
Abstract
BACKGROUND:
A potential relationship may exist between the vastus medialis (VM) and the contractile efficiency of the vastus intermedius (VI) due to their anatomical connection and common innervation by the same branches of the femoral nerve. However, this relationship has not been explored using physiological experiments.
OBJECTIVE:
This study aimed to use electromyography (EMG) to investigate the functional significance of the VI and VM muscles during isometric contraction.
METHODS:
Twenty-six healthy male volunteers underwent EMG recordings from the VI, VM, and vastus lateralis (VL) at 90∘ or 30∘ of knee flexion. The EMG pre-motor time (PMT) was measured during rapid knee extension in response to an auditory stimulus. Additionally, the M-wave latency was determined by femoral nerve stimulation and subtracted from the PMT to calculate the descending conduction time (DCT).
RESULTS:
DCT was shortest at VM, followed by VL and VI. The DCT difference of VI-VM was significantly greater than that of VI-VL during maximal effort at 90∘ of knee flexion.
CONCLUSIONS:
VM is neurally driven to act as a pre-activator that primes the VI muscle; this tendency is particularly pronounced when explosive high-moment knee extension is required.
Introduction
In the rehabilitation of patients with knee disorders, such as knee osteoarthritis (OA) and anterior cruciate ligament (ACL) injuries, training the quadriceps femoris muscle is essential to prevent disease progression, reduce symptoms, and facilitate a return to competitive activities [1, 2, 3, 4]. The specific training methods employed depend on various factors, such as symptom severity, disease stage, and type of surgical procedure. Nonetheless, it is essential to ensure adequate contraction of each muscle using palpation. Among the various muscles, therapists often focus on the contraction of the vastus medialis (VM), which may be because of the significant role of VM, particularly its distal part known as the VM obliquus (VMO), in counteracting lateral patellar deviation during quadriceps contraction induced by the Q angle [5, 6, 7, 8]. The patella functions as a pulley, effectively transmitting tension from the quadriceps muscles to the tibial tuberosity. Therefore, there is considerable attention given to the ability to predominantly contract the VMO, as it plays a pivotal role in patellar stability [9].
The VM is closely connected to the aponeurosis of vastus intermedius (VI) [10, 11], which is considered to be the most effective contributor to knee extension moment [5, 12, 13, 14]. In an old experiment performed by Lieb & Perry [5], an amputated femur was used to measure the tensile load on each quadriceps muscle as it was pulled by a weight during knee joint extension. They reported that VI exhibited the lowest tensile load. In addition, a fine-wire electromyography (EMG) study showed that VI made the highest contribution to exerting low-intensity moment [12]. Recent studies using ultrasound imaging have found muscle architectural changes to be associated with knee extension contraction [13, 14]. Grob et al. [11] reported that the VM is extensively connected to the aponeurosis of VI and exhibits a “clip-type double insertion” aspect, wherein the VM encloses the medial portion of the VI from the anterior and posterior sides. This anatomical feature of the VM enhances its contractile efficiency by tensing the VI. Furthermore, the VM and VI are innervated by the same medial branch of the femoral nerve [11]. The vastus lateralis (VL) is also associated with the VI [10, 15, 16, 17]. However, only morphological reports regarding the location and fusion extent between the aponeurosis have been found, without a functional anatomical description similar to the increased contractile efficiency observed for VI [10, 15, 16, 17]. While the anatomical relationships between the VM, VL, and VI have been reported, the physiological mechanisms through which the VM or VL enhance the contractile efficiency of the VI remain unclear. Investigating these mechanisms can provide a theoretical basis for quadriceps training.
Despite the deep location of the VI, Watanabe & Akima [18, 19, 20] demonstrated that surface electrodes placed on the distal lateral thigh can record EMG signals from the VI. Their method also provides insights into the muscle activity patterns of the VI during dynamic knee movements [21, 22, 23]. Using this technique for VI EMG recording, the functional relationships between the vasti muscles can be investigated using EMG. The hypotheses tested in this study are as follows: 1) the VM exhibits the earliest onset of muscle activity, employing a neural strategy to increase the VI tension as a form of pre-activation, and 2) the muscle activity of the VM shows a stronger correlation with that of the VI compared to that of the VL. Therefore, the aim of this study was to investigate the correlation between the difference in EMG reaction time, known as pre-motor time (PMT), and the onset of muscle activity and the muscle activity of the VM and VL in relation to the VI.
Materials and methods
Participants
To determine the required sample size for the study, a power analysis was conducted using G*power 3.1.9.4. The analysis used the following factors: “a priori: compute required sample size given
The participants were informed regarding the study purpose, procedures, and risks. Written informed consent was obtained from the participants before the study was conducted. The study was conducted after obtaining approval from the Research Ethics Review Committee of Kansai University of Health Sciences (approval no.: 21–30; approval date: March 18, 2022). The study was conducted in accordance with the 1964 Declaration of Helsinki and its later amendments.
Experimental setup. (a) Positions of the recording electrodes for the three vasti muscles and a reference electrode; (b) Experimental procedure. VI, vastus intermedius; VM, vastus medialis; VL, vastus lateralis; PMT, pre-motor time; ME, maximal effort; SME, submaximal effort.
EMG signals from the VI, VM, and VL were recorded using an MQ16 telemetry electromyograph (Marq-Medical, Farum, Denmark) and the VitalRecorder 2 waveform data collecting program (KISSEI COMTEC Co., Ltd., Nagano, Japan). The amplification was as follows: type, single differential; input impedance, 1 G
Electrical stimulation to evoke M-wave
A VikingQuest System version 9.0 stimulator (Natus Neurology Inc., Middleton, WI, USA) was used to evoke M-waves. The cathode of the stimulation electrode was a 20-mm-diameter self-adhesive disposable circular electrode (Natus Neurology Inc., Middleton, WI, USA), which was placed over the femoral nerve in Scarpa’s triangle, with reference to the pulsation of the femoral artery. For the anode, a 40
Knee extension moment measurement
Knee extension moment was measured using a Biodex System 3 dynamometer (Biodex Medical Systems Inc., Shirley, NY, USA) with the knee joint at 90∘ or 30∘ of flexion (0∘ of full extension). The participant was seated with the backrest adjusted to maintain 60∘ of hip flexion (0∘ being the mid-flexion-extension position). The moment signal was recorded by the Biodex System 3 at a sampling rate of 2 kHz (16 bit) using an AD converter (AIO-163202FX-USB, Contec Co., Ltd., Osaka, Japan) and was simultaneously transmitted to the VitalRecorder along with the EMG signals.
Experimental procedure
Prior to the experiment, participants were instructed to engage in a series of warm-up exercises, including pedaling on a bicycle ergometer for 10-min, followed by exercises to achieve maximum voluntary moment in knee extension using the Biodex System 3.
Following the warm-up exercises, three tasks were performed: 1) the ramp task, 2) PMT measurements, and 3) M-wave measurements, which were conducted at both 90∘ and 30∘ of knee joint flexion. The magnitude of extension moment at different knee joint angles is influenced by factors such as muscle length, moment arm, and the tilt angle of the patellar tendon [25, 26, 27]. However, it is believed that the bony stability of the patella particularly affects the muscle activity of the VM and VL [28]. Therefore, two knee joint angles were selected for the measurements: 90∘ of flexion, which provides high bony stability, and 30∘ of flexion, which is considered to have minimal impact on patellar stability. The order of the measurement angles was randomized for each participant, and a series of measurements were taken by changing the angle after completing the measurements at one angle. A rest period of 2–3-min was provided between angle changes, as deemed necessary based on the participant’s level of fatigue (Fig. 1b).
Knee extension ramp task
The participant was instructed to perform the knee extension ramp task three times with the goal of achieving maximum voluntary isometric contraction (MVC) moment. This task involved exerting force gradually over a period of approximately 5-s. During the task, EMG signals were recorded from the VI, VM, and VL muscles. The participant received visual feedback of the moment curve displayed on a monitor positioned in front of them. The examiner provided encouragement to the participant, emphasizing the importance of exerting smooth and maximal moment throughout the task.
PMT measurement
To record the PMT, EMG signals were recorded from the VI, VM, and VL while the participant performed knee extension as rapidly as possible, cued by an auditory stimulus emitted from a speaker positioned behind the participant. The auditory stimulus signals and a trigger signal to synchronize the EMG and moment signals were recorded using VitalRecorder 2.
The auditory stimulus served as a cue for the participants to initiate the knee extension movement. The intensity of the knee extension moment was assessed in two ways: application of maximal effort (ME) as quickly as possible and submaximal effort (SME) at 50% of the maximal voluntary contraction (MVC) moment. The target moment line for the SME was calculated based on the MVC moment obtained from the ramp task and displayed on the monitor. Two types of tests were conducted to examine how the vasti muscles respond when instantaneously generating muscle power or controlling muscle power to some degree. In each test, the participants performed a total of 20 knee joint extensions, with 10 repetitions at each intensity. To ensure equal conditions among the participants, the ME test was performed first, followed by the SME test, and then alternated. During the measurements, the examiner provided encouragement to the participants to exert moment as rapidly as possible. PMT measurements were conducted by allowing the participants to relax for intervals of at least 10–15-s while monitoring for the absence of EMG activity in the VI, VM, and VL muscles.
M-wave recording
After the PMT measurement, the maximum M-wave was recorded three times from the VI, VM, and VL while the participant was in a relaxed state. During this time, the same investigator (Y.T.) manually positioned the cathode of the stimulation electrode as close as possible to the participant’s femoral nerve and applied supramaximal stimulation.
Data analysis
EMG, moment, auditory stimulus, and synchronization signals obtained from the VitalRecorder 2 waveform data collecting program were imported into the electromyogram analysis program BIMUTAS®-Video (KISSEI COMTEC Co., Ltd., Nagano, Japan). The data were analyzed after synchronization processing. The moment curves were smoothed using a 6-Hz low-pass filter before analysis. The following analyses were performed by a single investigator (Y.T.).
PMT analysis. (a) EMG of the three vasti muscles, auditory stimulus, and knee extension moment. (b) Extended view showing the EMG of the three vasti muscles and auditory stimulus. Arrows indicate the onset of auditory stimulus and EMG. VI, vastus intermedius; VM, vastus medialis; VL, vastus lateralis; PMT, pre-motor time; EMG, electromyography.
The sweep rate of the EMG analysis screen was set to 0.05 s/division. After processing the raw EMG signals into full-wave rectified waveforms, the investigator visually checked the EMG onset of each muscle. Furthermore, the time from the auditory stimulus signal was measured. The mean and standard deviation of the PMTs performed in ME and SME for each muscle were calculated. Similarly, the mean, standard deviation, and coefficient of variation of the knee extension moment were also calculated.
Measurement of M-wave latency. The dashed line indicates the onset of the stimulus artifact and the arrows indicate the M-wave onset. VI, vastus intermedius; VM, vastus medialis; VL, vastus lateralis.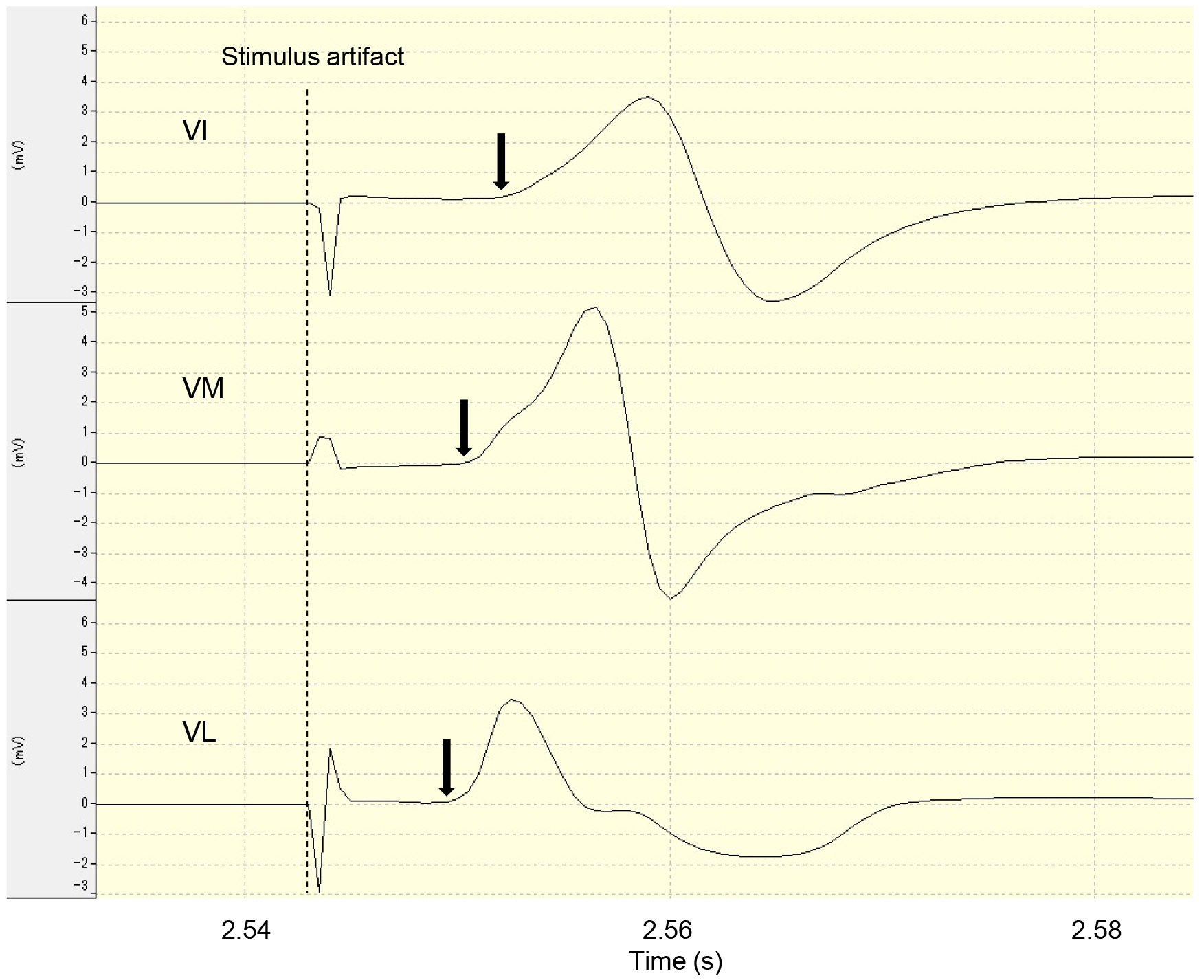
The M-wave latency was measured by setting the sweep rate of the EMG analysis screen to 0.02 s/division. The measurement involved determining the time interval from the baseline to the rise of the waveform from the electrical stimulation artifact. The average latency from three measurements was recorded.
The PMT refers to the duration taken by the central nervous system to process information in response to stimulation and to transmit instructions to the effector for executing the appropriate movement [29]. Because the PMT of VI, VM, and VL is influenced by physical factors, such as the variation in distance to the electrode attachment position, the latency of each M-wave induced by femoral nerve stimulation was measured and subtracted from the PMT to calculate the descending conduction time (DCT) from the central nervous system to the inguinal femoral nerve, which reflects voluntary contraction.
PMT and M-wave latency (ms) of VI, VM, and VL
PMT and M-wave latency (ms) of VI, VM, and VL
Each value represents the mean
Measurement of EMG root mean square during ramp task. The EMG root mean square was calculated for the gray zones. VI, vastus intermedius; VM, vastus medialis; VL, vastus lateralis; EMG, electromyography; MVC: maximal voluntary contraction.
In the ramp task, the moment curve that exhibited the smoothest and highest moment exertion was selected as the reference trial. For this trial, the root mean square (RMS) values of EMG signals were determined for a duration of 100 ms before and after the 20%, 40%, 60%, and 80% points of MVC time. This resulted in a total duration of 200 ms. Each RMS value was then normalized as a percentage relative to the RMS value obtained during the 200 ms period near the peak moment development.
Statistical analysis
To analyze the time difference from VI, the DCTs from VM and VL (VI-VM and VI-VL) were compared. Because the results of the Shapiro-Wilk test showed normal distribution of the ME and SME data at 90∘ flexion, Student’s
As the Shapiro-Wilk test showed that the coefficient of moment variation during DCT measurements was distributed normally, the Student’s
Because the RMS values for VI, VM, and VL obtained during the ramp task did not satisfy the normality assumption, as indicated by the Shapiro-Wilk test results for both the 90∘ and 30∘ knee flexion positions, Spearman’s rank correlation coefficient was used for correlation analysis between VI and VM, VI and VL, and VM and VL.
IBM SPSS Statistics (version 28; IBM Inc., Armonk, NY, USA) was used for statistical analyses. The significance level was set at 5%.
Results
DCT
PMT and M-wave latency (Table 1)
The mean PMT values were lowest for VM at 90∘ and 30∘ knee flexion and for both ME and SME, followed by VL and VI. Although the M-wave latency was shorter for VL than for VM, the opposite was true for PMT.
Descending conduction time (ms) of VI, VM, and VL
Descending conduction time (ms) of VI, VM, and VL
Each value represents the mean
Knee extension moment during PMT
Each value represents the mean
The mean DCT, calculated by subtracting the M-wave latency from the PMT, was found to be the shortest for VM at 90∘ and 30∘ knee flexion, as well as for both ME and SME, followed by VL and VI. In the 90∘ flexion position, the DCT tended to be faster in ME compared to SME. However, the opposite trend was observed in the 30∘ flexion position, with faster DCT in SME compared to ME.
In the 90∘ flexion position, when comparing the DCT difference with VM or VL active prior to VI, ME exhibited a significantly larger DCT difference with VM compared to VL, with a larger effect size (VI-VM: 8.46
In the 90∘ flexion position, the number of participants with VI preceding VM, which would affect the mean DCT difference, was 1 (3.8%) in ME and 2 (7.7%) in SME. By comparison, in the 30∘ flexion position, the number of participants with VI preceding VM was 4 (15.4%) in ME and 5 (19.2%) in SME. On the other hand, VI preceded VL in 7 (26.9%) participants in ME and 6 (23.1%) participants in SME at 90∘ flexion and in 8 (30.8%) participants in ME and 10 (38.5%) participants in SME at 30∘ flexion.
Knee extension moment during PMT measurement (Table 3)
The knee extension moment for the ME and SME at each knee joint angle were 91.00
Correlation coefficient (
) of EMG RMS
Correlation coefficient (
*p< 0.001. EMG, electromyography; RMS, root mean square; VI, vastus intermedius; VM, vastus medialis; VL, vastus lateralis.
The EMG RMS values of VI showed a significant positive correlation with both VM and VL (
Discussion
Sequence of neural drive to three vasti muscles
Surface electrodes can be used to record EMG signals [18, 19, 20, 21, 22, 23, 24, 30], even in the case of the deep-seated VI within the quadriceps muscle. In this study, the EMG recording site for VI was identified on the distal lateral thigh using sonographic imaging. EMG recordings from this site have demonstrated minimal crosstalk from the nearby VL muscle [18]. Furthermore, a strong correlation was observed between the muscle activity dynamics of the proximal muscle belly and EMG recordings obtained from distal localized sites [20]. While efforts were made to align the VM electrode as near to the VI as possible along the femoral long axis, it was not possible to align the conduction distance to the muscle in all three dimensions. In addition, the VL electrode had to be positioned more proximally than the VI electrode. Therefore, the PMT incorporated the above-mentioned elements of varying impulse conduction distances. Therefore, in the present study, the maximum M-wave latency recorded from the same electrode during stimulation of the femoral nerve in the Scarpa’s triangle was subtracted from the PMT to ascertain the sequence of neural drive to the vasti muscles through the information processing mechanism of the central nervous system, as indicated by a difference in DCT. The results demonstrated that the DCT of VM was consistently shorter at all knee joint angles and contraction intensities, followed by VL and VI. This trend was also observed in PMT, with VL exhibiting the shortest M-wave latency, followed by VM and VI, suggesting that the physical positioning of the recording electrodes was related to their proximity to the site of electrical stimulation. Interestingly, during voluntary contraction, the PMT and DCT of the distal part of VM were shorter than those of the proximal VL located in the thigh. These results are similar to those of a previous study that measured PMT using light stimulation, which also found a shorter PMT in the VM despite the more distal placement of the electrode compared to the VL [29]. This implies that the motor unit recruitment is regulated by the central nervous system [31], resulting in variations in the arrival of impulses associated with the onset of muscle activity in VM and VI, despite receiving their nerve supply from the medial branch of the same femoral nerve [11]. According to Thiranagama [32], the femoral nerve supply to the VM is divided into lateral and medial branches. The lateral branches originate from the 3rd and 4th lumbar medulla, supplying the proximal VM and VI, whereas the medial branches originate from the 2nd and 3rd lumbar medulla (some from the 1st lumbar medulla), supplying the distal VM [32]. On the other hand, it has been suggested that the nerve supply pattern of VL overlaps with that of VI [17], with branches from the nerve trunks supplying the VI mainly involved in the nerve supply of the VL, extending more distally than the EMG recording sites of the VL in this study [15]. Although the specific medullary segments of VI and VL receiving innervation are unknown, it is possible that the PMT and DCT of VM recorded in this study were shorter due to the descending impulses from the corticospinal tract reaching the upper lumbar spinal cord, innervating the distal VM first, while impulses from alpha motor neurons travel linearly through the medial branch to the muscle. The degree of VM pre-activation was influenced by the differences in knee joint angle, followed by the manner in which the knee extension moment was applied.
Knee angle specificity
At 30∘ of knee flexion, the VI muscle length was shortened compared to 90∘ of knee flexion, resulting in decreased muscle tension. Therefore, we hypothesized that the VM, which is closely connected to the VI, would contract earlier to increase the tension in the VI, particularly at 30∘ of knee flexion. Furthermore, we anticipated that achieving bony stability of the patella would be challenging at 30∘ of knee flexion, considering the contact between the patella and the femoral intercondylar groove. Therefore, we expected to observe a significant difference in DCT between VI-VM and VI-VL at 30∘ of knee flexion, which could contribute to medial patellar stability. However, unexpectedly, significant results were observed at 90∘ of knee flexion.
The contribution of VM to knee extension moment has been investigated through electrical muscle stimulation (EMS) and femoral nerve stimulation-induced knee extension moment. Selective fatigue of the VM using EMS resulted in significant reduction in femoral nerve stimulation-induced knee extension torque (triplet, twitch). However, no significant difference was found in the rate of moment reduction across different knee joint angles (10–90∘) [33]. Furthermore, even after selectively fatiguing the VMO, which was involved in medial patellar stability, with EMS, there was no significant difference in the rate of decrease in knee extension torque (twitch) between 90∘ and 30∘ of knee flexion [34]. These findings indicate that, in healthy individuals, the VM contributes to knee extension moment across all knee joint angles, and there is no specific angle that exclusively provides patellar stability. Thus, there is no evidence of the need of VM pre-activation for medial patellar stability prior to the VI during knee extension moment exertion at 30∘ of knee flexion compared to 90∘ of knee flexion. However, notably, despite the fact that 90∘ of knee joint flexion is less favorable for moment application than 30∘ of knee flexion due to the reduced moment arm and patellar tendon tilt angle in the sagittal plane [26, 27], the knee extension torque was significantly greater at 90∘ of knee flexion [35] (Table 3). This difference in moment can be attributed to the increase in muscle length. It is plausible that the condition requiring a higher moment to be generated more rapidly emphasizes VM pre-activation to enhance the contractile efficiency of the VI.
Manner of moment exertion
Even in the 90∘ of knee flexion position, we did not observe significant differences in DCT between VI-VM and VI-VL during SME compared to ME. This lack of significance may be attributed to the magnitude of moment exertion.
During isometric knee extension, the median power frequency of the VL muscle increases linearly with the increasing contraction force, while the VM muscle shows minimal change from 50% of the MVC [36, 37]. This suggests that, when considering the pure contraction intensity, ME may not differ significantly from VI-VM, as the greater difference in VI-VL is primarily due to the faster muscle fiber conduction velocity in the VL. However, the present study focuses on the power component, such as the ability to rapidly generate knee extension moment in response to a sound stimulus. As the power demand of the motor task increases, higher threshold motor units are recruited [38]. Therefore, it is plausible that, in this study, where both ME and SME tasks required rapid knee extension moment generation, relatively high threshold motor units were readily mobilized in both cases, resulting in a less pronounced difference in DCT. Furthermore, SME also involves adaptation to a target moment profile. Evidence suggests that the coefficient of variation in moment exertion was significantly greater for SME than for ME, with significant difference between them. This suggests that VM pre-activation is advantageous in situations where a large knee extension moment needs to be generated explosively. However, further detailed studies are warranted in the future.
Finally, although on average the VI muscle exhibited the slowest DCT across all joint angles and contraction intensities, there were participants in whom the VI had a faster DCT than the VM and VL muscles. It can be inferred that achieving the desired motor goal does not necessarily require differences in the timing of muscle activity initiation between the VI and VL. Despite their superficial or deep placement within the anterolateral thigh, both VL and VI are involved in exerting knee joint extension moment. In contrast, VM provides medial stability to the patella through its contraction vector. The “clip-type double insertion” between VM and VI provides faster neural drive to the VM, ensuring tension in the VI during pre-activation.
Recruitment patterns in the VI and other vasti muscles
The VI muscle, which plays a key role in generating knee extension moment, is located deep within the quadriceps muscle, making it challenging to assess its contraction strength through visual and palpation methods commonly used in rehabilitation settings. Therefore, we investigated the extent to which the VI is related to the contraction of the superficially located VM and VL muscles and explored the possibility of monitoring VI activity based on the contraction patterns of the superficial muscles.
The correlation coefficient between VI and VM and VL was 0.85 in the 90∘ knee flexion position and 0.88 in the 30∘ knee flexion position. Similarly, the correlation coefficient between VM and VL at the surface layer was above 0.9. Watanabe and Akima [19] examined the relationship between normalized EMG RMS and normalized torque in a 90∘ knee flexion position, similar to the present study, and reported lower normalized EMG RMS in VI compared to VL by up to 20–40% of MVC. Although a large deviation was observed in VI, no significant difference was observed in the deviation from normalized torque. These results suggest that the neural control of VI differs from that of other vasti muscles. Therefore, the correlation coefficients for VI-VM and VI-VL were slightly lower than those for VM-VL. Saito and Akima [30] further investigated the knee joint at 60∘, 30∘, and 90∘ of flexion, and found that the normalized EMG RMS of VI was significantly lower than that of VM and VL at 30∘ of knee flexion. They attributed the lower normalized EMG RMS of VI compared to that of VM and VL to the limb position with the shortest muscle length. Based on previous reports, we anticipated that the correlation coefficients between the 90∘ and 30∘ knee flexion positions would be different in our study. One possible factor that may have influenced these differences in results is that our study used a ramp task. In addition, the VL electrode was placed distally to account for the M-wave recording, which should be evaluated in future research. However, considering that the VM contributes to muscle coordination and provides medial patellar stability to the VI, monitoring the VM muscle contraction is crucial during rehabilitation interventions.
Study limitations and clinical implications
In this study, we used EMG to investigate the functional anatomy hypothesis proposed by Grob et al. [11], which posits that the VM contraction increases the efficiency of the VI contraction. The EMG signals from the VI were recorded from the distolateral part of the thigh, similar to previous reports [18, 19, 20, 21, 22, 23, 24, 30]. This part corresponds to the lateral part of the VI and differs from its medial part, where a “clip-type double insertion” of the VM is observed. Therefore, the muscle activity of the lateral part of the VI differs from that of the medial part and cannot be tested completely based on the functional anatomical hypothesis proposed by Grob et al. [11].
A musculoskeletal model simulation demonstrated that delaying the onset of VM muscle activity by 5 ms relative to the VL during lower limb ground contact during running increased the lateral compressive load on the patellofemoral joint, while preceding it by 5 ms decreased the load [39]. Although a direct comparison between the previous and present studies is not possible, the results suggest that the initiation of the VM muscle activity preceding the VI by more than 5 ms may significantly contribute to increasing the VI tension. These findings could have clinical significance under the hypothesis that it increases the efficiency of VI contraction.
The relationship between the VM and VI muscles extends beyond enhancing the efficiency of knee extension moment exertion. It also plays a crucial role in the static support mechanism for medial patellar stability. The VMO and VI are involved in communicating with the medial patellofemoral ligament, which is a critical component of the static support mechanism and forms a part of both tendinous membranes [40, 41]. The focus should be on strengthening the VM in relation to the VI, as it plays a crucial role in maintaining patellar stability in conjunction with both muscles. Furthermore, the function of the VM has received considerable attention among patients with knee OA. Studies have found that factors that cause muscle weakness, such as decreased muscle cross-sectional area and accumulation of non-contractile elements (e.g., intramuscular fat), are associated with OA progression and exacerbation of symptoms [42, 43, 44]. If VM muscle weakness impairs the moment exertion efficiency of the VI, it can hinder daily activities, such as standing up, walking, and climbing stairs. Impairment in these activities can contribute to further advancement of knee OA. Therefore, strengthening of the quadriceps muscles, with particular emphasis on the VM muscle, is crucial during the rehabilitation of patients with knee OA.
Conclusion
VM is neurally modulated to act as a pre-activator for efficient contraction of the VI, which is particularly evident when a rapid and large knee extension moment is exerted. This study reaffirms the importance of palpating the superficial VM muscle contraction to determine the muscle activity dynamics of the deeply located VI during knee rehabilitation, as well as the significance of strengthening the VM to stabilize the knee joint and exert a large knee extension moment.
Author contribution
CONCEPTUALIZATION: Yoshitsugu Tanino.
METHODOLOGY: Yoshitsugu Tanino, Wataru Yamazaki, Yuki Fukumoto, Takaki Yoshida, Tetsuya Nakao, Toshiaki Suzuki.
INVESTIGATION: Yoshitsugu Tanino, Wataru Yamazaki, Yuki Fukumoto.
FORMAL ANALYSIS: Yoshitsugu Tanino, Yuki Fukumoto.
WRITING – ORIGINAL DRAFT PREPARATION: Yoshitsugu Tanino.
WRITING – REVIEW & EDITING: Yoshitsugu Tanino, Wataru Yamazaki, Yuki Fukumoto, Takaki Yoshida, Tetsuya Nakao, Toshiaki Suzuki.
All authors have read and agreed to the published version of the manuscript.
Ethical considerations
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards. The study was approved by the Research Ethics Review Committee of Kansai University of Health Sciences (approval no.: 21–30; approval date: March 18, 2022). Informed consent was obtained from all individual participants included in the study.
Funding
The authors report no funding.
Footnotes
Acknowledgments
The authors have no acknowledgments.
Conflict of interest
The authors declare that they have no conflict of interest.