis [6]. Cyclin D1 (CCND1) has been recognized as a Wnt/-catenin downstream target with essential roles in the carcinogenesis [7]. GSK3 was chosen as a main component of cytoplasmic -catenin degradation complex [2]. Finally, TCF4 is one of the downstream effectors of Wnt/-catenin signaling. Moreover, certain TCF4 frameshift mutations have been reported in colorectal cancer cells which act as activating mutations [8].
Material and methods
Cell culture
The study protocol has been approved by the ethical committee of Shahid Beheshti University of Medical Sciences. Human colon cancer cell lines (HT-29 and SW48) were purchased from the Pasteur Institute, National Cell Bank of Iran. IEC-18 cells derived from epithelial cells of rat small intestine were used as controls. SW48 and HT-29 cells were cultured in RPMI 1640 and DMEM mediums respectively, plus 10% heat inactivated fetal calf serum, 1.5% 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) and 1%penicillin/streptomycin (all from Invitrogen, Carlsbad, CA, USA). IEC-18 cells were cultured in DMEM medium supplemented with 0.1 Unit/ml insulin in addition to the afore-mentioned materials. The cells were preserved as monolayer cultures at 37C in a humidified 5% CO situation for 24 hours to attach onto the dish.
MTT assay
The effect of koenimbine (The nature network, PhytoLab, Germany) on cultured cells was assessed using the MTT assay kit (Sigma, St. Louis, MO, USA). Each well had 10 cells seeded in 100 ml standard medium. After overnight incubation, different concentrations (1.56, 3.12, 6.25, 12.5, 25, 50, 75 and 100 g/ml) of koenimbine were added to wells. Plates were incubated at 37C under 5% (v/v) CO levels. Cell viability was calculated using the following formula:
Viability (percentage of the control) [(absorbance of the sample-absorbance of the blank)/[absorbance of the control- absorbance of the blank)] 100
Apoptosis assay
Cells were seeded in a 6 wells plate (3 wells for treated cells and 3 wells for controls). HT-29 and SW48 cells were treated with 50 g/ml of koenimbine, while IEC-18 cells were treated with 75 g/ml of this agent. After 24 h incubation, the supernatant (floating apoptotic cells) were collected and the adherent cells were treated with trypsin. The floating and trypsinized cells from each well were collected in a single tube. The collected cells were washed with PBS and centrifuged (300 g, 5 min). The supernatant was disposed and cells ( 1 10) were eluted in 100 L Annexin-binding buffer. Next, Annexin V and 5 L of propidium iodide (PI) were added to tubes. Tubes were incubated at room temperature for 5 minutes in the dark. Annexin V-FITC binding was evaluated by flow cytometry (Ex 488 nm; Em 350 nm) using FITC signal detector and PI staining by the phycoerythrin emission signal detector. Cells cultured without koenimbine were used as negative control.
Cytotoxic effects of different concentrations of Koenimbine on SW48, HT-29 (A) and IEC-18 cells (B).
Expression assays
RNA was extracted from cultured cells using TRIzol reagent (GeneAll, Korea). RNA quantity and quality was evaluated by Nanodrop 2000c spectrophotometer (Thermo Scientific, USA). Transcript levels of genes were quantified by qRT-PCR (Rotor Gene 6000, Corbett, Australia) after 24 hours treatment of cells with certain concentrations of koenimbine. PrimeScript RT reagent kit (Takara Bio, Japan) was used for cDNA production. SYBR Premix Ex Taq (Takara Bio, Japan) was used for expression analysis. The primer sequences are provided in the Table 1.
Nucleotide sequences of primers and the anticipated product sizes
Primer
Sequence
[l]Product
size (bp)
B2 MF
AGATGAGTATGCCTGCCGTG
105
B2 MR
GCGGCATCTTCAAACCTCCA
TCF4F
GCACTGCCGACTACAATAGG
150
TCF4R
CTGCATAGCCAGGCTGATTC
TBLR1F
GGGAGGAGAATGGAGCACAT
141
TBLR1R
CAGGGTTCCAGGCACAGATA
CCND1F
GAGGCGGAGGAGAACAAACA
180
CCND1R
GAGGCGGTAGTAGGACAGGA
CTNNB1 (-catenin) F
TCTGAGGACAAGCCACAAGATTACA
122
CTNNB1 (-catenin) R
TGGGCACCAATATCAAGTCCAA
GSK3BF
TCGAGAGCTCCAGATCATGAGAA
124
GSK3BR
CGGAACATAGTCCAGCACCAGA
DKK1F
CCAGACCATTGACAACTACC
120
DKK1R
CAGGCGAGACAGATTTGC
Statistical analysis
Transcript levels of genes were quantified in treated and control cells using the randomization test in the relative expression software tool (REST). Mann-Whitney test was applied for comparison of pretreated controls with inhibitory concentration 50% (IC50). All data were expressed as a mean SE of three distinct experiments. 0.05 was considered as statistically significant.
Results
MTT assay
The IC50 values of koenimbine in HT-29 and SW48 was calculated to be 50 g/ml based on the results of MTT assay. This value was 75 g/ml in IEC-18. Figure 1 shows the results of MTT assay in the assessed cell liens.
Apoptosis assay
Annexin V assays revealed induction of cell apoptosis and necrosis in HT-29 and SW48 cells but not IEG18 cells by koenimbine (Fig. 2).
HT-29 (A), SW48 (B) and IEG18 (C) cells were labeled with Annexin V-FITC after no treatment (left panels) and treatment with IC50 concentrations of koenimbine for each cell (right panels) (Q1: Necrosis, Q2: late apoptosis, Q3: early apoptosis, Q4: viability).
Relative expression of selected genes from Wnt/-catenin pathway in HT29 and SW48 cell lines after treatment with Koenimbin.
Expression assays
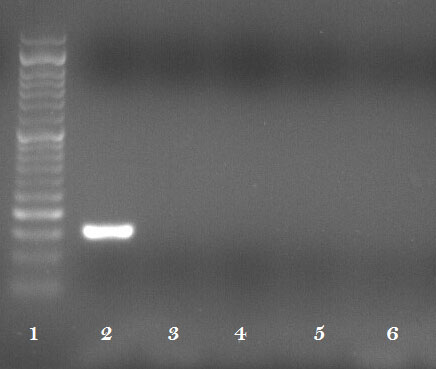
Figure 3 shows relative expression of selected genes from Wnt/-catenin pathway in HT29 and SW48 cell lines after treatment with Koenimbin. Koenimbin treatment resulted in significant down-regulation of CYCLD1 expression in SW48 cell line, but up-regulation of this gene in HT29 cell line. Expression of TBLR1, DKK1, GSK3B and -catenin was significantly decreased after koenimbin treatment in HT-19 cell line. Moreover, expression of DKK1 and GSK3B was significantly decreased after koenimbin treatment in SW-40 cell line. TCF4 expression was not detected in any of cell lines either before or after treatment with koenimbin (Fig. 4).
Expression of TCF4 in HT29 and SW48 cell lines after treatment with Koenimbin (Lane 1: size marker, lane 2: positive control, lanes 3 and 4: HT29 before and after treatment respectively, lanes 5 and 6: SW48 before and after treatment respectively).
Discussion
Wnt pathway has fundamental role in the carcinogenesis of colon cancer. Notably, the APC gene which is frequently mutated in colon cancer cells is negative regulator of this pathway. Several treatment strategies have been developed to target this pathway in colorectal cancer [2].
In the current study, we evaluated cytotoxic effects of koenimbine in colon cancer cell lines as well as a normal intestine cell line. The calculated IC50 values of the mentioned substance in cancer cell lines were significantly lower than the normal cell line. However, these values were higher than the reported values in prostate and breast cancer cell lines [4, 5]. Previous studies have shown that in both MCF-7 breast cancer cells and PC-3 prostate cancer cells, koenimbine has induced intrinsic apoptotic pathway [4, 5]. Consistent with these studies, we demonstrated that koenimbine induced cell apoptosis and necrosis in HT-29 and SW48 cells but not IEG18 cells. However, we did not assess the main route of apoptosis in this study. So, we propose conduction of further investigations to unravel which route of apoptosis is influence by koenimbine in colorectal cancer cells.
We also detected alterations in expression of Wnt- catenin genes after treatment with koenimbine. Koenimbin treatment resulted in significant down-regulation of CYCLD1 expression in SW48 cell line, but up-regulation of this gene in HT29 cell line. Expression of TBLR1 and -catenin was significantly decreased after koenimbin treatment in HT-29 cell line TBLR1 and -catenin have reciprocal intercations with each other recruiting each other to the promoter regions of the Wnt target genes to enhance expression of these genes and promote carcinogenesis [9]. Notably, TBLR1 is SUMOylated in a Wnt signaling-dependent fashion. When SUMOylated, it is relaesed from the corepressor complexes. On the other hand, deSUMOylation of this factor has been shown to inhibit the carcinogenic potential of SW480 cells [10]. Conseuqntly, further studies are needed to assess SUMOylation satatus of TBLR1 after treatment with koenimbine to precisely address the effct of this subsatnec in the carcinogenic process. Over-expression of CYCLD1 as a G1 phase marker has been assocaited with decreased overall survival of patients with colorectal cancer [11]. Therefore, the observed down-regulation of this gene in SW-40 after koeminbine treatmnet might be regarded as a potential useful effect of this substance for clinical application.
Expression of DKK1 and GSK3B was significantly decreased after koenimbin treatment in both cell lines. Suppression of GSK3-induced -catenin phosphorylation is an essential step in Wnt--catenin signaling. Moreover, GSK3 participates in Wnt signaling through phosphorylating the Wnt receptors LRP5/6 [12]. Investigations in several cancer types including colorectal cancer have shown that GSK3 inhibitors can inhibit cell proliferation [13]. Threrfore, the significant down-regulation of GSK3 after treatment with koenimbin is a promisiong result that should be further verified both in vitro and in vivo. However, the mentioned treatment has also decreased the levels of DKK1 which is regraded as a Wnt inhibitor. A previous study in SW480 cells has shown that hindering of secreted DKK1 in cell-conditioned medium eliminated -catenin downregulation [14]. Consequently, the altered expression levels of Wnt pathway genes in the current study does not have a consistent pattern to deduce the overall activity of this signaling pathway. Moreover, direction of such alterations is in favour of tumorigenesis. Consequently, further experiments are needed to elaborate the mechanism of action of koenimbin in colorectal cancer.
Footnotes
Acknowledgments
The current study was supported by a grant from Shahid Beheshti University of Medical Sciences.
Conflict of interest
Authors declare no conflicts of interest.
References
1.
SchatoffE.M.LeachB.I. and DowL.E., Wnt signaling and colorectal cancer, Current Colorectal Cancer Reports13(2) (2017), 101–110. PubMed PMID: 28413363. Pubmed Central PMCID: PMC5391049. Epub 2017/04/18.eng.
2.
NovellasdemuntL.AntasP. and LiV.S.W., Targeting Wnt signaling in colorectal cancer. A review in the theme: Cell signaling: Proteins, pathways and mechanisms, Am J Physiol-Cell Ph309(8) (2015), C511–C521. PubMed PMID: WOS:000364055500001.English.
3.
Comprehensive molecular characterization of human colon and rectal cancer, Nature487(7407) (2012), 330–337. PubMed PMID: 22810696. Pubmed Central PMCID: PMC340 1966.Epub 2012/07/20.eng.
4.
KamalidehghanB.Ghafouri-FardS.MotevaseliE. and AhmadipourF., Inhibition of human prostate cancer (PC-3) cells and targeting of PC-3-derived prostate cancer stem cells with koenimbin, a natural dietary compound from Murraya koenigii (L) Spreng, Drug Des Dev Ther12 (2018), 1119–1133. PubMed PMID: WOS:000431643900003. English.
5.
AhmadipourF.NoordinM.I.MohanS.AryaA.PaydarM. and LooiC.Y. et al., Koenimbin, a natural dietary compound of Murraya koenigii (L) Spreng: Inhibition of MCF7 breast cancer cells and targeting of derived MCF7 breast cancer stem cells (CD44 (+)/CD24 (-/low)): An in vitro study, Drug Design, Development and Therapy9 (2015), 1193–1208. PubMed PMID: 25759564. Pubmed Central PMCID: PMC4346015. Epub 2015/03/12.eng.
6.
LiJ. and WangC.Y., TBL1-TBLR1 and β-catenin recruit each other to Wnt target-gene promoter for transcription activation and oncogenesis, Nature Cell Biology10(2) (2008), 160.
7.
MohammedM.K.ShaoC.WangJ.WeiQ.WangX.CollierZ. et al., Wnt/β-catenin signaling plays an ever-expanding role in stem cell self-renewal, tumorigenesis and cancer chemoresistance, Genes & Diseases3(1) (2016), 11–40.
8.
Cuilliere-DartiguesP.El-BchiriJ.KrimiA.BuhardO.FontangesP.FlejouJ.F. et al., TCF-4 isoforms absent in TCF-4 mutated MSI-H colorectal cancer cells colocalize with nuclear CtBP and repress TCF-4-mediated transcription, Oncogene25(32) (2006), 4441–4448. PubMed PMID: 16547505. Epub 2006/03/21.eng.
9.
LiJ. and WangC.Y., TBL1-TBLR1 and beta-catenin recruit each other to Wnt target-gene promoter for transcription activation and oncogenesis, Nature Cell Biology10(2) (2008), 160–U36. PubMed PMID: WOS:000252897800011. English.
10.
ChoiH.K.ChoiK.C.YooJ.Y.SongM.KoS.J.KimC.H. et al., Reversible SUMOylation of TBL1-TBLR1 regulates beta-catenin-mediated Wnt signaling, Molecular Cell43(2) (2011), 203–216. PubMed PMID: 21777810.Epub 2011/07/23.eng.
11.
BahnassyA.A.ZekriA.R.N.El-HoussiniS.El-ShehabyA.M.R.MahmoudM.R.AbdallahS. et al., Cyclin A and cyclin D1 as significant prognostic markers in colorectal cancer patients, Bmc Gastroenterol23 (2004), 4. PubMed PMID: WOS:000224726100001.English.
12.
WuD.Q. and PanW.J., GSK3: A multifaceted kinase in Wnt signaling, Trends Biochem Sci35(3) (2010), 161–168. PubMed PMID: WOS:000275912600006.English.
13.
MiyashitaK.NakadaM.ShakooriA.IshigakiY.ShimasakiT.MotooY. et al., An emerging strategy for cancer treatment targeting aberrant glycogen synthase kinase 3 beta, Anti-Cancer Agents in Medicinal Chemistry9(10) (2009), 1114–1122. PubMed PMID: 19925395.Epub 2009/11/21.eng.
14.
JumpertzS.HennesT.AsareY.SchutzA.K. and BernhagenJ., CSN5/JAB1 suppresses the WNT inhibitor DKK1 in colorectal cancer cells, Cellular Signalling34 (2017), 38–46. PubMed PMID: 28229932.Epub 2017/02/24.eng.