
Abstract
MiRNAs (microRNAs) are defined as micro directors and regulators of gene expression. Since altered miRNA expression is signified in the pathobiology of diverse cancers such as colorectal cancers (CRCs), these molecules are described as therapeutic targets, either. Manipulation of miRNAs could lead to further therapy for chemo and radio-resistant CRCs. The usage of microRNAs has indicated prominent promise in the prognosis and diagnosis of CRC, because of their unique expression pattern associated with cancer types and malignancies. Nowadays, many researchers are analyzing the correlation between miRNA polymorphisms and cancer risk. With continuous incompatibility in colorectal cancer (CRC) miRNAs expression data, it is critical to move toward the content of a “pre-laboratory” analysis to speed up efficient accuracy medicine and translational study. Pathway study for the highest expressed miRNAs- regulated target genes resulted in the identification of a considerable number of genes associated with CRC pathway including PI3K, TGF
Introduction
Colorectal cancer (CRC) with one million new cases and more than 600,000 annual deaths is still one of the five most leading cause of cancer worldwide [49, 94]. In addition to the sporadic type, it has hereditary colorectal polyposis form which consists of various disorders passed on to the generations via autosomal dominant inheritance and is classified as familial adenomatous polyposis (FAP) and Lynch syndrome [12]. The latest syndrome is known as the lack of polyposis [48], a positive family history with susceptibility for extracolonic malignancies such as gastric, ovarian, and endometrial cancers [143]. CRC is a multifactorial disorder that could be under genetics, environmental factors, and gene/environmental interactions influence based on its etiology. Genetic and epigenetic abnormalities are participated in a compound network and may both predispose or lead to the progression of each other (Fig. 1) [20, 25, 105, 124, 141].
Mechanisms associated with CRC. Genetic and epigenetic changes (DNA methylation, Histone modification, Chromatin alteration, etc.) involved in modulation of gene expression patterns. Silencing of TSG and/or Activation of Oncogene is developing CRC. miRNAs, which represent mutual interconnections with epigenetic mechanisms, would be straightly regulated via epigenetics either by histone modifications or by atypical methylation in the promoter region [9, 10]. * MiRNA: MicroRNA/TSG: Tumor Suppressor genes/CRC: Colorectal Cancer (Illustrated by the first author).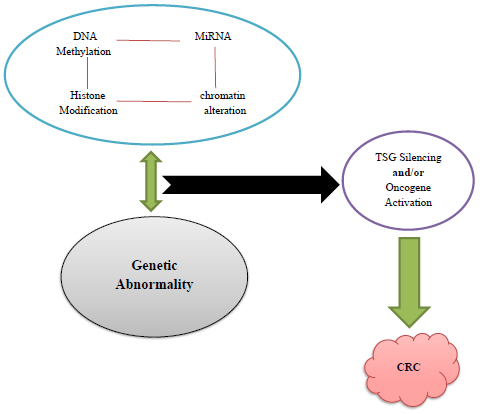
Understanding the molecular process of CRC is a crucial step in the recognition of novel molecular targets that could be effective in describing the prognosis of CRC patients and find their appropriate therapy [107]. Its processes of genesis contain a succession of molecular pathways “such as Wnt pathway” and abnormalities in key regulatory genes “including p53” from adenoma to carcinoma and eventually to metastatic disease [2].
The Hippo signaling pathway “a novel tumor suppressor pathway” (Fig. 2) is well conserved in various species [3, 42]. Whilst the Hippo pathway is active, its downstream oncogene YAP1 and the associated TAZ are phosphorylated and inactivated with the aid of the Hippo core complex [100]. When the Hippo pathway is suppressed, YAP1 and TAZ are un-phosphorylated; therefore, they enter the nucleus and promote transcription of genes which involved in proliferation [127].
Hippo signaling pathway. Working theory for the function of the Hippo pathway and YAP during tumor development and metastasis. The activation of the Hippo signaling pathway by modifies in cell shape, cell adhesion, and cell density causes phosphorylation of the Hippo kinases (MST1/2 in mammals), which is in fusion with the adaptor protein Sav phosphorylate LATS1/2 kinases and their associate, MOB. The LATS/MOB complex then phosphorylates YAP “the transcriptional co-activator” and therefore suppresses YAP function with advancing both cytoplasmic sequestration by 14-3-3 proteins and proteasomal degradation. Previous studies report that suppressing the capability of the Hippo signaling pathway to suppress YAP results in enhanced YAP/TEAD-dependent gene expression, which impresses both tumor growth and metastasis via increasing processes that take place at both the primary tumor and at the metastatic area (Illustrated by the first author).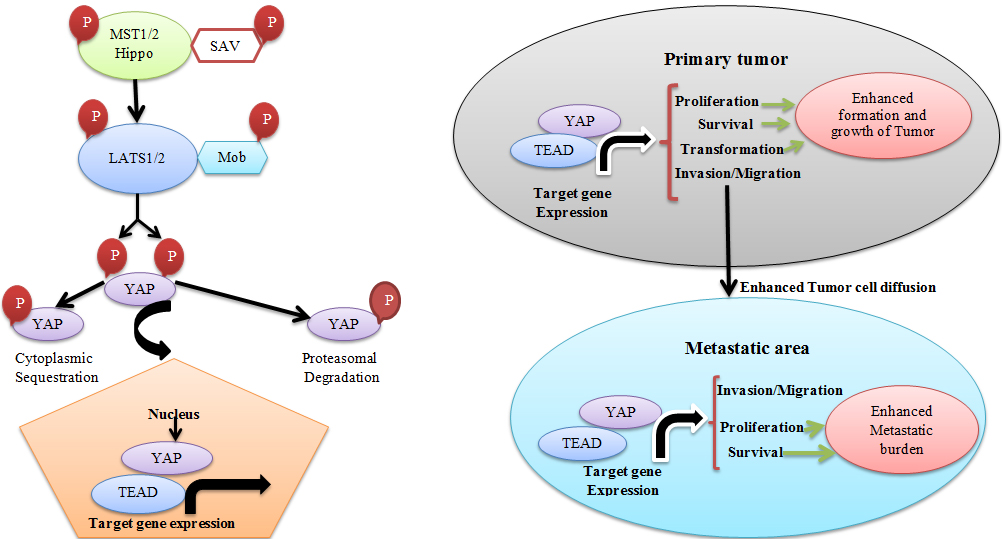
Deregulation of these genes has been reported in many human cancers, including CRC [39, 54, 126, 128, 129, 148]. Moreover, YAP1 and TAZ play significant roles in the progression of CRC by their overexpression, which induce proliferation of CRC cells [3, 42, 126, 128, 152].
Three elements of the Hippo pathway, TEAD4, CTGF, and YAP1, were appeared to be direct targets of miR-375 [13]. The expression of these genes revealed a negative connection with miR-375 expression and YAP1 re-expression relatively nullified the tumor-suppressive efficacy of miR-375 [53].
TEAD transcription factors and YAP1 a transcription co-activator, are the most important binding accomplice for YAP1; together they apply oncogenic functions in tumorogenesis [83] although; the downstream rules of TEAD/ YAP1complex in Gastric Cancer are unknown [11]. Moreover, the evidence demonstrates that the Hippo-YAP1 pathway is under the influence of deregulated microRNAs (miRNAs) [55, 155].
Genomic instability (GI) is known as an essential mechanism for CRC progression and is ascribed to microsatellite instability (MSI) and chromosomal instability (CIN) [99]. CIN is a major deficiency observed in about 80% of CRC cases, which leads to gain or loss of whole chromosomes or chromosomal segments and the abnormality of critical genes such as adenomatous polyposis coli (APC), p53 and Ras. In MSI tumors, an imperfection in mismatch repair genes (MMR) is reported [21, 81, 124]. Many researchers tried to discover genetic alterations as susceptible biomarkers for CRC prognosis and diagnostic evaluation.
Single nucleotide polymorphisms (SNPs) in miR-219-1 and miR-608 were observed to be correlated with CRC survival and recurrence, and probably helpful in predicting therapy response [5].
MicroRNAs (miRNAs) are endogenous short noncoding single-stranded RNA sequences with 18 to 25 nucleotides which are involved in various biological processes such as differentiation, growth, proliferation, and apoptosis capable of binding to target mRNAs’ complementarity sequences for post-transcriptional regulation of their functions as oncogenes or tumor suppressors [32, 35, 37, 38].
The detection of miRNAs has opened a new conception into CRC management. miRNAs are genetically linked to the progression of different cancers such as CRCs based on their altered expression patterns [140]. The eventual mechanisms of miRNA deregulation are mutation and epigenetic changes [36, 145]. In CRCs, a feature of plasma miRNAs could be used for estimating patient prognosis and the effect of therapeutic medicines [111].
This article investigates the methodological approaches for the roles of miRNAs in the pathobiology of CRCs and their potential clinical efficiencies.
MiRNAs interact with various cancer pathways and manages cellular homeostasis, therefore, suppressing overexpressed oncogenic miRNAs [28]. Also restoring downregulated tumor suppressor miRNAs could finally lead to tumor development, progression, inhibition, and apoptosis. The miRNAs associated with CRC are shown in Table 1 [4, 9, 14, 16, 30, 31, 41, 47, 51, 60, 61, 66, 76, 91, 95, 97, 102, 103, 110, 116, 117, 118, 123, 125, 129, 131, 134, 135, 142, 147, 151, 156].
The associated miRNAs with CRC
The associated miRNAs with CRC
The distinguished oncogenic miRNAs in CRC
Oncogenic and tumor suppressor miRNAs and their predicted target genes are shown in Tables 2 and 3 [16, 17, 24, 31, 34, 46, 51, 70, 71, 77, 89, 90, 101, 112, 114, 115, 123, 125, 133, 135, 137, 138, 139, 144, 146, 150]. Substantial strengths of miRNA relevant research: The capability for a remarkable complementary method to evaluate CRC risk, determining responses to chemo-and radio-therapies and pharmacologic interventions through designing targeted medications to decrease CRC risk and/or recurrence cutback [121].
The identified tumor-suppressive miRNAs in CRC
Utilization of miRNAs using synthetic miRNA or chemically altered oligonucleotides would change biogenesis, maturity, functionality and binding sites on the target mRNAs [109]. Although a major impediment to the usage of these molecules for therapeutic aims is the occurrence of nonspecific side effects, either lead to cross-sensitivity to endogenous miRNAs and/or additionally nonspecific binding sites to noncancerous cells [79]. The utilization of a manipulated, site-specific delivery system and synthesis of less toxic anti-miRNA oligonucleotides may also reduce such side effects [6].
Similar modified miRNA expression patterns between different tumor types present their effects in the cellular pathways changes during cancer pathogenesis [10]. However, these aberrant forms of miRNA expression that are in conjunction with diverse cancers are well known except that it remains to better understand the functional consequence of such miRNAs dysregulation on their targets [82]. Further attempts are required to elucidate the role of miRNAs in proliferation, differentiation, and apoptosis.
The roles of miRNAs in carcinogenesis and tumor development have been proven by multiple functional studies in the past decades [108]. Via the suppression of target messenger RNAs, microRNAs regulate several cellular pathways that are involved in proliferation, differentiation, and apoptosis [22].
MiRNAs may positively or negatively impress the function of the tumor-suppressor P53 gene (TSG), which is momentous in controlling cell cycle and apoptosis in CRC [1].
For instance, miR-96 upregulation is reported in CRC and is discovered to target p53 inducible nuclear protein 1 (TP53INP1), exerting down regulatory influences on p53 function [34].
On the other hand, miR-34a has been reported to enhance p53 expression via its inhibition of Sirtuin-1 (SIRT1) [84].
MicroRNAs received valuable consideration due to their dysregulation in CRC [15]. Aberrant expression of many miRNAs has been proven to be associated with treatment response in CRC patients [120]. An increasing amount of dysregulated miRNAs showed to have correlations with sensitivity or drug resistance which reveals their ability of forecasting patients’ responses to some anticancer factors [44].
Researchers have previously identified the altered expression of 13 miRNAs in patients affected by CRC [33] and miRNA expression alterations in CRCs with either BRAF or KRAS mutations that demonstrate these modified expressions could be related to miRNAs’ regulatory action in the RAS signaling pathway [75]. The upregulation of miR-31 was reported to be correlated with stage IV CRC [136].
Downregulation of miR-143 and miR-145 was shown in precancerous adenomatous polyps in comparison with normal tissue indicating that these miRNAs play curial roles in the primary progression of the tumors [92].
Interestingly, previously studied notable upregulations of miR-20, miR25, miR-17-5p, miR-17-92, miR-92-1, miR-92-2, miR-93-1 and miR-106a in the microsatellite stable (MSS) CRC but not in MSI CRC [58]. More studies demonstrated an enhanced expression of miR-17-5p, miR18a, miR-20a, miR-31, miR-92 and miR-183 in tumor tissue in comparison with normal colorectal mucosa where a correlation between worse CRC prognosis and high miR-18a was observed [43, 92].
Lately, the expression of miR-203 has been found to be related to poor survival in Caucasians with stage IV CRC and significantly, it is an indicator of poor survival in blacks with either stage I or II CRCs [8]. Eventually, the expression of miR-21 predicted a poor prognosis in CRC patients with stage IV [9].
Research shows that tumor suppressor and oncogenic miRNAs make a promising potential to be manipulated for clinical tests such as CRC therapy, blocking the development of precursor lesions, prevention of distant metastasis, and improving responses to chemo- and radio-therapies [94]. In addition, fast degradation of miRNAs or anti-miRNAs by cellular nucleases and poor knowledge of cellular mechanisms are other disadvantages of these clinical trials [68]. Thus, utilizing the lowest optimum concentration of miRNAs accompanied by impressive delivery systems which include viral and non-viral vectors and nanoparticles may also limit such side effects and express the dose-dependent accumulation of targeted vectors in CRC cells [119].
The investigation of the pathway of the regulated target genes by the most highly expressed miRNAs discovers a considerable number of genes associated with CRC development including PI3K, TGF
a: CRC pathway and highly expressed miRNAs. The data was taken from KEGG database, the miRNAs showed with black font to demonstrate the targets regulated by these miRNAs (Illustrated by the first author). b: CRC pathway and highly expressed miRNAs. Pathway investigation outcomes for the target genes of the most highly expressed miRNAs in CRC pathway. The data was taken from KEGG database, the miRNAs showed with black font to demonstrate the targets regulated by these miRNAs (Illustrated by the first author).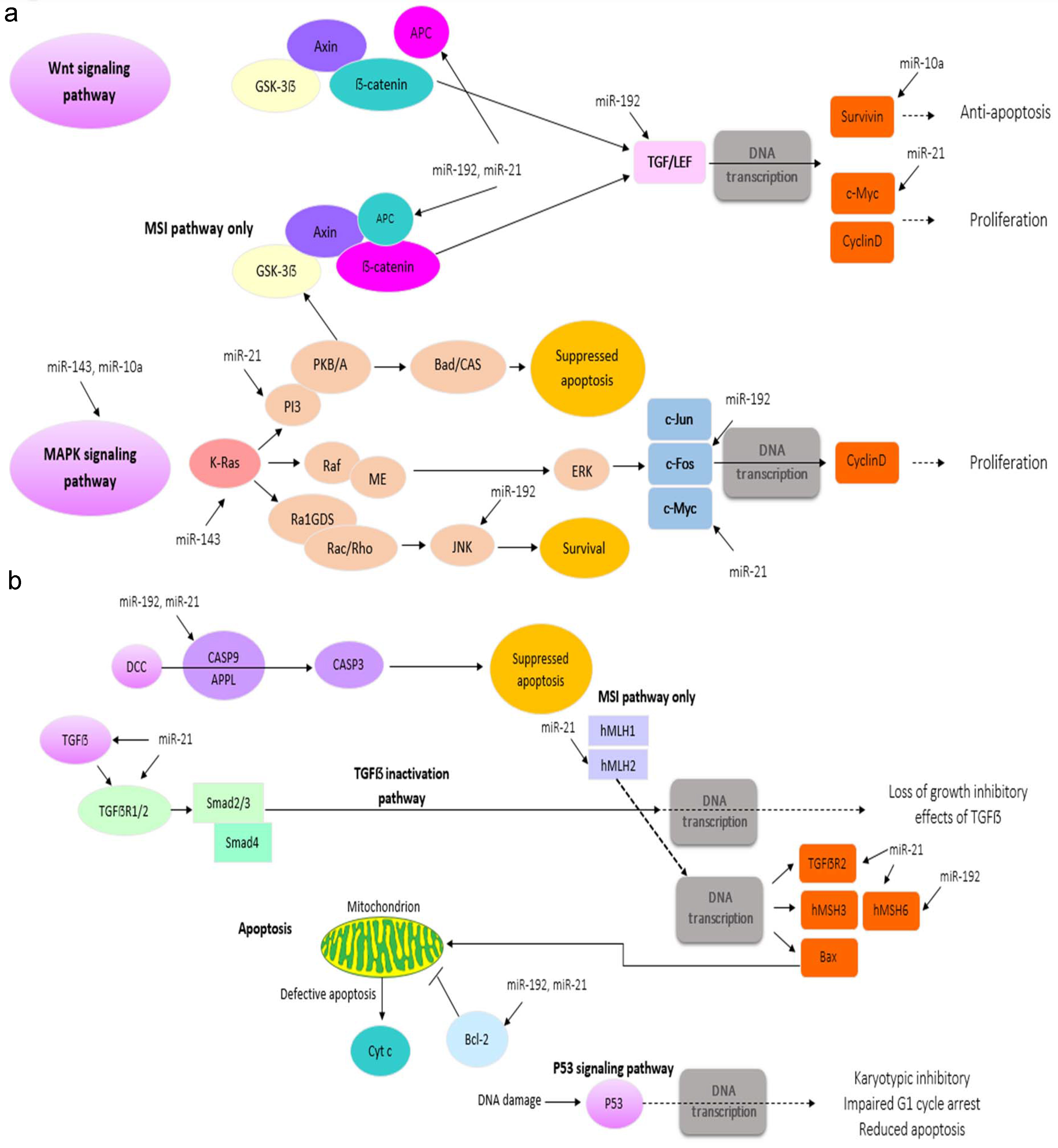
Human genome SNPs have been shown to affect cancer susceptibility [27]. Functional SNPs in miRNAs’ promoter regions could change miRNAs’ expressions or impress mRNA activity, potentially contributing to cancer risk [87]. Thus, miRNA polymorphisms may be used as potential markers of susceptibility for CRC prevention and diagnosis [96].
miRNAs may play a tumor-suppressive or oncogenic role in their regulation of signaling pathways leading to cancer development [74].
Oncogenic miRNAs, so-called oncomiRs, commonly target and downregulate endogenous tumor-suppressor genes [50]. On the other hand, tumor-suppressive miRNAs play a momentous role in downregulation genes associated with development and metastasis [57].
The downregulation of tumor-suppressive miRNAs and the upregulation of oncomiRs have notable effects on the progression of cancer [153]. Inactivation of Adenomatous polyposis coli (APC), leading to activation of the Wnt pathway through free
As mentioned previously, miRNAs play a significant role in the progression of the metastatic form of tumors, and as a result, these signs may be used as diagnostic biomarkers [40]. Hur et al. discovered a miRNA expression pattern including let-7i, miR-10b, and miR-885-5p that is particularly seen in CRC metastasis [45]. MiR-552 and 592 levels could also discriminate between primary lung cancer and lung metastasis secondary to CRC [56]. One of the currently used biomarkers for CRC metastasis is the Carcinoembryonic antigen (CEA). Researchers have found that miR-141 could be used in association with CEA to increment its predictive abilities [19, 52].
Having a migratory and invasive phenotype by cancer cells is an essential point in the progression of metastasis [59]. As previously is reported, one of the most highly upregulated miRNAs in CRC is miR-21 which downregulates many genes that are involved in migration and invasion processes, such as those of PTEN, PDCD4, SPRTY and TIAM1 [24, 26, 133].
Overexpressed targeting miRNAs in CRCs could not only stop the recurrence of tumor formation but also could regulate the development of progressive metastatic tumors [64]. Such miRNA alterations would be possibly used as an alternative therapy in chemo and radioresistant CRC patients [130]. The experimental patterns of miRNAs’ expression have to be validated for both preclinical and clinical settings with the intention to advance a miRNA-based therapy for CRCs [130]. The comprehension of gene regulation, and its correlation to cancers mainly CRCs, is enhanced after the recognition of miRNAs [149]. The dysregulation of miRNAs is near conjunction with cancer pathobiology [18].
MiRNAs have the capability of clinical efficiency in primary diagnosis, in estimating prognosis, and in predicting the effectiveness of therapy, especially in CRC patients [98]. The CRC-specific miRNAs wait for major validation in larger prospective groups previous to implementation in clinical oncologic practice [113].
Targeting miRNAs for CRC therapy
The fact that miRNAs play remarkable roles in CRC development establishes a rationale for CRC therapeutic researches [157]. The participation of miRNAs in the pathophysiology of CRC shows prominent support for their usage in novel therapeutics [104].
As mentioned before, many currently used chem-otherapeutic medicines demonstrate altered effectiveness belonging to the expression levels of certain miRNAs [86]. It seems that this information would be used via the introduction of miRNA suppressors to develop drug response [63]. For instance, miR-143 has been discovered to enhance CRC cell sensitivity to 5-FU [73]. The recovery of downregulated tumor suppressor miRNAs in CRC patients could provide a considerable therapeutic advantage [119].
Many miRNAs have demonstrated anti-tumorigenic impresses, therefore, it seems that at least some of these molecules are suitable as potential therapy tools with some roles in tumorigenic signaling pathways [74]. For instance, miR-26b has been reported to target many various oncogenic genes in CRC [67]. MiRNAs have also represented promise in the improvement of drug targeting in particular novel signaling pathways associated with cancer [119]. Also, Anti-angiogenic therapies are being used more often in CRC management [80]. Bevacizumab is a commonly approved monoclonal antibody that has anti-angiogenic influences through targeting VEGF [119].
It has been reported that miR-126 has also therapeutic potential in CRC via its targeting VEGFA and its following regulation of angiogenesis [112]. On the other hand, miRNA substitution therapy strives to enhance the influences of tumor-suppressive and drug sensitizing miRNAs via the introduction of similar miRNA molecules [69]. This would be accomplished via a gene substitution therapy analogous to that used for small interfering RNAs (siRNAs), which might be providing fewer problems than gene treatment with protein-coding genes [62].
Increasingly more dysregulated miRNAs have been disclosed to have conjunction with drug resistance or sensitivity which implies their functionality of predicting patients’ responses to some anticancer factors [44].
Conclusion
The miRNA-based gene therapy is often based on both negative and positive miRNA expression alterations. Technologies that combine RNA sequencing, system biology, and proteomics would allow a comprehensive evaluation and perception of miRNA function which prepare thrilling chances for new pathogenetic and treatment intuition into CRC management. Novel therapeutic tactics will face the crucial challenge of improving standardized principles for miRNA prevention that integrate great transfection efficiency with targeted delivery.
The special capabilities of microRNAs to influence numerous downstream signaling pathways demonstrate a novel approach for CRC therapy. However, still, early in its improvement, we believe that microRNAs could be used as biomarkers and therapeutic targets for CRC in the future.
Footnotes
Conflict of interest
All the authors listed have approved the manuscript and no conflict of interest exists.