
Abstract
Neurodegenerative brain changes can affect the functional connectivity strength between nodes of the default-mode network (DMN), which may underlie changes in cognitive performance. It remains unclear how the functional connectivity strength of DMN nodes differs from healthy to pathological aging and whether these changes are cognitively relevant. We used resting-state functional magnetic resonance imaging to investigate the functional connectivity strength across five DMN nodes in 25 healthy controls (HC), 28 subjective cognitive decline (SCD) participants, and 25 prodromal Alzheimer’s disease (AD) patients. After identifying the ventral medial prefrontal cortex (vmPFC), posterior cingulate cortex (PCC), retrosplenial cortex (RSC), inferior parietal lobule, and the hippocampus we investigated the functional strength between DMN nodes using temporal network modeling. Functional coupling of the vmPFC and PCC in prodromal AD patients was disrupted. This vmPFC-PCC coupling correlated positively with memory performance in prodromal AD. Furthermore, the hippocampus de-coupled from posterior DMN nodes in SCD and prodromal AD patients. There was no coupling between the hippocampus and the anterior DMN. Additional mediation analyses indicated that the RSC enables communication between the hippocampus and DMN regions in HC but none of the other two groups. These results suggest an anterior-posterior disconnection and a hippocampal de-coupling from posterior DMN nodes with disease progression. Hippocampal de-coupling already occurring in SCD may provide valuable information for the development of a functional biomarker.
Introduction
The brain is organized into overlapping networks whose regions can dynamically interact and compete with each other in order to mediate and integrate cognitive functions [1, 2]. Cortical hubs are highly connected functionally specialized areas [3] and display heightened metabolism [4]. Importantly, regions vulnerable to Alzheimer’s disease (AD) pathology show high anatomical overlap with the hubs of the default mode network (DMN) [5]. The DMN is associated with episodic memory functions and has indeed consistently been shown to be affected in AD [4, 6]. Data therefore suggest that this high metabolic activity may make these cortical hubs particularly vulnerable to brain diseases, suchas AD [3].
Pathological changes in these cortical hubs will affect their functional connectivity [7] and, as such, influence cognitive performance [2]. While earlier studies report altered functional connectivity in AD patients, the precise pattern remains ambiguous, ranging from increased to decreased to no change in global connectivity and lower or no change in local connectivity in AD patients relative to healthy controls (HC) [8–15]. These studies focused on whole brain or predefined regional connectivity. However, studies examining exclusively the connectivity across DMN nodes [8, 17] consistently showed disrupted patterns in AD patients compared to HC, albeit with conflicting results for the integration of the posterior cingulate cortex (PCC). These inconsistencies may result from the heterogeneity in the disease stages of patients included in these studies or the anatomical definition of the PCC.
The PCC is a key hub among the DMN nodes as it is the only hub directly interacting with other DMN nodes [18]. Interestingly, the PCC is an anatomically and functionally heterogeneous region. We recently demonstrated that the retrosplenial cortex (RSC) can be functionally dissociated from the PCC [19], showing different functional connectivity patterns in prodromal AD patients compared to cognitively normal individuals. Furthermore, the PCC is hypometabolic and atrophic in the early stages of AD [20–23]. In addition to the PCC/RSC, the hippocampus, a structure crucial for memory formation [24, 25] and anatomically connected to the RSC, has consistently displayed pathological changes early on in the disease [26–28]. Despite its pre-eminent role in memory formation, there is a recent debate as to whether the hippocampus belongs to the DMN or is indirectly related to the DMN [29].
We hypothesize that the functional coupling between DMN nodes decreases with disease progression, reflecting progressive disintegration of the DMN. To investigate this, we included patients across various stages, ranging from subjective cognitive decline (SCD) to prodromal AD, which, to our knowledge, no study has done to this extent. Secondly, we sought to study the association between changes in functional connectivity strength of DMN nodes and memory performance. This is a critical issue in AD research as changes in the functional brain architecture can lead to cognitive deficits. Last, we speculated that the interaction with the hippocampus depends on the choice of DMN node [29, 30]. We thus took a complementary approach by also including the hippocampus. Together, our findings provide more insight into alterations in the connectivity strength of DMN nodes underlying the neurobiology of AD and its relationship to cognition, may eventually help to identify at-risk patients, and may thereby serve as potential groundwork for the development of new disease-modifying treatments including neurostimulation [31].
Materials and Methods
Participants
Initially, a grand total of 106 subjects (28 HC, 33 SCD participants, 45 prodromal AD patients) were recruited into our larger study. After careful pre-screening, we included 87 individuals (27 HC, 29 SCD participants, 31 prodromal AD patients) into the current study. HC were recruited from the local research facility, both SCD and prodromal AD patients from the outpatient Memory Clinic Cologne Jülich. All study participants were carefully screened to ensure inclusion criteria [e.g., for undergoing magnetic resonance imaging (MRI)] and rule out any neurological or psychiatric diseases or medications that affect cognitive functions (except for acetylcholinesterase inhibitors). All participants were native German speakers or sufficiently fluent in German.
To be included into the current study, HC were required to have normal scores on neuropsychological tests (corrected for age, gender, and education) with no cognitive complaints on the memory complaint questionnaire (MAC-Q) using a cut-off score of < 25 [32] and no symptoms of depression as assessed using the Hamilton depression rating scale [33] (cut-off <8) [34]. Inclusion criteria for SCD participants consisted of self-perceived memory complaints with a cut-off value of≥25 on the MAC-Q [32] but average scores on neuropsychological tests (corrected for age, gender, and education) [35]. For both, HC and SCD participants, average scores on standardized tests were defined as within 1.5 SD of the adjusted normative mean. If one test score was below 1.5 SD from normative scores, this individual was only included if i) the score was not 2 SD below the age norm, ii) all other sub-parameters and test scores fell within the average range, so that normal intraindividual variability could be assumed. Importantly, neuropsychological test results were evaluated by our team of multi-disciplinary clinicians prior to categorization as neuropsychological deficits can be the result of a variety of factors including early signs of dementia, but also sleeping problems, motivation, anxiety, depression, medication, or emotional state. Identification of SCD participants further included self-perceived memory decline compared to other cognitive functions and in reference to others of the same age occurring during the past five years as determined by the individuals’ medical history and an informant report, if available [35], at an age cut-off of 55. We additionally strived to exclude participants where other etiologies could explain self-perceived memory deficits including vascular, psychiatric, or other systematic etiologies by carefully evaluating laboratory results including blood samples, the patient’s medical history, and additional questionnaires. Our team of multi-disciplinary clinicians (HIJ, JK, öO, NR, BR) meticulously ensured the compliance of these criteria for every included SCD participants prior to classifying them as such. Prodromal AD patients had to fulfill the latest research criteria proposed by Albert et al. [36], Dubois et al. [37, 38], and Petersen [39]: self-reported memory complaints or informed by a knowledgeable caregiver (verified during the anamnesis or using the cut-off value of≥25 on the MAC-Q) [32] along with objective cognitive deficits (corrected for age, gender, and education) of at least 1.5 SD from the normative mean [39, 40], a mini mental status examination score of ≥23 [41], and, most important, evidence of biomarkers, i.e., cerebrospinal fluid tau/Aβ42 ratio of > 0.52 [42], medial temporal lobe atrophy score≥3 using MR imaging [43], or typical temporoparietal hypometabolism on fluorodeoxyglucose positron emission tomography (FDG-PET) imaging [36, 44], but preserved activities of daily living (confirmed by a clinician during the anamnesis or by the Bayer Activities of Daily Living Scale cut-off of > 5) [45]. This combination of evidence ensures the selection of patients that have the highest probability of converting to AD and are thus considered prodromal AD patients [37, 38]. A mild clinical depression (as per pathophysiology often a comorbid condition [46]) was not considered an exclusion criterion for both SCD and prodromal AD patients provided it was not the primary cause of the cognitive deficits, as determined by our multi-disciplinary team of clinicians. Depressive symptoms were assessed with the Hamilton depression rating scale using a cut-off score of < 17 [33] at the time of the study visit.
To ensure adherence to the inclusion criteria and accurate categorization of our three groups, all available data of each individual including laboratory results, cerebrospinal fluid values, neuroimaging data, the patients’ medical history, and additional questionnaires were carefully reviewed. Consequently, participants with structural abnormalities, including incidental findings such as cysts or vascular infarcts (n = 7) were excluded from the present study, as were two data sets due to technical problems. The final sample size thus consisted of 25 HC (15 male, 10 female), 28 SCD participants (13 male, 15 female), and 25 prodromal AD patients (15 male, 10 female). Within the prodromal AD group, we ensured the presence of at least one biomarker, that is, cerebrospinal fluid (n = 18), medial temporal lobe atrophy (n = 25), or typical FDG-PET temporoparietal hypometabolism (n = 14) (Supplementary Table 1).
This study was part of a larger study, which was approved by the local ethics committee, in accordance with the declaration of Helsinki [47], and performed after informed written consent of each participant. Healthy participants were reimbursed. In-patients were not reimbursed since imaging (FDG-PET) was part of their diagnostic procedures. We did, however, pay for and organize their traveling costs and lunch.
Neuropsychological assessment
Every participant underwent extensive neuropsychological testing. This included the mini mental status examination [41], verbal learning memory test (VLMT) [48] (a German equivalent to the Rey auditory verbal learning test [49]), logical memory, design memory and symbol span subtests of the Wechsler memory scale (WMS-IV) [50], digit span and digit symbol substitution subtests of the Wechsler adult intelligence scale (WAIS-III) [51], autobiographical memory subtest of the Cambridge examination for mental disorders of the elderly [52], Stroop test [53], Trail making test part A and B [54], mental rotation subtest of the “Leistungsprüfsystem” +50 (German intelligence test) [55], Money road map test [56], brief test of attention [57], and the Regensburger word fluency test [58]. As patients had their neuropsychological testing as part of the clinical routine at the Memory Clinic Cologne Jülich, test results were used for the current study if administered no more than three months prior to study participation. Otherwise, an identical test battery was repeated at the time of the study visit with alternative versions of the tests (when available). The mean time between the neuropsychological testing and MR scanning was 15.4±27.8 days. Due to mental exhaustion, some data were missing from one HC (for one test score), two SCD patients (for 12 test scores), and nine prodromal AD patients (for 30 test scores) (see Table 1 for details).
Sociodemographics, clinical characteristics, and neuropsychological scores for all study groups
Numbers are given as means (SD) unless stated otherwise. Scores reflect number of correct items unless stated otherwise. If significant group differences were found at p < 0.05 (ANOVA F-test) post-hoc pairwise t-tests were performed, FDR-corrected at p < 0.05. Asterisks next to the values indicate significant post-hoc differences at p < 0.05. *compared to HC, **compared to SCD, a1–3 missing data set, b4–6 missing data sets, c7–9 missing data sets, d10–12 missing data sets, elower scores indicate better performance. HC, healthy control; SCD, subjective cognitive decline; AD, Alzheimer’s disease.
MR procedure
Data acquisition
All SCD and prodromal AD patients underwent FDG-PET and structural MR brain imaging as part of the clinical routine at the Memory Clinic Cologne Jülich. Anatomical MRI and resting-state (RS)-functional (f)MRI data of every study participant were obtained at a 3T MR-Brain-PET scanner (Siemens, Erlangen, Germany) combining a 3T MR system MAGNETOM Trio with a custom-build BrainPET insert in the bore of the magnet using both a transmit-receive and 8-channel receive coil [59]. Vacuum cushions were used to reduce head motion. Automated and manual shimming was applied prior to all data acquisition to account for field inhomogeneities potentially resulting from the BrainPET insert. For functional imaging, a T2* weighted, single-shot echo planar imaging (EPI) sequence was used with the following imaging parameters: repetition time (TR) = 3000 ms, echo time (TE) = 30 ms, flip angle (FA) = 90°, field of view (FOV) = 200×200 mm2, matrix = 80×80, voxel resolution = 2.5×2.5×2.8, 50 oblique slices parallel to the infra-supratentorial line, gap = 0.28 mm, interleaved, scan time = 7 min. During this RS scan, participants were instructed to keep their eyes open, to not fall asleep, and to not think of anything in particular. For structural registration, a high-resolution image was obtained using a T2*-weighted EPI sequence with: TR = 3450 ms, TE = 30 ms, FA = 90°, FOV = 200×200 mm2, matrix = 128×128, voxel resolution = 1.6×1.6×2.8, 50 slices, gap = 0.28 mm, interleaved, scan time = 15 s. For anatomical reference, high-resolution T1-weighted structural images were collected using a magnetization-prepared rapid gradient echo sequence with:TR = 2250 ms, TE = 3.03 ms, FA = 9°, FOV = 256×256 mm2, matrix = 256×256, voxel resolution = 1 mm isotropic, 176 sagittal slices, no gap, interleaved, scan time = 5 min and 14 s.
Image preprocessing and data analysis
Image processing and statistical analyses were carried out using FMRIB’s Software Library tools (FSL, v5.0) [60, 61].
High-resolution T1-weighted structural images were skull-stripped with an automated, standardized method using FSL Brain Extraction Tool [62] with bias field correction [63–65] and manual editing if necessary.
Resting-state data were analyzed with FSL’s MELODIC ICA (Multivariate Exploratory Linear Optimized Decomposition into Independent Components) toolbox [66] (v3.1, http://www.fsl.fmrib.ox.ac.uk/fsl/fslwiki/MELODIC). ICA is a statistical technique to separate fMRI time series into independent spatial and temporal components and can be used i) to remove artefactual components, and ii) to identify neural networks that are activeduring rest.
Data preprocessing on single-subject level
The first 10 volumes were discarded for equilibration of the magnetic field and the remaining 130 images were pre-processed with the following procedures: motion correction [66], spatial smoothing using a 4 mm full width at half maximum Gaussian kernel, and high-pass temporal filtering at 0.02 Hz. Using a six-parameter, rigid-body linear transformation procedure in MCFLIRT [67], all functional images were realigned to the middle volume of the time series. Following this, the resulting functional images were registered to their realigned high-resolution functional images (3DOF), then to realigned structural images (6DOF), which were spatially normalized to the MNI152 standard space template (12DOF) using the linear image registration tool [67], and finally refined with the nonlinear image registration tool [68]. Data were resampled to 3 mm3 resolution and had a warp resolution of 10 mm3. To ensure adequate registration, each data set was visually inspected prior to furtheranalyses.
A common problem in analyzing RS-fMRI data is the presence of artefacts as physiological noise occurs at the same low frequency of < 0.1 HZ as spontaneous fluctuations measured by RS-fMRI. Such artefacts can produce spurious correlation patterns in RS-fMRI data and therefore need extensive data pre-processing, particularly in an elderly and clinical population [69]. The identification and removal of such artefacts is therefore of particular importance prior to further analysis. As standard linear motion regression methods often fail to fully remove the effects of motion, we opted for an accurate and objective cleaning approach of motion and other artefacts. First, we performed ICA on each individual resting state data set, which is a commonly applied technique to decompose data into independent spatial maps. The number of components was determined by an automatic dimensionality estimation implemented in MELODIC. Pre-processed images were then denoised using an objective automated tool provided by FSL (FIX FMRIB’s ICA-based X-noiseifier) [70]. FIX identifies and removes artefactual components with 95% classification accuracy based on temporal and spatial features including the sagittal sinus, cerebrospinal fluid, and white matter, and also adds motion regressors. This method has been proven to be an accurate, reliable, and consistent method of denoising fMRI data [71]. Resulting data were visually inspected according to the criteria proposed by Kelly et al. [72] for quality assurance. These cleanup steps resulted in the exclusion of one prodromal AD patient. To assure an improved and objective identification of artefacts in the cleaned data, we then applied the artifact detection tool (https://www.nitrc.org/projects/artifact_detect/).
Data preprocessing on group level
ICA-denoised preprocessed data were further used as input in a group-level ICA analysis using the temporal concatenation algorithm to obtain a single 4D file across all participants [66]. To correct for atrophy, partial volume information of grey matter was estimated and maps for all participants were created based on the T1-weighted images using the FSL script feat_gm_prepare. These maps were added as voxelwise explanatory variable in the GLM design [73]. Demeaned age and depression scores were entered as covariates of no interest. Resulting data were used for a voxel-wise between-subject analysis using the dual regression method implemented in FSL [74]. The dual regression method generates subject-specific spatial maps and time courses corresponding to each group independent component, required for use in FSLNets. However, instead of using the data-driven decomposed independent components from the group-level ICA, we selected predefined DMN nodes based on our hypotheses as input for the main analysis as we were particularly interested in investigating the connectivity strengths across DMN regions. We thus applied ICA using our predefined DMN regions to identify them independently as a preparatory step to derive specific group-level spatial maps to feed into the dual regression. They were carefully chosen according to Buckner et al. [22], and constrained within existing independent DMN templates [75, 76], containing the bilateral ventral medial prefrontal cortex (vmPFC) (Brodmann area (BA) 10; we condensed the vmPFC to BA 10 excluding BA 32 and 24 and verified the anatomical accuracy with a current DMN template by Smith et al. [76] resulting in our mask including parts of the superior and middle frontal gyrus (but will refer to this hub as the vmPFC for the remainder of the paper), PCC (BA 23, 31), inferior parietal lobule (IPL) (BA 39, 40), and in addition the RSC (BA 29, 30), and both hippocampi. Conforming to the method by Franco et al. [75], we identified these nodes using BAs in the Wake Forest University Pickatlas toolbox [77], which is based on histological and cytoarchitectonical observations and therefore allows for a functional and anatomical accurate selection of DMN nodes of interest (Fig. 1, left panel). To gain further insight into the interaction of the hippocampus with other DMN nodes, we ran a second analysis with the hippocampus excluded from the overall DMN mask, thus contrasting the remaining four bilateral DMN nodes (merged into one 4D file for both hemispheres) (Fig. 1, right panel) against both hippocampi.
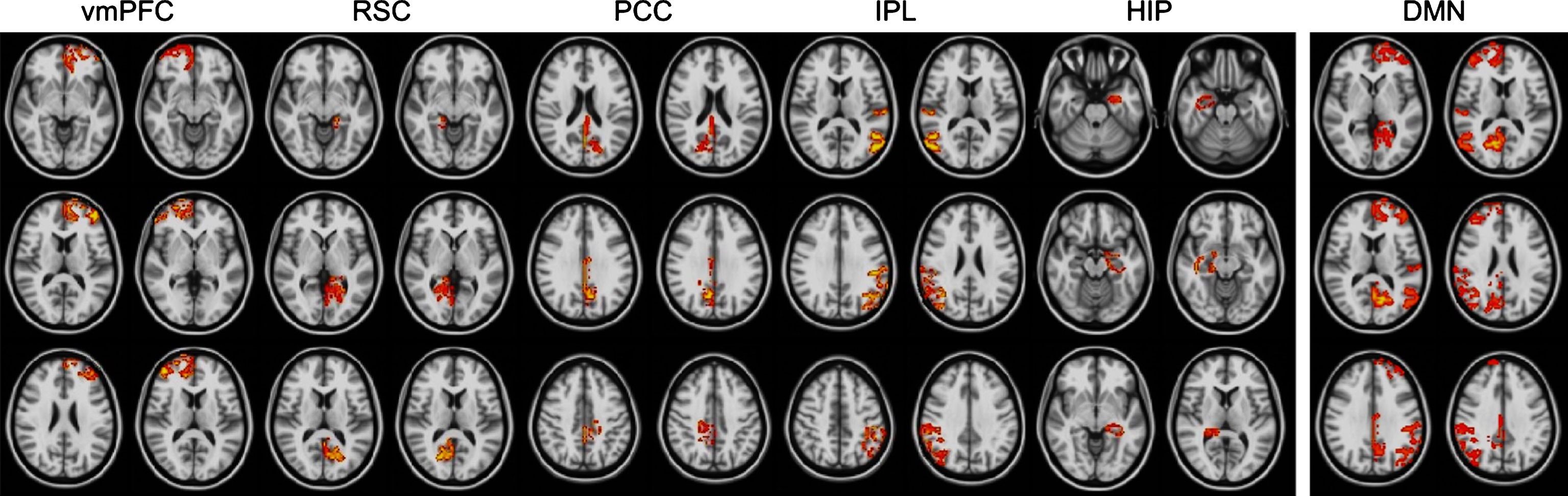
Spatial layout of selected DMN nodes (bilateral). Shown are the vmPFC, RSC, PCC, IPL, and hippocampus (from left to right) (left panel) and the vmPFC, RSC, PCC, and IPL merged into one 4D file (right panel). Images are displayed in radiological convention. DMN, default mode network; vmPFC, ventral medial prefrontal cortex; RSC, retrosplenial cortex; PCC, posterior cingulate cortex; IPL, inferior parietal lobule; HIP, hippocampus.
To ensure the selected DMN nodes only included voxels belonging to the grey matter, we applied a segmented grey matter mask, created with FSL Automated Segmentation Tool [78]. These were then merged into a single 4D file comprising ten volumes representing the five bilateral DMN nodes to be further used as spatial regressors against each individuals 4D fMRI dataset for the main analysis. This resulted in ten subject-specific timecourses, one for each DMN node (stage 1), which were then regressed into the same 4D datasets, yielding a set of subject-specific spatial maps, again one for each DMN node (stage 2). For the second analysis, this procedure remained essentially the same, except the resulting 4D files consisted of four volumes, the bilateralhippocampus and DMN.
Within-group comparisons of functional connectivity strength of DMN nodes using FSLNets
Functional connectivity strength across DMN nodes was computed using the FSLNets toolbox (v0.6) (http://fsl.fmrib.ox.ac.uk/fsl/fslwiki/FSLNets), implemented in MATLAB (http://www.mathworks.com/index.html). Each participants’ time series from the ten (or four in the second analysis) DMN nodes generated during stage 1 of the dual regression method was used as input to compute both full (indirect network connections) and regularized partial (direct network connections) correlation matrices. The partial correlation matrix provides a better estimate of direct connection strengths than the full correlation does as all other time series are regressed out of any given pair of time series before the correlation is estimated, controlling for the effects of all other brain networks [79]. Both correlation coefficients were then converted into z-statistic (using the Fisher r-to-z transform) and empirically corrected for a temporal autocorrelation in the data (i.e., every node correlating with itself). Resulting nodes were hierarchically clustered based on full correlation matrices. This hierarchy descriptively displays both full and regularized partial correlation matrices (the latter using ridge regression to improve the mathematical robustness of the estimation) [79, 80]. The regularized partial correlation matrix was further used to investigate the functional connectivity strength of each node-pair within the three groups using 5000 permutations with threshold-free cluster enhancement inrandomize [81].
Statistical analyses
Group differences of demographic and neuropsychological data were tested with a one-way analysis of variance (ANOVA) for continuous measures and a chi-square test for dichotomous measures using SPSS (v.21, SPSS, Inc., Chicago, IL). Post-hoc tests were corrected for multiple comparisons using the false discovery rate (FDR) at p < 0.05 [82].
Within-group analyses of all possible node-pairs were analyzed with randomize (see functional connectivity strength method section). Based on the outcome of these results, we performed post-hoc mediation analyses using the PROCESS macro for SPSS [83]. In these simple mediation models, we examined whether the RSC mediated (“indirect effect”) the relationship between the hippocampus and specific DMN nodes. To establish mediation, three steps are essential:
Step 1 does not require statistical significance according to most contemporary mediation theories [84].
Step 2 is important to estimate the prediction of RSC signal from the signal of the hippocampus.
Step 3 estimates each DMN node from the RSC as well as the hippocampus.
Of particular importance is the indirect effect of the hippocampus on each specific DMN node through the RSC derived from a1b1, which is the product of the effect of the hippocampus on the RSC a1 and the effect of the RSC on a selected DMN node b1. To test for the indirect effect, we applied the bootstrapping method with 5000 iterations at a 95% bias-corrected confidence interval. Bootstrapping analyses are suited for smaller sample sizes because the estimates are more robust to violations of normality. Additionally, we used the Sobel test as a conservative estimate of the standard error of a1b1 .
To compare the functional connectivity strength of DMN nodes between-groups, correlation values were extracted for each individual and every node-pair, corrected for age, depression, and atrophy. These were then entered in SPSS (v.21, SPSS, Inc., Chicago, IL) using a one-way ANOVA with the z-transformed correlation values between two specific nodes and the dependent variable and group as the between-subject factor. Post-hoc pairwise comparisons were FDR-corrected at p < 0.05 [82].
To examine the functional relevance of DMN nodal coupling we computed Pearson’s correlations of the individual correlation values of each node-pair (r-to-z transformed) and neuropsychological raw memory scores within-groups, FDR-corrected at p < 0.05 [82]. We then performed linear regression analyses to investigate between-group differences (with the prodromal AD group as reference) using two specific DMN nodes, group coded as dummy variables, and the interaction term between two specific DMN nodes and group as independent variables and neuropsy-chological raw scores as dependent variables.
Results
Demographical and cognitive characteristics
Demographic information of the included 78 study participants can be found in Table 1. There was a significant group difference in age (F (2,75) = 9.026, p < 0.001) (Table 1). While HC and SCD participants were of similar age, both were significantly younger than prodromal AD patients, as revealed by pairwise post-hoc analyses. Age was therefore included as covariate of no interest. Conversely, there was no significant difference in gender or years of education between study groups.
As expected, our results showed a significant group difference in cognitive performance (all p < 0.004) (Table 1). Post-hoc analyses confirmed that prodromal AD patients scored lower on all cognitive tests, including memory, attention, executive functions, visuo-spatial abilities, and language functions compared to both HC and SCD. The only exception was the Stroop reading subtest, which measures reading ability and speed of processing, during which prodromal AD patients performed significantly lower than HC only (Table 1). As anticipated, no significant differences for cognitive performance were found between HC and SCD.
Between network functional connectivity strength of DMN nodes
DMN nodes hierarchy within-groups
Figure 2A-C qualitatively displays the network hierarchies of all five bilateral DMN nodes in the three groups separately. In HC, all bilateral nodes form one main cluster that consists of a specific hierarchy starting with the PCC and RSC, which, in turn, can be grouped with the IPL, then with the vmPFC, and last with the hippocampus (Fig. 2A). Conversely, in both SCD and prodromal AD patients, the DMN is grouped into two main clusters. In SCD, the first cluster comprises the PCC, IPL, and vmPFC, while the second cluster consists of the RSC and hippocampus (Fig. 2B). In prodromal AD patients, the first cluster contains the parietal nodes (PCC, IPL, and RSC), and the second cluster the vmPFC and hippocampus (Fig. 2C).
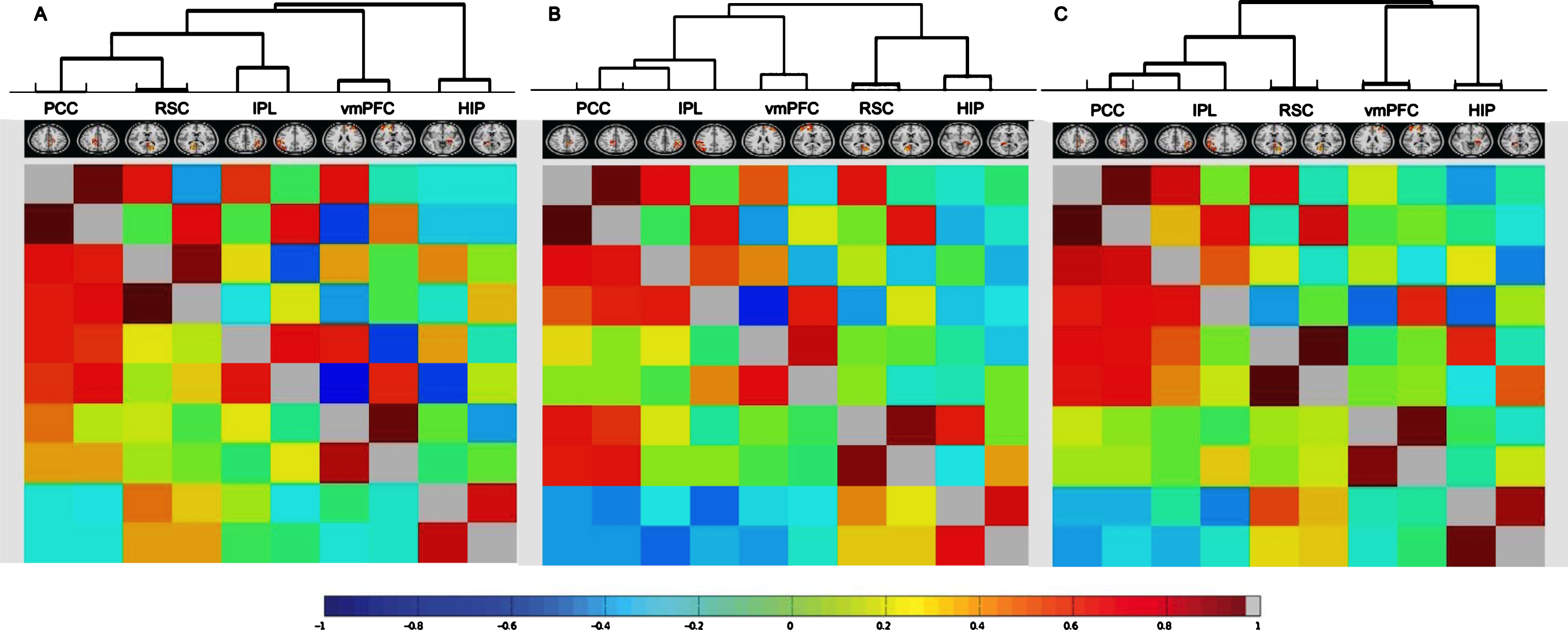
Full and partial correlation matrices of the five selected bilateral DMN nodes presented in a network hierarchy for healthy controls (A), subjective cognitive decline (B), and prodromal Alzheimer’s disease patients (C). Displayed are correlation coefficients between the time series of all five bilateral DMN nodes with positive correlations in red and negative correlations in blue as indicated by the color bar. Full correlation values are shown below the diagonal, regularized partial correlation values above the diagonal. Nodes are reordered bringing stronger correlated nodes together into clusters. DMN, default mode network; PCC, posterior cingulate cortex; RSC, retrosplenial cortex; IPL, inferior parietal lobule; vmPFC, ventral medial prefrontal cortex; HIP, hippocampus.
DMN nodal coupling within-groups
As expected, one-sample t-tests showed significant within-group correlations between each left- and right-sided DMN node (Supplementary Table 2). Figure 3 displays all significantly interacting node-pairs that survived significance thresholding within-group. These were all positively correlated to each other (Supplementary Table 2). The number of statistically significant node-pairs decreased with disease progression, i.e., 17 node-pairs in HC, 15 in SCD, and 12 in prodromal AD patients (Supplementary Table 2, Fig. 3). This decrease of functional coupling was particularly evident for i) the vmPFC and ii) the hippocampi (Fig. 3). To be more precise, the vmPFC was functionally coupled to the IPL, PCC, and ipsilaterally to the left-sided RSC in HC, to the IPL and left-sided PCC in SCD, but only ipslaterally to the right-sided IPL in prodromal AD patients (Supplementary Table 2, Fig. 3). The hippocampus was functionally coupled to the RSC and the ipsilateral left-sided IPL in HC, to the RSC in SCD, and to the ipsilateral left-sided RSC in prodromal AD patients (Supplementary Table 2, Fig. 3).
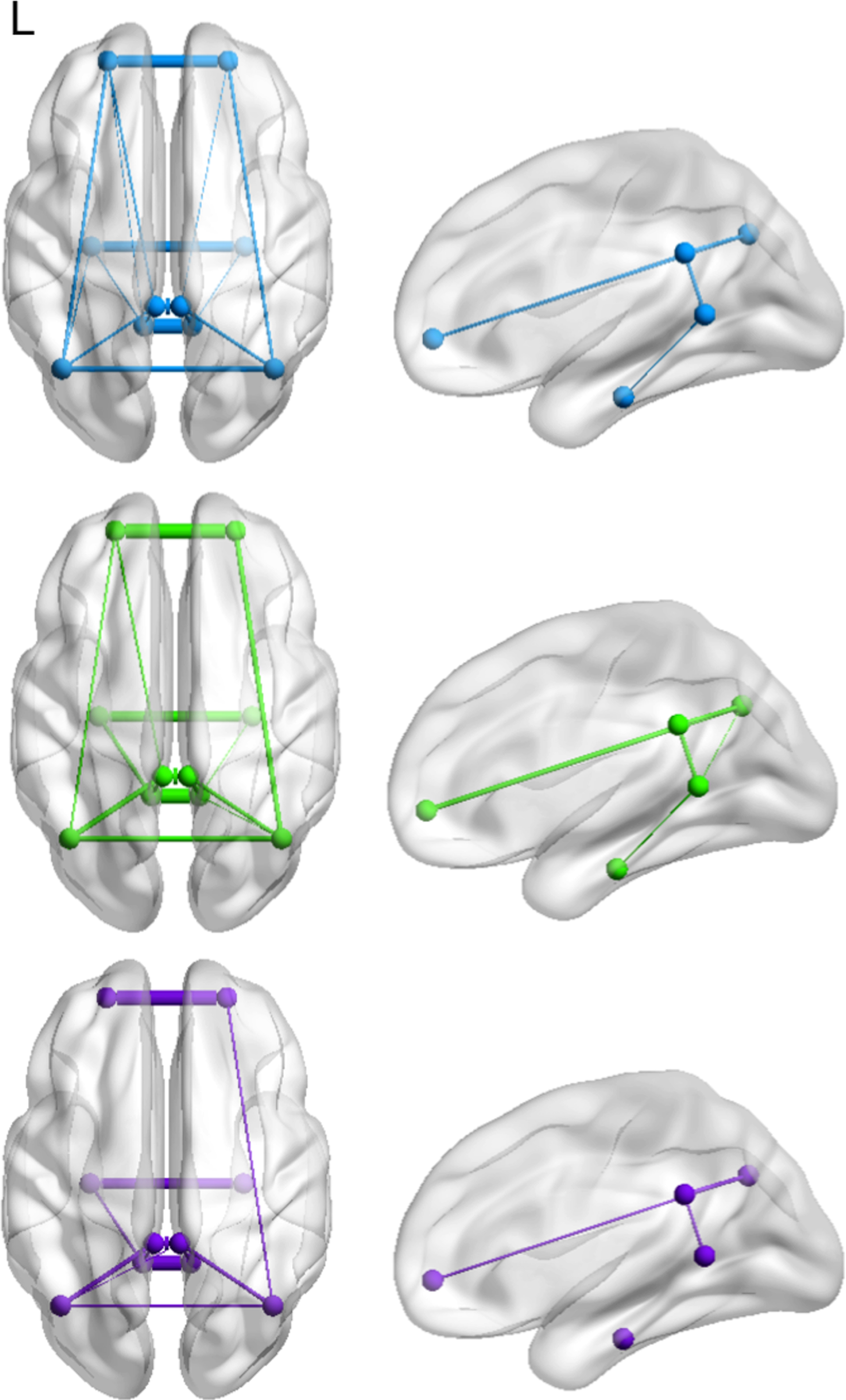
Significant coupling between default mode network nodes within healthy controls (blue), subjective cognitive decline (green), and prodromal Alzheimer’s disease patients (purple). The number of node-pairs decreased with disease progression. Thickness of lines indicates strength of correlation.
With respect to our third aim, the results revealed a strong positive correlation between the left- and right-sided DMN (all p < 0.001) as well as between both hippocampi (all p < 0.001) in all three study groups (Table 2). Additionally, there was a significant positive correlation between the left DMN and the left hippocampus in HC (p = 0.002), that was not found in SCD or prodromal AD patients. There were no between-group differences.
Mean z-scores reflecting coupling strengths between the combined nodes within the DMN and the hippocampi in all three study groups
Values represent regularized partial correlation values (r-to-z transformed)±SD with associated p-values. *significant at the 0.05 level (FDR-corrected). DMN, default mode network; HIP, hippocampus.
Post-hoc mediation analyses revealed that the RSC mediated the association between the hippocampus and vmPFC and IPL in HC only (Table 3, Fig. 4). There was full mediation by the right RSC for the relationship between the left hippocampus and right vmPFC (Table 3, Fig. 4, left panel). Furthermore, the right RSC partially mediated the relationship between the right hippocampus and right IPL in HC (Table 3, Fig. 4, right panel). There was no mediation by the RSC for any of the models in SCD or prodromal AD (Supplementary Table 3).
Simple mediation results with the left and right RSC as mediator located between the left (top) and right (bottom) hippocampus and all bilateral DMN nodes in healthy controls
Values represent path coefficients B±SD with associated 95% confidence intervals. Analyses were bootstrapped at 5000 iterations. All models were corrected for age, depression, and atrophy. Values surviving significance testing at p < 0.05 (FDR-corrected) are indicated in bold. RSC, retrosplenial cortex; DMN, default mode network; vmPFC, ventral medial prefrontal cortex; PCC, posterior cingulate cortex; IPL, inferior parietal lobule.
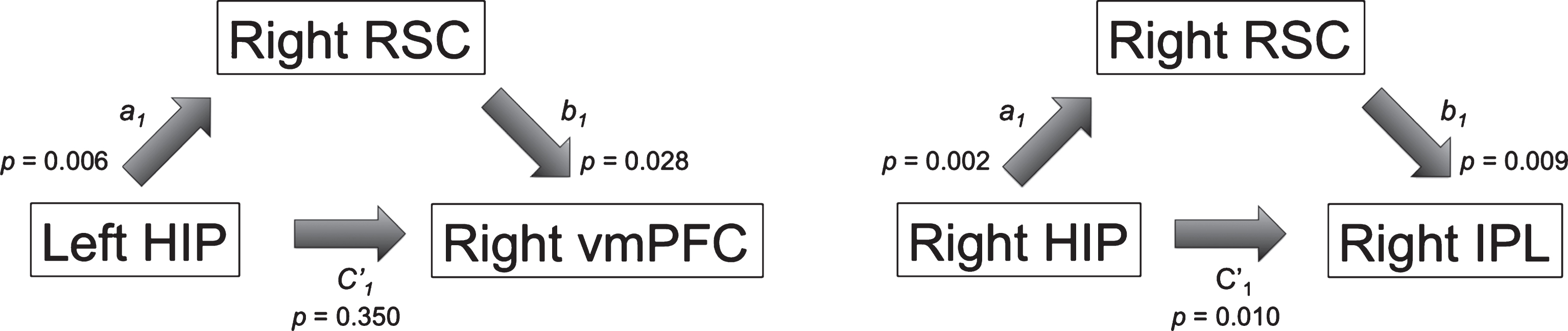
Statistically significant simple mediation model with the RSC as mediator M located between the hippocampus X and the vmPFC (left) and the IPL (right) in healthy controls (adapted from Hayes [83]). Results suggest full mediation by the RSC for the right vmPFC and partial mediation by the RSC for the right IPL. More detailed information on the regression formula involved and assessment of the indirect effect can be found in the methods section. RSC, retrosplenial cortex; vmPFC, ventral medial prefrontal cortex; IPL, inferior parietal lobule; HIP, hippocampus.
These findings suggest a dynamic network re-organization with disease progression involving functional impairment, but potentially also compensatory capacities, which could have important implications for early diagnosis and the development of newinterventions.
DMN node coupling between groups
The functional coupling between the left vmPFC and the left PCC (F (2,75) = 7.51, p = 0.001), right PCC (F (2,75) = 8.91, p < 0.001), and left IPL (F (2,75) = 7.89, p = 0.001) differed significantly across all three groups (Supplementary Table 2). Post-hoc analyses revealed a significantly lower positive coupling between the left vmPFC and left PCC in prodromal AD patients (0.35±1.16) compared to both HC (1.60±1.06) and SCD (1.16±1.26) (Fig. 5A). In contrast, the left vmPFC and right PCC were negatively correlated in all three groups with a weaker negative correlation in prodromal AD patients (–0.04±0.94) relative to HC (–1.21±1.16) (Fig. 5B). The correlation of the left vmPFC and the left IPL was positive and significantly lower in prodromal AD patients (0.40±1.50) compared to HC (2.15±1.73) (Fig. 5C).
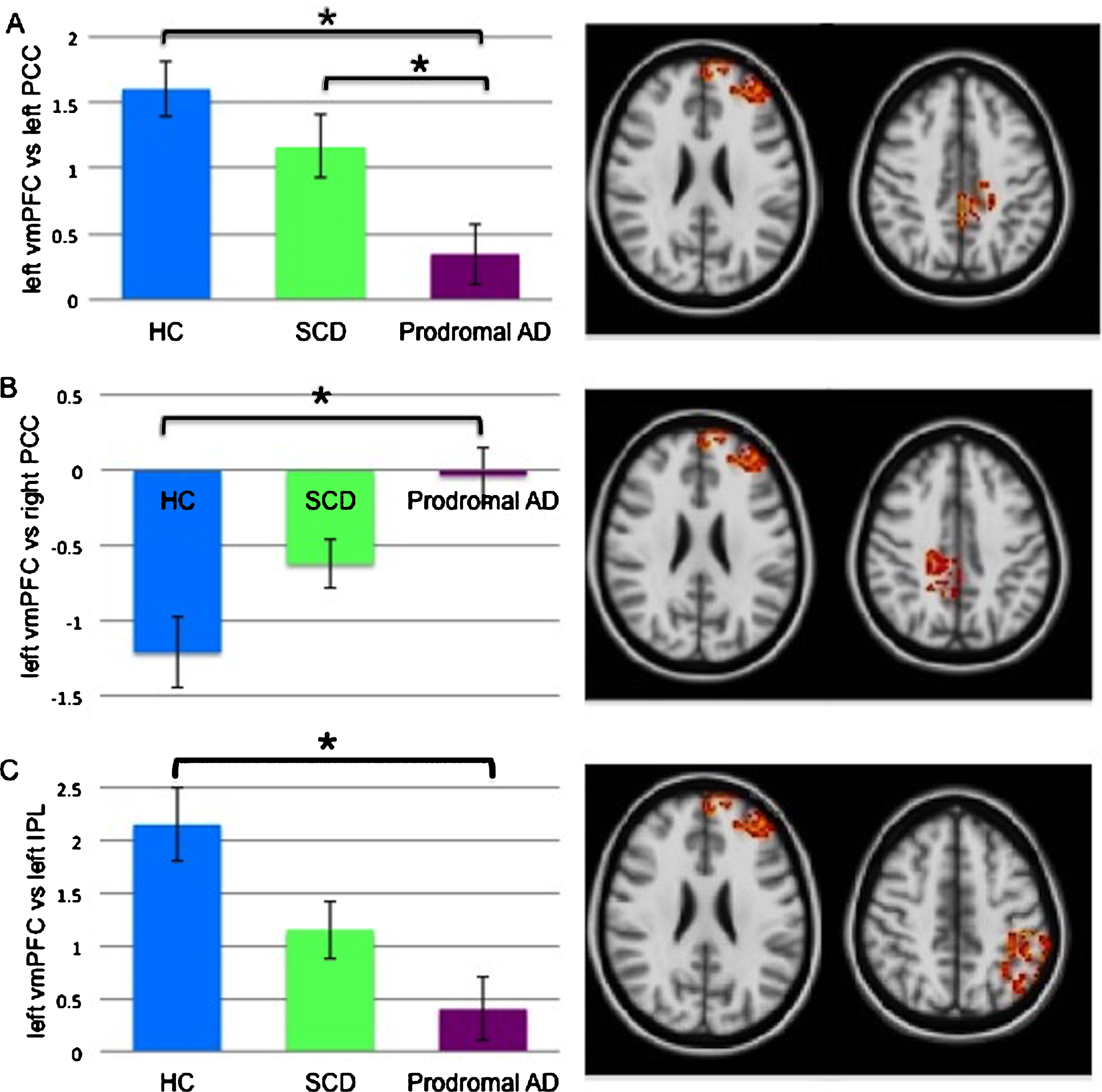
Significant coupling levels (r-to-z transformed regularized partial correlations) comparing the three groups. Between-group differences were found in the left vmPFC versus left PCC (A), left vmPFC versus right PCC (B), left vmPFC versus left IPL (C). Error bars indicate one standard error. *p < 0.05, FDR-corrected. vmPFC, ventral medial prefrontal cortex; PCC, posterior cingulate cortex; HC, healthy controls; SCD, subjective cognitive decline; AD, Alzheimer’s disease; IPL, inferior parietal lobule.
Associations with cognitive performance on the VLMT
With reference to our second goal, linear regression analysis for the left vmPFC/left PCC node-pair revealed that the interaction between functional coupling by group on delayed recall differed between prodromal AD patients and HC (ß= –1.47, t(77) = –2.27, p = 0.026) as well as SCD (ß= –1.68, t(77) = –2.92, p = 0.005) (Fig. 6). Within the prodromal AD group higher scores on the delayed recall were associated with higher levels of functional coupling between the left vmPFC/left PCC (r(23) = 0.641, p = 0.001). No within-group associations were found between functional coupling between the left vmPFC and left PCC, right PCC or left IPL and performance on total learning or delayed recall trials.
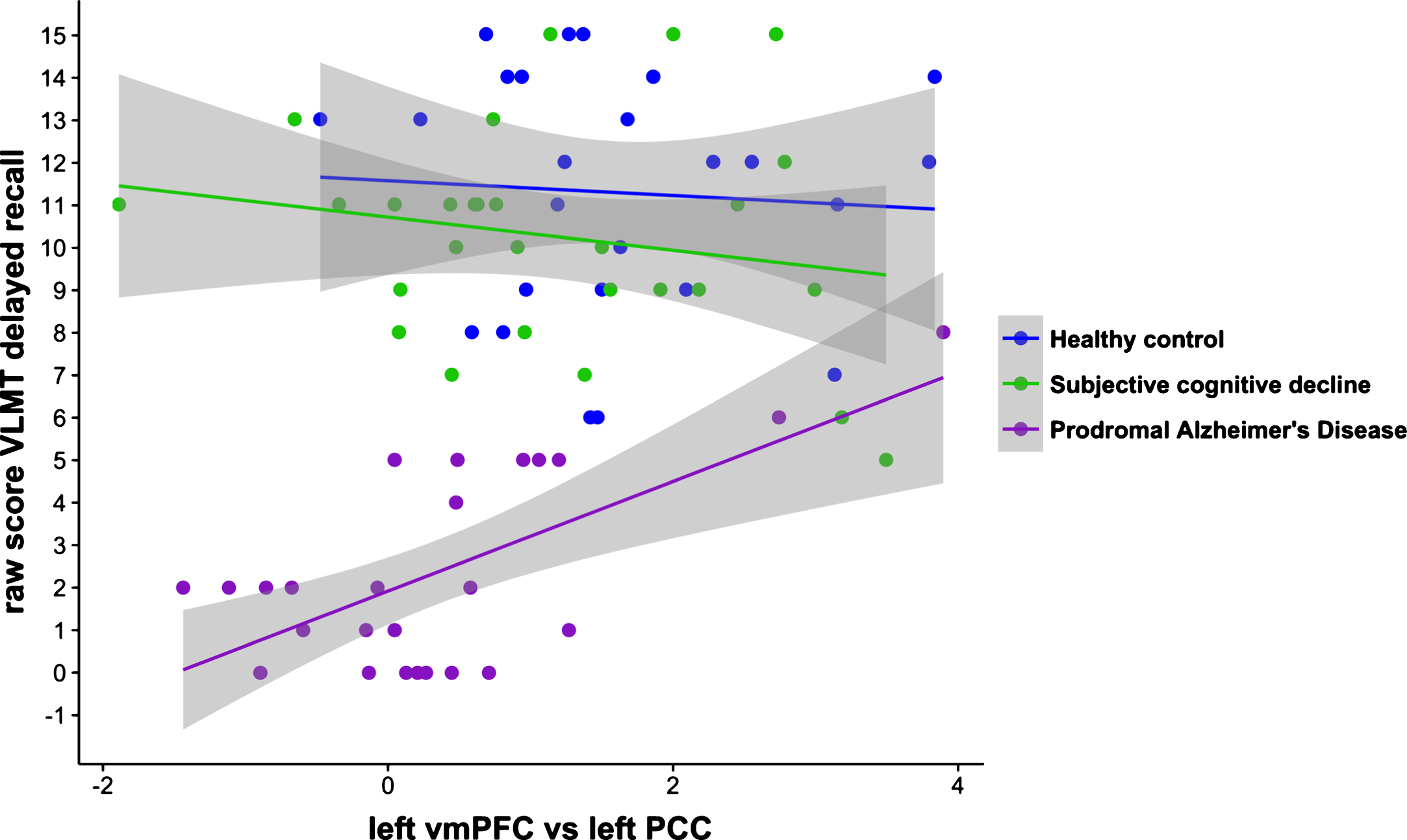
Significant associations between functional coupling of the left vmPFC/left PCC and the delayed recall on the VLMT for all three study groups. The positive correlation between this specific functional coupling and the VLMT in prodromal AD patients differed significantly from the negative correlation in both HC and SCD (p < 0.05, uncorrected). vmPFC, ventral medial prefrontal cortex; PCC, posterior cingulate cortex; VLMT, verbal learning memory test; HC, healthy control; SCD, subjective cognitive decline; AD, Alzheimer’s disease.
Discussion
The goal of the current study was to investigate the functional connectivity strength across DMN regions, including the hippocampus, along the continuum of healthy aging to prodromal AD. Our results extend earlier studies by conveying two key messages. First, we found disrupted functional coupling between frontal and parietal areas in prodromal AD suggesting a reduction of anterior-posterior coupling; positive cognitive–functional dynamic correlations suggest that the coupling between the left vmPFC and the left PCC might be able to differentiate prodromal AD patients with relatively preserved memory performance from those with memory decline. Secondly, our findings indicate that the hippocampi are only coupled with the posterior part of the DMN and become isolated from the DMN with disease progression. The RSC might form a gateway between the hippocampus and specific DMN regions.
Disrupted functional coupling of prefrontal and parietal DMN nodes in prodromal AD
AD has commonly been considered a disconnection syndrome. Numerous studies have linked AD-pathology and symptomology to disrupted functional coupling of cortical regions [85, 86] resulting in cognitive dysfunction [23, 87]. In support of the disconnection hypothesis, we found functional de-coupling between the vmPFC and posterior parietal regions, which is further supported by the different network hierarchies we found in prodromal AD compared to SCD and HC. In prodromal AD the PCC, IPL, and RSC form a main cluster. The role of these three parietal regions indicating a prodromal AD-related assembly of posterior parietal regions is well established in AD [23, 89]. This suggests that these regions segregate from remaining DMN nodes in prodromal AD, potentially resulting from the high metabolic activity and accumulation of amyloid plaques [22, 91]. The second cluster comprises the vmPFC and hippocampus. These two regions are prone to synapse degeneration and neuronal loss in AD, both likely to lead to memory deficits [92]. Our findings thus imply that both frontal and temporal regions act as one major circuitry affected in prodromal AD, independently from parietal regions. This segregation of parietal from frontal regions in particular might lead to memory loss in prodromal AD in contrast to individuals without quantifiable cognitive deficits, in which the vmPFC is part of a parietal-accentuated cluster. Our correlational analyses confirmed that the collaboration between the left vmPFC/left PCC is associated with better memory performance in prodromal AD compared to patients with lower and negative left vmPFC/left PCC correlations. Memory-related deficits can thus be explained by alterations in the functional connectivity strengths and interplay of DMN nodes [93]. The importance of interregional communication of parietal and frontal regions for episodic memory performance is well known [94, 95] and disturbed connectivity between PFC and PCC has been observed in AD [96]. Our findings suggest that a tighter positive functional coupling of frontal and parietal DMN nodes might help to preserve memory performance in the early stages of AD.
These results resemble past research on functional connectivity strength using graph-based analyses [10, 15]. In these studies, longer path lengths were found in MCI [12] and mild AD [10, 13], reflecting disrupted interactions of distant brain regions. This anterior-posterior disintegration has been explained by a breakdown of anatomical connections, e.g., through the cingulum bundle or the superior longitudinal fasciculus, which are particularly vulnerable in AD [97–99], and by an imbalance of parietal and frontal metabolism. The PCC in particular shows greater hypometabolism than frontal regions in AD [22, 100]. Future research could study the potential link of amyloid and tau pathology to this anterior-posterior disconnection.
Importantly, we did not find a significant difference in frontoparietal coupling between HC and SCD, which could indicate that not all SCD participants are characterized by an underlying AD-pathology or that these changes are yet too subtle to detect. The characterization of SCD individuals being at risk for AD is current work in progress [35]. For the definition of our SCD group, we followed the recently developed framework proposed by Jessen et al. [35]. Future studies could investigate the predictive value of enriching information about this group by reporting longitudinal stability [101] or including genetic data or biomarker evidence, which is of particular importance as individuals with SCD are twice as likely to develop AD as older individuals without complaints [102].
The lower positive functional coupling between left vmPFC/left PCC in prodromal AD compared to cognitively normal individuals is further supported by the different network hierarchies we found for SCD. These results indicate that, in contrast to prodromal AD, where the vmPFC was part of a second cluster, in SCD the PCC, IPL, and vmPFC work in coherence. Both metabolic [103] and neuroimaging studies [18] confirm the relevance and interplay of these DMN nodes in cognitively normal adults. Our results thus corroborate the importance of a coherent interchange of these DMN regions and suggest that any disruption to these regions might be associated with the progression to AD.
The overall declining trend of functional connectivity coupling within core DMN nodes with disease progression shows that the functional network connectivity of DMN regions not only decreases as neurodegeneration progresses but also as the risk to clinically manifested AD increases. Interestingly, our results show that the contralateral coupling of vmPFC/PCC has lower correlation values, i.e., direct connections, than its ipsilateral counterpart with prodromal AD showing significantly lower correlation values compared to both HC and SCD. While this remains speculative, this might be a result of asymmetric atrophy in AD [104] or the inherent lower direct anatomical connections of contralateral brain regions [105].
The hippocampus segregates with disease progression
To investigate whether the interaction of the hippocampus, a key region in AD, depends on the selection of specific DMN regions, we contrasted the hippocampi against an aggregate of the remaining DMN regions. Of interest is the positive correlation between the left hippocampus and remaining left DMN regions in HC, which vanished in both SCD and prodromal AD patients. This suggests that the hippocampus is coupled to the left-sided DMN in HC but that this coupling segregates from the DMN with disease progression. This is in line with previous studies [106] suggesting that the hippocampus becomes progressively isolated from the neocortex [107], which could contribute to memory deficits in AD [27]. Of interest is the de-coupling of the hippocampus and core DMN regions already in SCD, a group of individuals without any objectifiable memory deficits but subjective memory complaints. Our results thus confirm that pathological changes precede clinical symptoms [108, 109] and suggest a temporal process of hippocampal disintegration occurring prior to frontoparietal decoupling. Longitudinal neuroimaging studies are necessary to replicate this and to model temporal changes in functional coupling in thisinteresting group.
As these results show a left-sided hippocampus-DMN coupling, we examined hippocampal coupling to each DMN node in more detail and performed mediation analyses. Our results indicate that the hippocampus is interacting directly with the posterior but not anterior part of the DMN. Previous work already suggested that anterior and posterior DMN parts behave differently [110–112]. Most likely, the anterior DMN interacts with the hippocampus via indirect trajectories. We found that in HC, the RSC mediates the association between the hippocampus and other DMN regions. Thus, while our results confirm previous research on reduced hippocampal connectivity to posterior regions [113], they offer new insights into the importance of the RSC enabling the communication between the hippocampus and other DMN regions, suggesting that the functional connection between the hippocampus and DMN regions is indeed indirect and mediated by the RSC. Importantly, there was no mediation by the RSC in SCD or prodromal AD patients, again confirming that the hippocampus becomes isolated from the DMN very early on in the disease process. The underlying mechanism might be related to the propagation of tau pathology from the medial temporal lobe regions to associative cortical areas as a result of structucal and functional connectivity affecting regional neurons and white matter fibers connecting them[114, 115].
Limitations
There are two major limitations of our current study. Prodromal AD patients were significantly older than both HC and SCD. To account for this putative confound, we included age as a regressor in our statistical model. However, we cannot rule out that some of the explained variance might still be due to age effects. Future studies are needed to validate our results and exclude the possibility of a potential age bias of our data.
A second limitation relates to the fact that spontaneous fluctuations measured by RS-fMRI occur at the same frequency of < 0.1 HZ as physiological noise. These respiratory and cardiac fluctuations may affect the specificity of low-frequency fluctuations to functionally connected regions [116]. Such aliasing of physiological noise typically occurs at long TRs of 2-3 s as the temporal spacing of data points increases with the TR. Thus, ideally short TRs are recommended for a better sampling of the data. However, De Luca et al. [117] found similar spatiotemporally RSC patterns at both long (3 s) and short (0.12 s) TRs. We also found reliable results with a relatively long TR of 3 s.
Conclusion
Our data show that the hippocampus becomes progressively isolated from posterior DMN regions and that this process starts already in the preclinical stages of AD. An important node in maintaining communication between the hippocampus and the other DMN regions appears to be the RSC. In the prodromal phase of the disease, frontal and parietal DMN regions collaborate less, but a tighter functional coupling between frontal and parietal DMN regions is associated with better episodic memory performance in prodromal AD patients. Therefore, reduced DMN frontoparietal coupling may be used as a clinical tool for early identification of individuals at risk to manifestation of AD. These results may furthermore serve as groundwork for future intervention studies using non-invasive brain stimulation techniques focusing on altering the functional coupling between DMN regions to induce compensatory changes in prodromal AD thereby delaying memory decline.
Footnotes
ACKNOWLEDGMENTS
This work was supported by a DFG individual grant JA 2336/1-1 (HJ). GRF and JK gratefully acknowledge the support from the Marga and Walter Boll Foundation. We would like to thankDr. Gabriele Stoffels, Dr. Christian Filss, and Nathalie Judov for their assistance and generous support. We also acknowledge the technical support and advice of Prof. Dr. Hans Herzog, Dr. Elena Rota Kops, Lutz Tellmann, and Dr. Daniel Pflugfelder. Finally, we are grateful to Kornelia Frey, Suzanne Schaden, and Silke Frensch for their important help in data acquisition. Thanks are extended to Prof. Dr. Nadim Jon Shah for support with the MRI.