
Abstract
Recent advances in assisted reproductive technology (ART) have allowed couples with severe infertility to conceive, but the methods are not effective for all cases. Stem cells as undifferentiated cells which are found in different stages of embryonic, fetal and adult life are known to be capable of forming different cell types, tissues, and organs. Due to their unlimited resources and the incredible power of differentiation are considered as potential new therapeutic biological tools for treatment of infertility. For reproductive medicine, stem cells are stimulated in vitro to develop various specialized functional cells including male and female gametes.
The epigenetic patterns can be modified in the genome under certain drugs exposure or lifestyle alterations. Therefore, epigenetics-related disorders may be treated if the nature of the modifications is completely admissible. It is proved that our understanding of epigenetic processes and its association with infertility would help us not only to understand the etiological factors but also to treat some type of male infertilities.
Exploration of both genetic and epigenetic variations in the disease development could help in the identification of the interaction patterns between these two phenomena and possible improvement of therapeutic methods.
Introduction
Around 15% of couples suffer from fertility problems [25]. Infertility is revealed when the relevant investigations and probable treatments initiate. It is defined as a failure to conceive a clinically detectable pregnancy after 12 months of unprotected intercourse [37]. The disorder is a heterogeneous condition and is caused by various pathologic conditions and different underlying mechanisms [63].
Genetic factors make a small contribution to infertility. Therefore, in order to fully understand the causes of infertility, another contingency factor should be considered. Epigenetics is a phenomenon that is known as a major cause of various diseases, including infertility [74]. This phenomenon includes events such as DNA methylation, histone post translational modification, and chromatin remodeling. The epigenetic changes may be short or long-term [9] and play a fundamental role to define the involved genes and the precise expression time of these genes in specific cells [20]. Even though the genetic code is thought to be unmodified in every cell during an organism’s entire life, it is believed that the epigenetic code is dynamic and tissue-specific [57]. The variable arrangement of epigenetic deviations can be induced and provide irregular epigenetic amendments which have been applied as therapeutic goals for a number of diseases [52]. Maintaining a proper guideline of epigenetic processes during spermatogenesis is not only important to confirm appropriate sperm function but also for proper embryonic development [7, 50]. It has been found that the sperm epigenetic setting plays a serious role in creating epigenetic marks in the embryo, thus unusual epigenetic regulation in spermatogenesis has an adverse effect on both male fertility and embryonic development [27, 29, 58].
Stem cells are undifferentiated cells existing in the embryonic, fetal, and adult stages of life and convert them to differentiated cells to create the building blocks of tissue and organs. Stem cells are considered as potent new therapeutic biological tools for infertility treatment because of unrestricted origin and high variation capacity [34, 66].
Recently it is shown that for developing male and female gametes, as well as other specialized cells for reproductive medicine, “stem cells” could be stimulated in vitro [4, 43]. Lately a significant advancement in origin of male germ cells from pluripotent cells has been completed [35]. Also, stem-cell based tactics for ovarian rejuvenation and oocyte production have been proposed as forthcoming cell remedies for treating sterility in women [19, 55]. In recent years several improvements have been made in assisting reproduction treatment. More than 80% of the couples who suffer from infertility may have a chance of conceiving a child [15].
Current treatments for infertility
Infertility is a global incident prevalent in up to 15% of couples [1]. Almost 40% of couples, both female and male factors are causative factors in infertility. Infertility has vital public health subsequences, consisting of, economic limitation, psychological distress, social stigmatization and later onset adult disorder in both women and men, beyond pregnancy or its deficiency/absence [2, 3, 40, 71]. If male factors are the cause of a couple’s infertility, some limited and invasive procedures can be employed for treatment [49]. When a sperm sample is obtained from the male patient, IVF can be performed by either incubation of the sample with a partner’s oocyte or by its intra-cytoplasmic sperm injection (ICSI). If the functional spermatozoa or elongated spermatids ICSI is uncommon, three invasive techniques can be used to acquire samples appropriate for ICSI, including testicular biopsy, testicular sperm removal or percutaneous sperm aspiration [47]. The recent technique has been widely successful in isolating testicular spermatozoa capable of fertilizing a partner’s oocyte even for highly azoospermic cases, including men with Klinefelter syndrome [18]. All assisted reproduction options are highly invasive and often involve hormone therapies to stimulate ovulation which leads into ovarian hyper-stimulation syndrome due to increased mediated vascular permeability by vasoactive substances and other additional health concerns [30]. These concerns ascertain the invasive nature and potentially health risks of the current women infertility therapy methods [15, 48]. Current female infertility treatment includes intrauterine insemination, ovarian drilling and ovulation induction for IVF/ICSI, which carries the ethical and financial dilemma of the implantation of too many blastocysts posts IVF/ICSI, leading to potentially undesired multiple pregnancies and births [39].
Current treatment technology rely on functional gamete production and haploid gamete function from both partners. If a partner can’t produce a functional gamete (oocyte or spermatozoon), therapeutic option is gamete donation [59]. Numerous factors negatively affect patients’ gamete function such as genetic factors in primary ovarian insufficiency in females and failure to produce spermatozoa through spermatogenesis in males, exposure to environmental and industrial toxicants, chemotherapies, and immune suppressant treatments [74].
Classifications of various stem cells characteristics according to which stem-cell based the therapies are performed, reconstructed from [15]
Classifications of various stem cells characteristics according to which stem-cell based the therapies are performed, reconstructed from [15]
Previously, medical professionals focused on treating cancers in prepubescent boys and girls to extend their lives with little concern on attentive patients’ fertility. As cancer survival rates in adolescents have raised, the focus has shifted to preserving fertility. One of the caring goals of the Oncofertility Consortium is to preserve fertility in patients undergoing rigorous cancer treatments such as high dose chemotherapies [11]. However, fertility preservation can be extended to any medical treatment that impacts fertility, such as immune suppressant treatment which has been shown to cause permanent sterility in some male patients [13, 31]. Cryopreservation of oocytes or ovarian tissue is an option for female patients prior to medical treatment, oocytes can be later utilized for IVF [10, 28]. For adult male patients, cryopreservation of a sperm sample is the least invasive procedure, but for prepubescent patients or those who unable to provide a sperm sample, cryopreservation of testicular tissue has become the latest innovation to preserve a patient’s fertility [32, 36, 56]. Several studies in mouse models have shown the ability to reintroduce recovered spermatogonial stem cells (SSC) from testis biopsies to restore fertility in sterilized mice [65]. Recently, this work has been extended to non-human primates and showed that isolated rhesus macaque SSC obtained from biopsies taken prior to busulphan-mediated chemosterilization can be retransplanted back into the testis to recolonize the testes, resulting in the production of functional spermatozoa [12].
This outstanding breakthrough now paves the way for reintroducing isolated human SSC from adolescent male patient’s tissue biopsies prior to high dose chemotherapy [12]. Of course, using the recent method cancer cells will be reintroduced in the patient after chemotherapy [23, 24]. While improvements have been made to separate SSC from cancer cells, the risk must still be considered [24]. Furthermore, SSC recolonization requires the intact somatic environment of the testes after the medical intervention. SSC expansion and differentiation are inhibited in patients with damaged or genetic defects in the somatic environment, which is ineffective at curing a patient’s infertility [42].
Application and derivation of patient-specific induced pluripotent stem cells (iPSCs) in male infertility: Diverse kinds of somatic cells derived from patients with idiopathic infertility are reprogrammed into iPSCs which latter are differentiated into male germ cells by numerous techniques. In case vital, iPSCs with known genetic deficiency could be corrected by genome editing technology. These cells could be utilized for regeneration studies, cell-based therapy, and in vitro disease modeling. In disease modeling, comparison between patients and normal derived cells, potentially gives novel signs to the fundamental methods for idiopathic male infertility that could additionally lead to the advancement of therapeutic plans.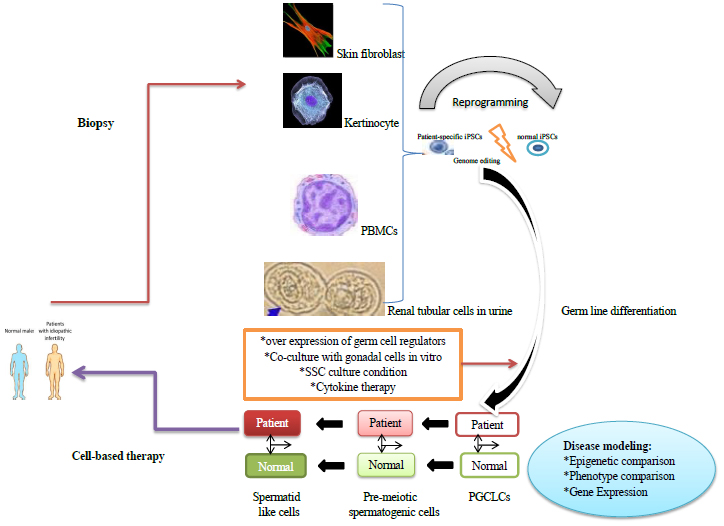
Recently, several independent groups evidenced the ability of human, non-human primate and mouse pluripotent stem cells to differentiate into primordial germ cells [66], precursor cells that contribute to gametogenesis in both males and females (Table 1). Studies with mouse embryonic stem cells have even shown the ability of the cells to make functional spermatozoa [45]. The recent work by Hayashi et al. demonstrates that stem cells can be differentiated into primordial germ cell-like cells and then transplanted into a sterile mouse testis for re-colonization and the generation of functional haploid sperm cells. Primordial germ cells have shown limited capacity to recolonize sterile tests in rodents [21].
This technique has not been fully examined in other mammalian models, including non-human primates. Thus, the possibility exists that pluripotent stem cells can be differentiated into a germ cell lineage more suitable for re-colonization and restoration of spermatogenesis. Human induced pluripotent stem cells (hiPSCs) might be made from in vitro derivation of functional germ cells from patient and patients’ somatic cells specific iPSCs could give novel therapeutic technique for infertile couples [19]. In fact, the current study group has recently reported that human embryonic stem cells (ESC) and induced pluripotent stem cells (iPSC) can be differentiated into SSC-like cells [14], capable of recolonizing the testis in several animal models. The ability to generate GMP (good manufacturing practice) human iPSC for potential clinical uses [26]; it is now possible to generate iPSC from patients sterilized by medical interventions, environmental or industrial toxicants or injury.
Combining this protocol with that of Hermann and colleagues introduces the first stem cell replacement therapy for male infertility, whereby iPSC differentiation into SSC-like cells followed by transplantation of the late cells into the patient’s testes restores fertility. To limit a step for stem cell replacement therapy is the testis somatic environment.
In case of damaged somatic environment not receptive to SSC transplant, the patient’s fertility would not be restored, and the only possible solution would be complete in-vitro differentiation into functional spermatids [14].
Herman study group [23, 24] demonstrated that human ESC and human iPSC can be differentiated in vitro into advanced spermatogenic stages, including round spermatids. While round spermatids do not fertilize oocytes in higher-order mammals, the results indicate that it is at least feasible to differentiate pluripotent stem cells into haploid spermatids.
Murine studies have shown fundamental intuition into the advancement of male germ cells from iPSCs both in vivo and in vitro [6, 33, 72]. Zhou et al. indicated the generation of haploid male gametes from mESCs that may provide feasible and fertile descendants [73]. Eminently PGCLCs derived from various mouse iPS cell lines shown diverse proficiency for spermatogenesis in vivo and a few of the descendant died prematurely [22].
Human iPSCs with azoospermia factor deletions created essentially less germ cell-like cells in vivo with a particular deficiency in gene expression. The results demonstrate that xeno-transplantation of hiPSCs directs germ cell differentiation in a way dependent on donor genetic history [51]. Theoretically, iPSCs from male infertility patients with genetic deficiency may be genetically corrected, differentiated into spermatogonia or PGCLCs in vitro, and after that transplanted back into the patient’s seminiferous tubules for therapeutic aim.
Hence, xeno-transplantation of iPSCs may also serve as tools for genetic study of human germ cell improvement in vivo (Fig. 1) [19].
The Improved differentiation strategy could lead to the maturation of round spermatids into elongated ones, capable of fertilizing an oocyte in IVF clinics. Future treatment manners for infertility/sterility could target in-vitro differentiation into functional spermatids and thus not necessitate testis cell transplantation [15].
Characterization of human male germ line cells
Primordial germ cells (PGCs) are originator cells of the germ line and are specified throughout early embryonic development in mammals. Mouse PGC (mPGC) Characterization has been considered broadly, which prepared a significant model for mammalian improvement. Briefly, mPGC specification is started by WNT signals and bone morphogenetic protein (BMP) from extra-embryonic tissues that stimulate the expression of PGC fate regulator genes in a small number of germ line cells of the primary post-implantation embryo [67].
The mPGCs at the base of the allantois (the fetal membrane lying below the chorion) start emigrate and colonize the genital ridge, along with genome-wide epigenetic reprogramming to remove imprints and other somatic epigenetic memories [62]. Post-migration PGCs began to sex-specific improvement and the male germ cells are under the influence on mitotic arrest, showing the end of the PGC level of germ line improvement.
Spermatogonial stem cells (SSCs) have remarkable usage in both regenerative and reproductive medicine. However, primary human SSCs are very rare, and a human SSC line has not yet been available.
In-vitro germ cell models as novel cures for infertility
Stem cell utility in the purpose of male infertility therapy has focused on those patients rendered sterile by exposure rather than a genetic component. In-vitro differentiation may alleviate some infertile patients with genetic causes such as those who experience defects in the somatic environment or possibly affected by the sertoli cell only syndrome [26, 44], but may not be useful for those with spermatogenic defects such as DAZ family deletion carriers or men with Klinefelter syndrome. However, development of in-vitro spermatogenic models is critical for understanding and identifying causes of either known or idiopathic infertility cases. These models could then be adapted for high-throughput screening to identify novel compounds capable of restoring spermatogenesis in vitro to be able to generate functional haploid spermatids or spermatozoa for fertilizing a partner’s oocyte in IVF clinics. This study highlighted the ability to differentiate human ESC and iPSC into various cell lineages found in spermatogenesis, including SSC, premeiotic spermatocytes, post-meiotic spermatocytes and round spermatids, although it has not yet shown whether individual cells track through all stages of spermatogenesis [14].
Stem cell therapies for infertile women
Until recent years, all attempts targeted to differentiate male haploid germ cells in to male stem cells. Investigators are now trying to generate haploid female germ cells from human pluripotent stem cells, an effort that has been failing so far, and could not produce even a similar cell to an oocyte with a functional ooplasm capable of fertilization [16].
However, the recent work by Hayashi et al. showed that mouse stem cells could be differentiated into oocyte-like cells capable of being fertilized by spermatozoa and generating normal progeny. This outstanding advancement further shows the ability of pluripotent stem cells to differentiate into all cells of the adult organism. Whether the work by Hayashi and colleagues can be adapted for human stem cells remains to be seen, but this advancement is a critical step forward in generating functional de-novo oocytes from human iPSC isolated from female patients rendered sterile by medical interventions, exposure to toxicants or by premature ovarian failure [21].
Mutations in mitochondrial DNA (mtDNA), inherited maternally, have been linked to severe human disorders, including myopathies, neurodegenerative diseases, diabetes, cancer and even infertility [8]. If severe enough, these mtDNA mutations are capable of preventing a woman from producing an offspring with her partner [53]. Tachibana et al. and Wolf et al. showed that using a non-human primate model, that mtDNA defects can be circumvented by spindle-chromosomal complex transfer from a mature metaphase-II oocyte into an enucleated mature donor oocyte. These oocytes are capable of being fertilized and giving rise to offspring that lack the deleterious mtDNA mutation, but maintain the maternal genomic DNA signature [60, 69]. This novel approach, while currently not confirmed in human oocytes, has the potential to remove deleterious mtDNA mutations contributing to infertility and would enable affected women to produce healthy offspring with their partner.
Recently, the work by White et al. [68] has identified a rare population of mitotically active germ cells in human ovaries that can be purified and cultured in vitro to spontaneously form oocytes. This work highlights a unique potential to generate oocytes in vitro from isolated cells in reproductive-aged women who may have a depleted follicle pool from genetic defects like Fragile X-associated primary ovarian insufficiency.
This recent advance, along with those described above, highlight the unique methodologies being developed to combat female-factor infertility [68, 70].
Epigenetics in assisted reproductive technologies
The DNA of spermatozoa is differentially methylated at several maternal and paternal imprinting regions, as well as exhibits unique global methylation patterns. Reprogramming of the epigenome and imprinted loci during gametogenesis and peri-implantation stages is very crucial for maintaining a proper pattern of inheritance, particularly at imprinted loci. There is a concern that assisted reproductive technologies such as intracytoplasmic sperm injection (ICSI) and round spermatid injection (ROSI) may increase the incidence of imprinting disorders and adversely affect embryonic development by using immature spermatozoa that may not have proper imprints or global methylation established. Deregulation of imprinted regions has been associated with the onset of Angelman syndrome in the cases undergoing intracytoplasmic sperm injection [5, 50]. Deregulation of Igf2 imprinted loci has been previously associated with malformed offspring in mice, characterized by retardation and Beckwith-Wiedemann syndrome [41]. However, long term studies on imprinting disorders in assisted reproductive techniques have denied an association between the two. The evidence describing ART procedures as increasing the frequency of imprinting disorders such as Prader-Willi Syndrome, Beckwith-Wiedemann Syndrome and Angelman Syndrome has remained contrasting [17, 64]. Odom et al., Eroglu et al. contend that use of ART does not increase the risk of imprinting disorders [17, 46], while Manipalviratn et al., express opposed the idea [38]. Large longitudinal studies must be conducted to examine this connection more thoroughly. Evidence has been provided, however, that ROSI is linked to abnormal zygotic epigenetic regulation [50].
Indeed, a study by Rajender et al. [50] and Tanaka et al. [61] demonstrated that injection with round spermatids versus mature spermatozoon results in distinguishable methylation patterns of the paternal zygotic genome. Data from this study show that zygotic genomes derived from round spermatids are remethylated after initial demethylation before completion of the first mitosis.
This inability to prevent global DNA remethylation does indeed lead to abnormal genome-wide DNA methylation in the paternal zygotic genome [50, 61]. Moreover, unlike spermatozoa, round spermatids exhibit H3K9 trimethylation, which is preserved through the first mitosis of the zygote? The authors believe that the differences in epigenetic patterns may account for the lower success rates of ROSI [50].
The novel innovation by Yamanaka and others of reprogramming adult somatic cells into embryonic stem-like cells has revolutionized patient-specific stem cell therapies in medicine, especially as GMP protocols for deriving iPSC are being established. Recent advances have shown the ‘promiscuity’ of stem cells to differentiate not only into somatic lineages but also into gamete lineages [54]. The ability to differentiate a patient’s iPSC into functional haploid products is an important step not only for providing material suitable for IVF, but also for developing a model system for chemical screens to identify novel compounds capable of curing a patient’s infertility. The generations of functional haploid products from patient-specific stem cells are a noble quest, but one that needs to be rigorously examined in non-human primate models before being utilized in a clinical trial. Long-term studies will need to be conducted to examine whether healthy offspring can be generated from a pluripotent stem cell-derived gametes. The best short-term uses of human research will be to develop in-vitro models for spermatogenesis and oogenesis for use with drug screens evaluating whether new medical treatments impact gametogenesis or to design novel contraceptives [15].
Conclusions
As against to the genome, that could be a constant unit over a period of time, the epigenome is incessantly altering, and it focuses on the effect that environment and external elements have on the genome. Epigenetic investigation has just lately achieved ground and novel exciting survey has led to an aggregation of scientific information about the gene expression regulation in biological homeostasis and establishment of techniques to artificially control gene expression [42]. Thus, a subsequent epigenetic study could contribute to the advancement of therapy for changes speci?cally associated with male gamete, as the spermatozoa. Epigenetic plan is exclusive and appropriate to fulfill the Requirements of this exceedingly specialized cell.
In addition, new information from epigenetic survey could be useful in realizing how male fertility could be altered by the stress or the environment and how to determine the harm precisely, which could be helpful in the protection and/or prevention against subsequent such damage. Epigenetics has lately achieved remarkable scientific consideration since it has added a new aspect to proteomic and genomic study, and holds significant promise in capturing the inconspicuous yet very momentous regulatory factors that would drive normal and abnormal sperm function. In spite of the fact that controversial, hiPSCs have huge potential for therapeutic and biological applications for male infertility. Recent advancement in generation of male germ cells from miPSCs not only hold remarkable promise for the foundation of in vitro human spermatogenesis patterns, but also give experiences into the human spermatogenesis regulation and mechanism of hPGC characterization. Based on these progressions, it is possible that hiPSCs would have more helpful suggestion for therapeutic implications for male infertility in combination with genome editing and EVs studies in the future. Nevertheless, the molecular function underlying human male germ cell improvements are still negligible understood. More comprehensive understanding of human germ cell improvement will be of considerable value for the use of hiPSCs in reproductive medicine and fundamental investigation.
Genetic changes contribute to a number of disorders either to a large or small extent; however similar knowledge about the contribution of epigenetics is being unraveled. The role of epigenetic modifications in other disorders must be revealed, which could explain the etiology of the disorders in several cases that genetics could not explain. Epigenetic changes have an advantage of being modified by certain drugs, thus understanding the process of epigenetic modification helps to treat epigenetic related disease completely. On contrast, genetic changes can’t be treated thus diagnosis of genetic changes is not helpful in curing disease. It is obvious that understanding epigenetic processes and their association with infertility would help us not only understand etiological factors but also treat male infertility.
Exploration of both genetic and epigenetic changes could go hand in hand, and this could help us explain the interaction between the two factors, if any.
Current development in the field of genetics and personal genomics have highlighted the potential diagnostic advantage of whole-genome evaluation for infrequent variants, epigenetic variations and copy number variations in identifying the reasons of male infertility. It is essential that researchers must also magnify the usage of these improvements with an improved focus on accurate and complete phenotyping of patients if this improvement ultimately is lead to improved diagnostic tests and treatment options.
The final execution of personalized medicinal drug based on the genomics revolution now happening is inside view; however will necessary precise analysis and consideration in both the clinic and the laboratory.
Footnotes
Acknowledgments
We would like to express our sincere thanks to Dr. Kyanoosh Malekzadeh from the Molecular Medicine Research Center, Hormozgan University of Medical Science, Bandar Abbas-Iran for his efforts in manuscript editing.
Conflict of interest
The authors report no conflicts of interest in this work.