
Abstract
Alzheimer’s disease (AD) is a progressive neurodegenerative disorder characterized by cognitive deficits, deposition of amyloid-β (Aβ) plaques, intracellular neurofibrillary tangles, and neuronal cell death. Neuroinflammation is commonly believed to participate in AD pathogenesis. CD44 is an inflammation-related gene encoding a widely-distributed family of alternatively spliced cell surface glycoproteins that have been implicated in inflammation, metastases, and inflammation-linked neuronal injuries. Here we investigated the expression patterns of CD44S (which does not contain any alternative exon) and CD44 splice variants in postmortem hippocampal samples from AD patients and matched non-AD controls. The expression of CD44S and CD44 splice variants CD44V3, CD44V6, and CD44V10 was significantly higher in AD patients compared to non-AD controls. Immunohistochemistry of human hippocampal sections revealed that CD44S differentially localized to neuritic plaques and astrocytes, whereas CD44V3, CD44V6, and CD44V10 expression was mostly neuronal. Consistent with these findings, we found that the expression of CD44V6 and CD44V10 was induced by Aβ peptide in neuroblastoma cells and primary neurons. Furthermore, in loss of function studies we found that both CD44V10-specific siRNA and CD44V10 antibody protected neuronal cells from Aβ-induced toxicity, suggesting a causal relationship between CD44V10 and neuronal cell death. These data indicate that certain CD44 splice variants contribute to AD pathology and that CD44V10 inhibition may serve as a new neuroprotective treatment strategy for this disease.
Keywords
INTRODUCTION
Alzheimer’s disease (AD), characterized by progressive loss of cognitive function, is the most prevalent neurodegenerative disorder. The neuropathological features include amyloid-β (Aβ)-containing plaques, neurofibrillary tangles (NFTs) comprising aggregates of hyperphosphorylated microtubule tau protein, mitochondrial dysfunction, and the neuronal loss and synaptic dysfunction [1]. Aβ peptides are produced by sequential β- and γ-secretase-mediated cleavages of amyloid-β protein precursor (AβPP). The resulting Aβ peptides, specifically the oligomeric forms can modify synaptic plasticity or cause neuronal toxicity [2]. AD pathology is also characterized by a neuroinflammatory response, primarily driven by glial cell activation and production of cytokines and chemokines [3] as well as infiltration of blood-derived cells such as neutrophils [4]. The currently approved therapies provide only modest and transient symptomatic benefit.
The CD44 gene encodes a family of alternatively spliced class I transmembrane proteins. CD44 functions as a receptor for osteopontin and hyaluronic acid (HA), and has been implicated, together with its ligand HA, in several inflammatory diseases and cancer metastases [5]. The gene includes variable regions that are located in the extracellular membrane-proximal portion of the protein and are encoded by alternative spliced exons [6]. CD44S, which does not contain any variable exon (Fig. 1A), is the most ubiquitous form and is expressed by most cell types including lymphocytes and microglia [7, 8]. CD44 splice variant proteins, in which one or more of the 9 variable exons (10 in mouse) are included, are mostly reported in association with cancer [9] and autoimmune diseases such as rheumatoid arthritis [10] and multiple sclerosis [11, 12]. In the CNS, CD44 expression was reported in white matter glial cells, particularly astrocytes [13–15] and specific types of neurons [16–20]. CD44 splice variants containing exons V4, V5, and V10 were reported to be localized in neurons but not astrocytes in normal human brain [21] and increased in several CNS pathologies (reviewedby [22]).

A) CD44 gene structure with exon numbers (C for constant, V for variant) and expression in AD and control patients. Total RNA was prepared from postmortem hippocampal samples obtained from AD and age and sex-matched non-AD patients and cDNA was amplified by RT-PCR using exon-specific primers and the junctions were validated by sequencing. B) Dot plot presentation of relative expression data as analyzed by qPCR on mRNA samples obtained from AD and control patients for CD44S and three splice variants.
The role of CD44 in cell adhesion and migration of immune cells [23] astrocytes and microglia [24] may contribute to AD pathology [25–27]. CD44 gene was linked specifically to AD in a genome-wide association study, where CD44 was identified as a one of several genes that are associated as modifiers of the age of onset of AD in PSEN1 mutation carriers [28]. Akiyama et al. reported a specific subset of CD44 positive astrocytes whose number is increased dramatically in AD brains [29] and Uberti et al. [30] found that CD44 expression is increased in lymphocytes derived from AD patients in comparison with healthy subjects.
Interestingly, CD44 was shown to be a γ-secretase substrate of a dual intra-membrane cleavage [31], in addition to ectodomain cleavage by the metalloprotease ADAM10 (a disintegrin and metalloprotease10 [32]).
Here we explored the expression of CD44 isoforms in human AD brain specimen and report on increased expression of CD44S and three CD44 splice variants in the AD patients compared to non-AD controls. To explore a possible causal relationship between CD44V10 expression and disease pathology, we studied the expression of these variants upon stimulation of neuronal cells with Aβ peptides and the ability of CD44V10 knockdown (using a specific siRNA) and blockade (using a specific antibody) on Aβ-induced neuronal toxicity and on serum starvation induced cell death.
MATERIALS AND METHODS
Human tissue, RNA preparation, and RT-PCR analysis
Frozen postmortem human hippocampal tissue samples were obtained from The Netherlands Brain Bank (NBB, Netherlands Institute for Neuroscience, Amsterdam). All samples have been collected from donors for or from whom a written informed consent for brain autopsy and the use of the material and clinical information for research purposes had been obtained by the NBB. All material has been collected and used in compliance with the ethical and legal declaration of the Netherlands Brain Bank. Patients should have met the National Institute on Aging (NIA)-Reagan criteria for AD (Braak stage IV/C or higher with no vascular dementia, frontotemporal dementia, or Lewy body pathology [33]).
Total RNA was extracted from frozen hippocampal samples from AD patients (n = 10) and normal control individuals (n = 10) using EZ RNA kit (Biological Industries, Beit Haemek, Israel) and 1.5 μg of total RNA was reverse transcribed using Superscript II (Invitrogen, Carlsbad, CA). Semi-quantitative PCR was conducted on cDNA samples using oligonucleotides pairs in which one oligonucleotide was derived from variant exons V2-V10 sequences and the other from a 3’ or 5’ constant exon. Amplification products were purified from agarose gel (QIAquick gel extraction kit, Qiagen, Chatsworth, USA) and sequenced using one of the primers used in the PCR reaction. Sequence data was analyzed using BlastN algorithm on human RefSeq and genomic databases (NCBI). For quantitative analysis, qPCR was conducted on Applied Biosystems 7500 Real-Time PCR System using Platinum SYBR Green qPCR SuperMix-UDG (Invitrogen) reagent and the following oligonucleotides (500 nM): CD44S (F: gcgcagatcgatttgaatataacc; R: gggcaaggtgctattgaaagc); CD44V3 (F: gcacttcaggaggttacatcttttaca, R: aaactgaggtgtctgtctctttcatc); CD44V6 (F: agaatccctgctaccatccag, R: gaatgtgtcttggtctccagct); CD44V10 (F: gacagaatccctgctaccaatagg, R: ggaatgtgtcttggtctcctgat); GAPDH (F: ccccttcattgacctcaactac, R: gagtccttccacgataccaaag).
The oligonucleotides used for CD44V6 and CD44V10 amplification are located on variant-constant exon junctions aimed to amplify the cDNA species that include only the targeted variant exon without additional variant exons. Single band products were verified using agarose gel electrophoresis and sequencing. The reaction mixture was first denatured at 95°C for 2 min and then 40 cycles were performed using the following protocol: 94°C for 15 s.; 60°C (or 1°C below lowest primer TM) for 30 s.; 72°C for 30 s.
Standard curves were performed for each pair of primers, to confirm linearity of the qPCR reaction and in order to transform the Ct values to relative expression values.
qPCR using TaqMan probes
The Following TaqMan sets of primers and probes (5’-3’) were used to quantify the expression of CD44 mRNAs in the various gene expression experiments:
The qPCR reactions were performed in Applied Biosystem (ABI) 7300 Real time PCR. The reaction mixture was denatured at 95°C for 2 min and then 40 cycles were performed: 94°C for 15 s.; 60°C for 60 s.
Cell culture
SK-N-SH human neuroblastoma cell lines were obtained from the ATCC. Cells were maintained in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal calf serum, L-glutamine, and 1% penicillin-streptomycin mix (Biological Industries). In some of the experiments SK-N-SH cells were treated with 3 μM retinoic acid (Sigma-Aldrich, St. Louis, MO) for 5 days to allow the differentiation of the cells toward neuronal cells.
Frozen primary embryonic cortical C57/bl mouse neurons were obtained from Lonza (Basel, Switzerland) and maintained according to the manufacturer’s instructions. Primary embryonic cortical E15 Wistar rats neurons were prepared at Neuro-Sys SAS (Gardanne, France) as described by Callizot et al. [34]. Human Aβ1-42 peptide (rPeptide (Bogart, GA, USA) or Bachem (Torrance, CA, USA) treatment of the cells was done after pre-incubation of the amyloid peptide in serum-free medium for 72 h at 37°C. Each experiment was repeated at least 2 times.
Knockdown of CD44 expression by siRNA
At least 3 siRNA sequences were screened for effective knockdown of relevant transcript using HeLa and B16-F10 for human and mouse-directed siRNA sequences, respectively. Cells were plated at 1.5×105 cells in 24 well plates and transfected the day after with the different siRNA molecules (10 nM, IDT) using Lipofectamine RNAi Max (Invitrogen) according to manufacturer’s instructions. For analysis of siRNA efficiency, total RNA was isolated from the cells 48 h post transfection and reverse transcribed into cDNA using SuperScript II reverse transcriptase (Invitrogen). cDNAs were subjected to semi-quantitative PCRs or TaqMan qPCR using β-Actin as the reference gene. Effective siRNA sequences (sense strand) used for functional analysis are (3’ tt or uu are overhangdinucleiotides):
Cell viability
The XTT (Biological Industries) and Alamar Blue (AbD Serotec, Kidlington, UK) were used as cells viability indicators according to the manual instructions. XTT (1:3 dilution with growth medium) was by the end of each experiment and incubated for 1-2 h. Medium Absorbance was measured at 420 nm. Alamar blue was added at 20 to 40-fold dilution to the growth medium, incubated for 0.5–2 h and the medium fluorescence (excitation/emission at 485/590 nm) was read.
Caspase3 activity assays
Activity and proteolytic processing of caspase 3 was measured by using the EnzChek Caspase3 Assay kit (Life technologies, Carlsbad, USA) according to the manufacture’s guidelines. The basis for the assay was the cleaving of the fluorogenic effector caspase substrate Ac-DEVD-AMC. This substrate yields a bright, blue-fluorescent product (excitation/emission maxima ∼342/441 nm) upon proteolytic cleavage. In brief, the cells were lysed and 150 μg of protein was incubated with 200 μM Ac-DEVD-AMC. The increase in fluorescence was measured after 30 min using a fluorescence plate reader.
Immunostaining for survival of rat cortical neurons
Done by Neuro-sys SAS (Gardanne, France). Rat cortical neurons were treated for 24 h of Aβ1-42 in the presence of 1 μg/ml CD44V10 polyclonal antibodies (Millipore, Billerica, USA) or CD44S mouse monoclonal antibody (clone Hermes-3, purified from hybridoma culture) or antibody-free control. Cells were fixed in ice-cold solution of 95% ethanol and permeabilized with 0.1% saponin. The cells were incubated with mouse monoclonal antibody against the neuronal-specific protein, microtubule-associated-protein 2 (MAP-2, Sigma-Aldrich) followed by incubation with Alexa Fluor 488 goat anti-mouse IgG (Molecular probe). The immune-labeled cultures were automatically examined with ImageXpress (Molecular device USA) at x20 magnification, 30 fields per well (representing ∼80% of the total surface of the well) from 6 wells were analyzed. The total number of neurons was automatically analyzed using Custom module editor (Molecular Devices, USA).
Immunohistochemical staining of hippocampal sections
Done by Premier Laboratory (Longmont, CO, USA). 4-5 μm sections were prepared from formalin fixed, paraffin embedded hippocampal tissues (Netherlands Brain Bank, Amsterdam, Netherlands). Sections were de-parafinized in Xylene and rehydrated through a series of alcohol treatments. Endogenous HRP activity was blocked with 3% H2O2. Antigen retrieval was achieved by proteinase K or by heating. After blocking in Protein Block Serum Free (Daco, Santa Clara, USA), Primary antibodies used were anti CD44S (HCAM, rat clone IM7, Santa Cruz, Dallas, USA), CD44V3 (mouse monoclonal vff-327, abcam, Cambridge, UK), CD44V6, and CD44V10 (both rabbit polyclonal, Millipore, USA). References for the specificity of the primary antibodies are given in the vendors’ webpages (e.g., [35] for IM7) Secondary antibodies used were against the relevant species conjugated to HRP, polymer-enhanced (Envision®, Dako, Santa Clara, USA), using DAB+ as a substrate which was followed by hematoxylin staining. Negative controls were isotype matched serum controls for the rabbit polyclonal antibodies; it was non-immune rabbit serum at the same protein concentration as the primary antibody, and for the mouse antibodies, it was a multiple isotype control (Dako). For the rat antibody, it was a matched isotype control at the same protein concentration as the primary antibody.
Akt activation
SK-N-SH cells were grown for 24 h and then medium was replaced with serum-free medium and incubated for the indicated time in the presence or absence of anti-CD44V10 rabbit polyclonal antibody (Millipore, USA). Cells were lysed using RIPA lysis buffer (Sigma-Aldrich) supplemented with CompleteTM and PhosSTOPTM (protease and phosphatase inhibitor cocktails, respectively, Roche diagnostics, Switzerland). Protein concentrations were determined with PierceTM BCA protein assay kit (ThermoFisher, USA). Lysate samples were separated on 10% SDS-polyacrylamide gel electrophoresis (PAGE). The gel was blotted and probed with anti-phospho-Akt (Ser473) and pan-Akt antibodies (Cell Signaling, USA). Band intensities were determined using the ImageJ gel analysis program (NIH, USA).
Statistics
All statistical analyses were done using 2-tailed, unequal variance Student’s t test. Data are expressed as mean±SEM. p values less than 0.05 were considered significant.
RESULTS
Expression of CD44V10 and other specific splice variants is enhanced in human AD brain hippocampus
Expression of CD44 transcripts was studied in postmortem hippocampal tissue specimens obtained from 10 AD patients and 10 cognitively and pathologically normal individuals with similar age and gender characteristics (Table 1). CD44S and various CD44 splice variants were initially identified by RT-PCR amplification and sequencing. To amplify the variant-containing transcripts, we used primers derived from all 9 human CD44 variant exons (Fig. 1) with the neighboring 3’ and 5’ constant exons (C5 and C6). Sequence analysis indicated multiple CD44 splice variants in the hippocampus that contained one or more human variant exons V2 to V10 (Fig. 1A and data not shown). Three of these identified transcripts, CD44V3, CD44V6, and CD44V10, containing a single variant exon, were selected for further characterization. Expression analysis by real-time PCR (qPCR) performed on the hippocampal samples indicated that the expression of CD44V3, CD44V6, and CD44V10 variants was significantly higher in AD patients compared to the non-AD controls (Fig. 1B and Table 2). Significant upregulation was also observed when a set of primers that amplify constant exons were used, representing the entire CD44 transcripts population. Since the latter population is predominated by CD44S, we refer to the respective RNA and protein products as CD44S. It is important to mention that the PCR amplification of CD44 splice variants required larger number of PCR cycles compared to CD44S (6–8 cycles), indicating a lower expression level of these variants compared to the standard form.
Main characteristics of AD and non-AD subjects in this study. n, number of subjects in each group; Age, age at death; Braak, Braak stage; PMI, postmortem interval (h). Values are mean±SEM
qPCR analysis (SYBR green, normalized to GAPDH) of CD44S and three splice variants which include a single variant exon (CD44V3, CD44V6, or CD44V10) showed significant higher expression in AD patients (n = 10, 2 technical repeats for each sample) compared to control, none AD individuals (n = 10). p values are shown as determined by unpaired Student’s t test
CD44V10 immunoreactivity is localized to AD hippocampal neurons
Immunocytochemical analyses of CD44s, CD44V3, CD44V6, and CD44V10 derived proteins indicated that these proteins were localized histoanatomically and at the cellular level in hippocampal tissue sections. Preparations stained for CD44S were readily distinguishable due to striking immunoreactivity in cortical senile plaques. The reaction product densely filled neuronal perikarya and extended into proximal dendrites, as well as smaller caliber, isolated neuronal processes. Positively stained astrocytes are likely present, as well. Although neuronal somata outside of the plaque were usually unstained, presumptive neuronal processes were commonly positive, although not abundant, even remote from the plaques (Fig. 2A). Positively stained astrocytes were also detected in a cerebellar section (data not shown).
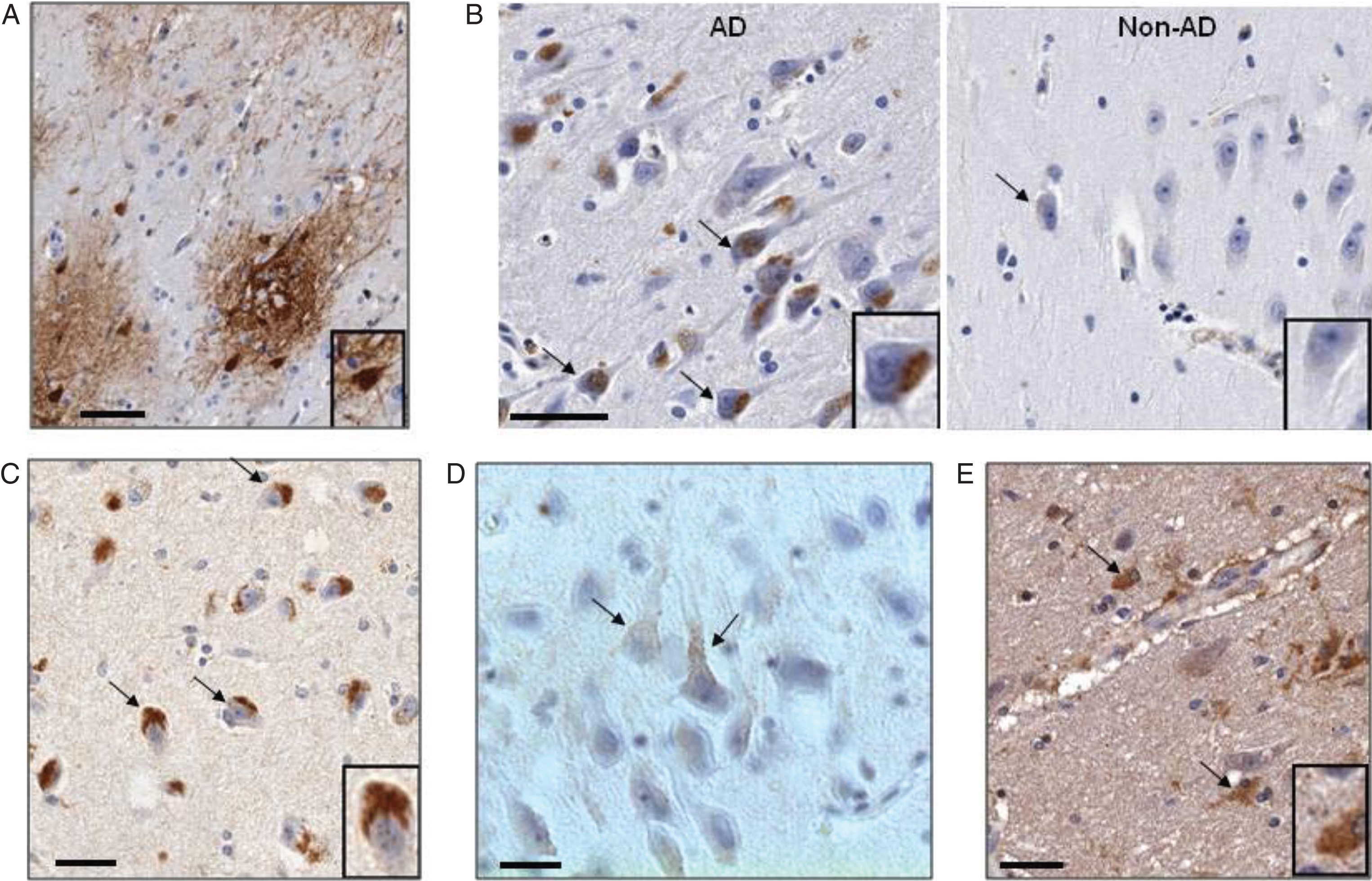
Localization of CD44 splice variants in postmortem hippocampal tissues of AD and control subjects. Immunohistochemistry (IHC) was done on sections of paraffin embedded hippocampus. A) IHC for CD44S showing plaque immunoreactivity involving neuronal somata, processes and astrocytes. The neurons are densely filled with reaction product (inset). Scale bar: 50 μm. B) IHC for CD44 V6 in hippocampal cortex: AD versus Non-AD. AD sample shows both perinuclear granular and superficial punctuate staining patterns. The small positive puncta are often connected with fine, immunoreactive linear structures, probably pre-terminal neuritis (arrows), more clearly appreciated as such in (C). Non-AD sample is minimally reactive. Inset: higher magnification of reactive neuron. C) IHC for CD44V10 with staining pattern similar CD44V6 (2B). D) IHC for CD44V6 in AD: Punctate staining of neuronal surface membrane in the hippocampal formation (CA) reminiscent of the appearance of axonal boutons terminating along several somata and their apical dendrites. E) In white matter, V3 showed dense cytoplasmic staining in astrocytic soma and processes; the latter were found perivascularly, in a typical distribution. AD cases and controls were not distinguishable.
Splice variants CD44V3, CD44V6, and CD44V10 protein products’ immunoreactivity was evident among neuronal cell bodies, and differed from respective findings for CD44S in several ways. Firstly, none of these variants stained elements of plaques. Secondly, neuronal somata never stained as completely or intensely as with CD44S (Fig. 2B, C). In the many immunoreactive neurons, reaction product was distributed both intracellularly and superficially. The intracellular staining was granular, and at least partially overlapped the perinuclear region of lipofuscin autofluorescence (Fig. 2B, C). The second type of neuronal staining consisted of scattered punctate loci that appeared localized to the cell membrane (Fig. 2D). These puncta were occasionally seen to be connected by fine, short processes, suggesting afferent terminal boutons. The staining of these bouton-like, afferent terminal structures was most clearly demonstrated in a CD44V6 stained AD case. In that sample, apical dendrites and/or the perikaryon of many neurons was distinctly outlined by a linear array of such structures (Fig. 2D). Regarding neuroglial staining, the majority of identifiable astrocytic structures were unstained in cortex, although occasional perivascular staining of glial foot-processes was observed. In contrast, astrocytic white matter staining regularly occurs with CD44S and CD44V3 (Fig. 2E) but not with CD44V6 and CD44V10. All components of oligodendrocytes appeared negative for CD44 immunostaining in all locations.
Since AD is on the spectrum of histopathological aging-associated changes in the brain, it was not surprising that some immunoreactivity was also seen in the non-AD, age matched control cases; with CD44S, even rare plaques were encountered. The primary difference was degree; neuronal staining in control cases was quite minimal and evident in only scattered neurons (Fig. 2B). In contrast, astrocytic white matter staining by CD44S and CD44V3 was somewhat comparable in all cases.
CD44 variants upregulation by Aβ1-42 exposure
We further tested the expression of CD44 splice variants in mouse primary cortical neurons. Using RT-PCR we validated the expression of CD44S, CD44V3, CD44V6, and CD44V10 in these cells (data not shown). We found that treatment of primary neurons with Aβ1-42 induced the expression of both CD44S, CD44V6, and CD44V10 at the mRNA level as evaluated by qPCR (Fig. 3A). CD44V6 and CD44V10 were also found to be induced by Aβ1-42 in the human neuroblastoma cell line SK-N-SH (Fig. 3B). SK-N-SH cell line was selected owing to previously published characterization of CD44 splice variants expression in this cellline [36].
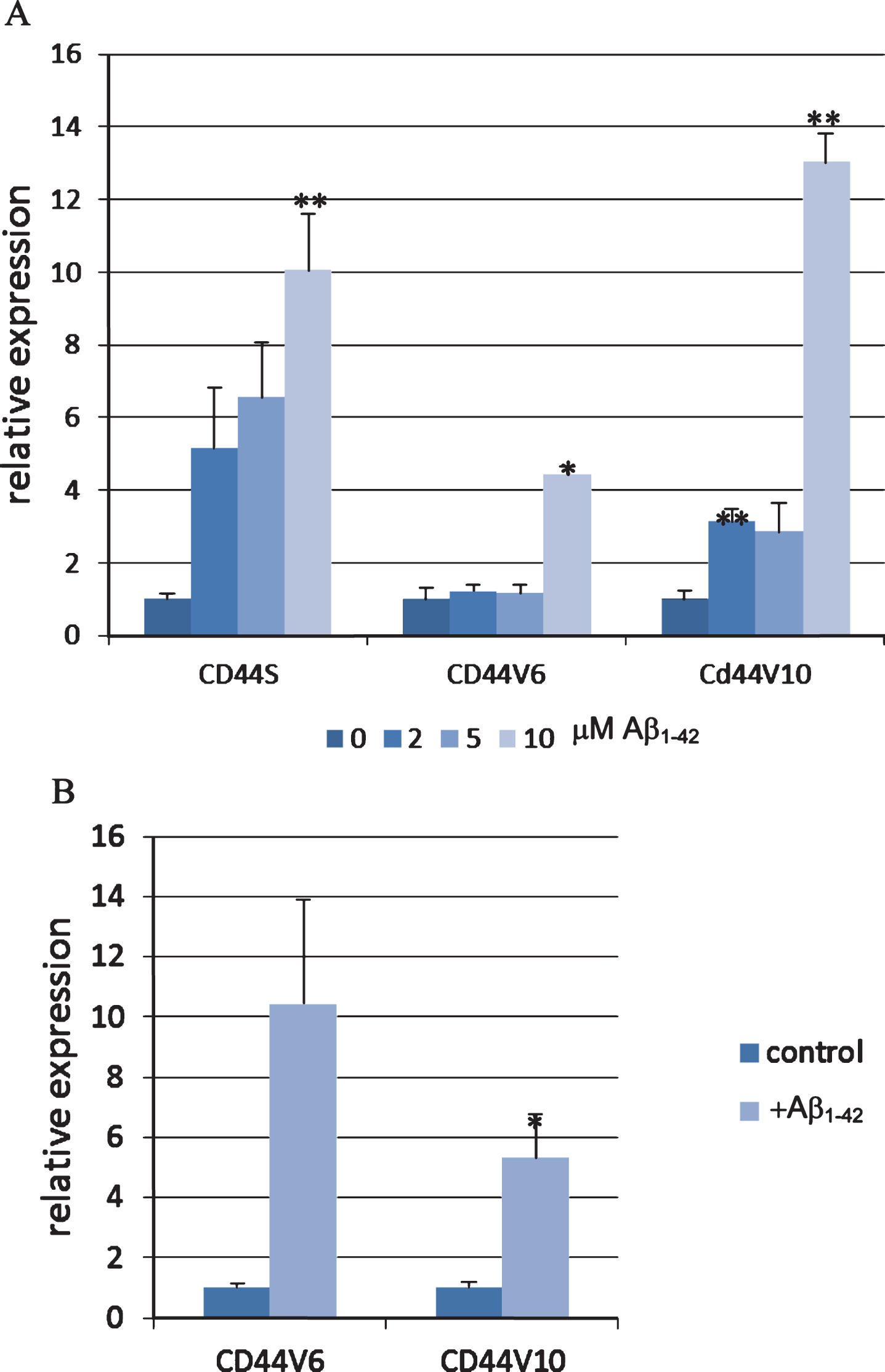
CD44 expression in neuronal cells is induced by Aβ1-42 peptide. A) Mouse primary neurons were treated with the designated concentrations of Aβ1-42 peptide for 20 h. The mRNA levels of CD44S, CD44V6, and CD44V10 were evaluated using qPCR normalized to β-actin expression (n = 2, 2 technical repeats). B) SK-N-SH cells were differentiated for 5 days using retinoic acid and then treated with Aβ1-42 peptide for 48 h. The mRNA levels of CD44V6 and CD44V10 were evaluated using qPCR normalized to β-actin expression (n = 3, 2 technical repeats). Bar graphs show means and standard errors. Significance: *p < 0.05, **p < 0.01 as determined by unpaired Student’s T test.
CD44V10 knockdown by siRNA and anti CD44V10 antibody are neuroprotective
To test whether CD44 splice variants are playing a functional role in AD-related neuronal cell death, we undertook a loss-of-function approach using exon-specific siRNA molecules. We first identified effective mouse and human siRNA molecules by checking reduction of relevant mRNA following transient transfection into relevant CD44-expressing cell lines (Supplementary Figures 1 and 2). We then tested the effects of the selected and non-relevant siRNA molecules on Aβ1-42 peptide induced cell death in SK-N-SH and in mouse cortical neurons using XTT or Alamar Blue as measures of viability and caspase 3 activity assay, as a marker of apoptosis (Fig. 4). As expected, incubation with Aβ1-42 resulted in marked increase in cell death (Fig. 4). Transfection of both cell types with CD44V10 siRNA selectively protected the cells against Aβ1-42 toxicity as evidenced by higher viability and reduced caspase 3 activation in the Aβ1-42 treated cells (Fig. 4), compared to non-relevant siRNA (NR). Partial but not significant protection was observed with CD44S siRNAs and no protection with CD44V6 siRNA in SK-N-SH and cortical neurons, respectively. This apparent neuroprotection was found consistently in both SK-N-SH neuroblastoma cells (Fig. 4A) and mouse cortical neurons (Fig. 4B), and was observed with two different V10 siRNA molecules, indicating that the effect is specific.

CD44V10-specific siRNA molecules protect neuronal cells from Aβ1-42 induced cell death. A) SK-N-SH cells were transfected with 10 nM of siRNA molecules with sequences corresponding to two different CD44V10 (V10) locations, CD44S (S) non-relevant (NR) or left untransfected (NT). After 24 h cells were treated with pre-aggregated 80 μM Aβ1-42 for an additional 24 h when cell viability was determined with XTT and induction of apoptosis evaluated by fluorescence caspase 3 activity assay (n = 2, 2 technical repeats). B) Mouse cortical neurons were transfected with different siRNA molecules in the presence (or absence) of 20 μM Aβ1-42 and incubated for 48 h. Alamar blue and caspase 3 activity assay were used for evaluation of viability and induction of apoptosis, respectively (n = 3, 2 technical repeats). Significance: p < 0.05, p < 0.01 as determined by unpaired Student’s T test.
We next tested the effect of commercial polyclonal CD44V10 antibody on Aβ1-42 toxicity in cortical neurons (Fig. 5). In agreement with the CD44V10 knockdown results, we found the CD44V10 antibody protected rat cortical neurons from Aβ1-42–induced cell death whereas CD44S specific antibody (Hermes-3) had no effect on the viability of these cells (Fig. 5A). We further explored the effect of the anti CD44V10 antibody on serum starvation induced SK-N-SH cell death and found that there was a significantly greater survival in the presence of the antibody compared to vehicle control (Fig. 5B). No protection against serum starvation induced cell death was seen with CD44S or GFP antibodies.
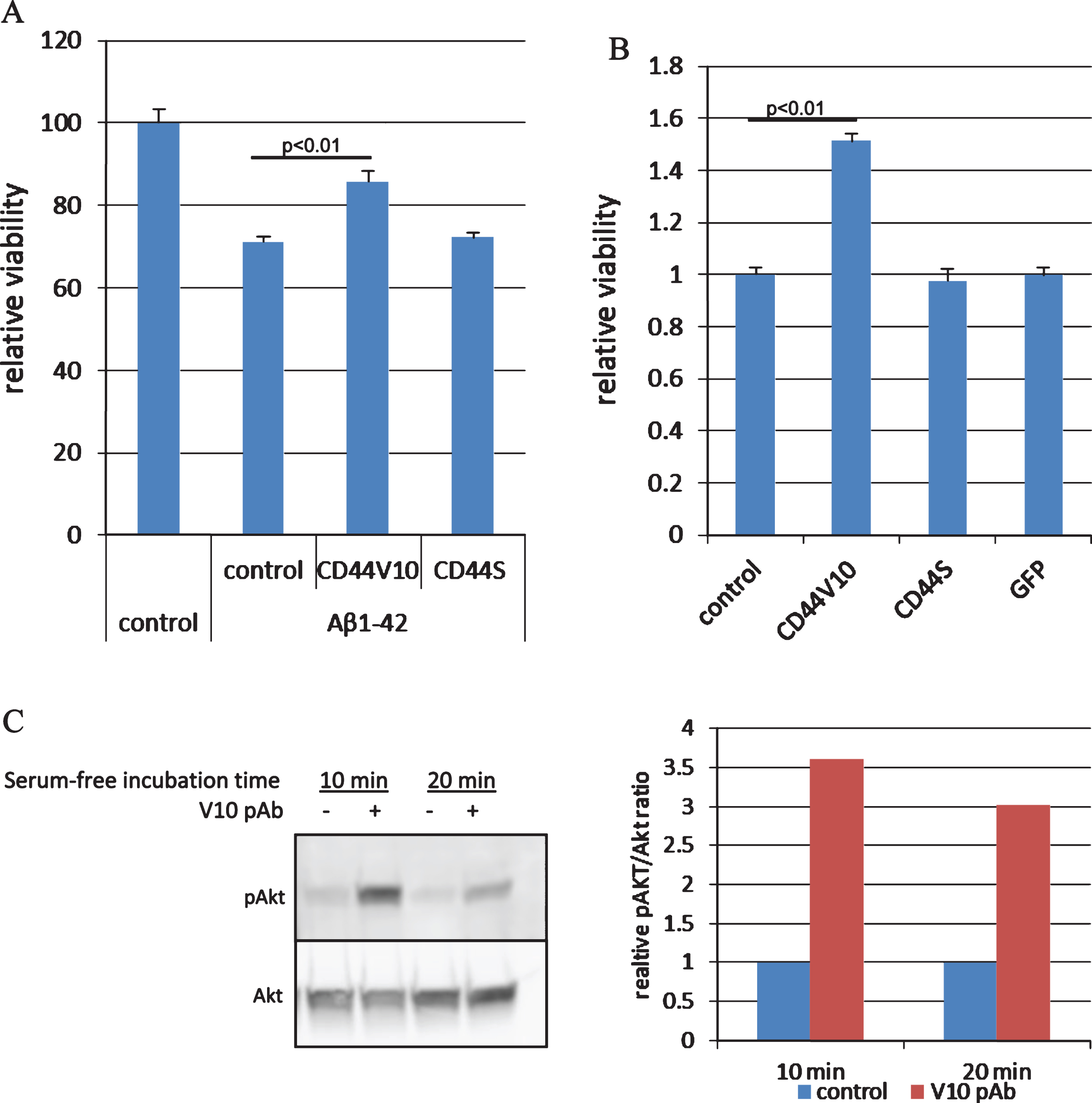
CD44V10 but not CD44S antibody protects cortical neurons from Aβ1-42 toxicity. A) Rat cortical neurons (E15) were treated with 20 μM Aβ1-42 for 24 h in the presence of 1 μg/ml CD44V10, CD44S antibodies, or no-antibody control, followed by immunostaining using MAP-2 antibody (a neuronal-specific protein). The number of MAP2-positive cells were evaluated and used as a measure for neuronal survival (n = 6). B) SK-N-SH cells were treated with serum-free medium for 48 h as an inducer of cell death, in the presence of 1 μg/ml CD44V10, CD44S, or GFP antibodies (n = 4). C) Immunoblot and the corresponding densitometry analysis of SK-N-SH cells following treatment with serum-free medium for 10 or 20 min in the presence or absence of 1 μg/ml CD44V10 antibody (V10 pAb). Detection was done by phosphor-Akt (ser473) and pan-Akt antibodies and densitometry analysis by the ImageJ program.
A potential role of CD44V10 in signal transduction
Accumulating evidence demonstrate that CD44 and its splice variants acts as a signaling hub controlling cell surface receptors, among them receptor tyrosine kinases (RTKs) [37]. Therefore we aimed at identifying downstream signaling cascade that may mediate the anti CD44V10 antibody neuroprotection. The PI3K/Akt pathway is activated by RTKs and is well characterized as an anti-apoptotic pathway which is known to mediate survival signaling in neuronal cells [38]. We found that CD44V10 antibody caused a fast and robust Akt activation upon transfer of SK-N-SH cells to serum-free medium compared to antibody-free control medium as reflected by an increase in the level of Akt phosphorylation on Serine 473 (Fig. 5C). AKT activation was not observed when the cells were treated with CD44S antibody (data not shown).
DISCUSSION
In this paper, we identified multiple CD44 splice variants in the hippocampus of AD patients. Of the various potential CD44 isoform transcripts, splice variants including CD44V3, CD44V6, and CD44V10 were found to be significantly upregulated in AD patients compared to age- and sex-matched control non-AD individuals. Immunohistochemical analysis of hippocampal sections showed that CD44V6 and CD44V10 proteins were localized in neurons. Although splice variants were predominantly expressed intracellularly, membrane associated staining in presumptive neuronal afferent terminals was also observed. In contrast, CD44S protein was localized to astrocytes and cortical senile plaques and CD44V3 was detected in both cell types. It is possible that the CD44S signal localized to senile plaques, represents soluble CD44 released from adjacent cells. The demonstration of CD44S immunostaining in astrocytes is in agreement with previous publications [21, 39]. Similarly, our findings of cytoplasmic localization of CD44 splice variants in human hippocampal cortical neurons, is in line with analogous observations on V4, V5, and V10 exon containing CD44 splice variants in normal human brain [21]. This data is compatible with a role of CD44 in AD pathophysiology.
The upregulation of CD44V6 and CD44V10 variant expression induced by Aβ peptide in neuroblastoma cells as well as in primary neurons, further supports a potential role of these splice variants in AD. Furthermore, loss of CD44V10 function in neuronal cells using CD44V10-specific siRNA and anti CD44V10 antibody, protected the cells from Aβ peptide-induced cell death. These data suggest a causal relationship between CD44V10 and neuronal death. We further found that the IP3K/Akt pathway is activated by the CD44V10 antibody. There are numerous papers indicate that this pathway is one of the central pathways that perform a pivotal function in mediating survival signaling in different neuronal cell types in response to growth factor stimulation (reviewed by Ahn [38]). To the best of our knowledge this is a first report describing a direct role of CD44 and specifically CD44 splice variants, in neuronal cell death.
CD44 seems to play an important role in the developing nervous system: it was shown to be essential for normal axon guidance during neuronal development [19] and dendritic arbor restriction [16], the latter through activation of the c-src tyrosine kinase and regulation of neuronal Golgi morphology. In agreement with the above is the finding that CD44 knockout mice show impaired hippocampus-dependent spatial memory retention and reduced sensorimotor function [40]. On the other hand, CD44 silencing by shRNA was shown to protect pyramidal neurons from glutamate-induced dendritic shortening [16].
The mechanism by which CD44V10 exerts its pro-apoptotic activity which is demonstrated here, remains to be elucidated. It has been well documented that CD44 constitute an important regulator of cell proliferation, survival and differentiation through interaction with and co-activation of several RTKs and initiation of intracellular signaling pathways [9, 37]. Specifically, this activity may be mediated through recently documented interaction of CD44V10 with the RTK EphA2 [41, 42]. Because Eph receptors were found to play a role in AD pathology (reviewed by [43]), a putative interaction of CD44V10 with EphA2 may provide the missing link. Since these RTKs are normally regarded as part of anti-apoptotic pathways [44], the suggested pro-apoptotic effects of CD44V10 in neuronal cells may be explained by negative regulation of CD44-RTK interactions by the V10 domain. This notion is in agreement with our finding that treatment of neuroblastoma cells with a CD44V10 antibody results in activation of the Akt pathway.
Another potential mechanism of action for CD44V10 is through cooperation with integrins and/or osteopontin, which have been reported in various physiological and pathological settings including neurodegeneration [45–47]. For example, an RGD-containing peptide fragment of osteopontin (which prevents binding of integrin ligands), was shown have a neuroprotective effect in stroke models [48] and to protect dopaminergic neurons from MPP+ and LPS toxicity [49]. In addition, the expression of osteopontin, which is a CD44 binding protein as well, is also increased in neurons of the AD brain [50].
CD44 role in cell death or survival was mostly investigated in various cancer cell lines and cells of the immune system. Most of the studies indicate that the interactions of CD44, predominantly CD44 splice variants, with HA contributes to resistance to apoptosis (reviewed by [51, 52]). However there are also examples in which CD44 crosslinking by HA or antibodies initiates or contributes to apoptosis [10, 53–55]. In the few studies of CD44 in animal and cell models of neurodegeneration the effect of CD44 was attributed to its known function in inflammation. Thus, CD44 knockout mice had a smaller ischemic infarct size compared to wild-type mice, following cerebral ischemia injury [56]. This effect was accompanied by selective reduction in inflammatory cytokines. There is also evidence to indicate that CD44 plays a critical role in the regulation of neuroinflammation in murine autoimmune encephalitomyelitis (EAE) model of multiple sclerosis [11, 57–59]. Moreover, it was shown by genetic ablation that CD44 splice variants V7 and V10 specifically participate in development of the EAE burden through their effect on the longevity of autoreactive T cells [12].
Overall, our data reveals a hitherto unknown role of CD44 splice variants and specifically CD44V10, in the neuronal cell death induced by Aβ and suggest that CD44V10 is a player in AD neuropathology. Therefore downregulation of CD44V10 expression and/or inhibition of its activity may potentially provide a new therapy for AD drug discovery.
DISCLOSURE STATEMENT
Authors’ disclosures available online (http://j-alz.com/manuscript-disclosures/16-1245r2).