
Abstract
Although tau protein was long regarded as an intracellular protein with several functions inside the cell, new evidence has shown tau secretion into the extracellular space. The active secretion of tau could be a physiological response of neurons to increased intracellular amounts of tau during the progression of tau pathology. We looked for potential differences in the serum levels of toxic tau oligomers in regards to cognitive impairment of subjects. We detected tau oligomers in the serum of Alzheimer’s disease (AD) patients, but they were also present to some extent in the serum of healthy older subjects where the levels positively correlated with aging (Spearman r = 0.26, p = 0.016). On the contrary, we found lower levels of tau oligomers in the serum of mild cognitive impairment (MCI) (p = 0.033) and MCI-AD (p = 0.006) patients. These results could suggest that clearance of extracellular tau proteins takes place, in part, in the periphery. In the case of MCI patients, the lower levels of tau oligomers could be the result of impaired clearance of tau protein from interstitium to blood and consequent accumulation of tau aggregates in the brain.
INTRODUCTION
Tauopathies are age-related neurodegenerative disorders that are characterized by the presence of abnormally phosphorylated tau aggregates. Alzheimer’s disease (AD), the most common tauopathy, is characterized by deposits of extracellular amyloid plaques and intraneuronal neurofibrillary tangles (NFTs) containing fibrillar tau protein [1, 2]. Tau, in its monomeric form, is natively unfolded which is essential for microtubule dynamics [3]. However, in pathological conditions misfolding occurs, thus enabling aggregation and formation of oligomers, which are considered to be the toxic forms of tau protein (rather than large, metastable tau aggregates such as NFTs) [4–12]. Although tau protein was long regarded as an intracellular protein with several functions inside the cell, new evidence has shown tau secretion into the extracellular space. First, the occurrence of tau in the interstitial and the cerebrospinal fluid (CSF) was believed to be a consequence of dying neurons and their released cell content. In the last few years, several proteins lacking signal peptide were shown to be secreted by unconventional pathways [13, 14]. Using cell cultures and transgenic mouse models, active secretion of tau protein from cells in the membrane-free “naked” form [15] or included in microvesicles/exosomes [16, 17] has been reported. The active secretion of tau could be a physiological response of neurons to increased intracellular amounts of tau during the progression of tau pathology. Therefore, secretion has been proposed as a mechanism to eliminate the excess amount of tau protein and thereby avoid its toxicity [17, 18]. In accordance with this, current CSF biomarkers of AD (total tau, phospho-tau181) are elevated when the pathology develops [18, 19]. In addition to these well-established tau measurements, we recently have demonstrated elevated levels of tau oligomers in AD CSF [20]. Moreover, in AD, the functions of the blood-brain and blood-CSF barriers are compromised. Several alterations are observed in the choroid plexus epithelial cells during aging and in AD [21]. These changes result in decreased CSF production, reduced amyloid-β clearance, increased glycation of proteins, and oxidative stress [21–27]. Such modifications may contribute to amyloid-β and tau protein oligomerization in CSF. The accumulated amyloid-β oligomers disrupt the integrity of the blood-CSF barrier, thus enabling uncontrolled passage of other proteins to the blood [28]. Nevertheless, no data concerning the occurrence of tau oligomers in the blood has been shown previously. Therefore, we were interested if the pathology of AD can be reflected in the blood. Moreover, collecting blood samples is an easier and less invasive intervention than lumbar puncture. To compare levels of tau oligomers with regards to cognition, we collected serum samples from cognitively normal subjects and individuals with cognitive impairment. We measured oligomers and conformers of tau protein in human sera by ELISA using novel anti-tau oligomer-specific rabbit polyclonal antibody T22 and tau conformation specific rabbit polyclonal TTC-99 antibody, respectively. T22 antibody recognizes oligomeric forms of tau in vitro and in vivo, but it is not reactive with monomers [29]. The antibody TTC-99 recognizes aggregated full length conformers and fragments of tau (unpublished data).
MATERIALS AND METHODS
Participants
All experiments were conducted in accordance with The Declaration of Helsinki. The study was approved by the Ethical Committees of Kralovske Vinohrady University Hospital and Prague Psychiatric Center, in accordance with the Laws 129/2003 and 130/2003 of the Czech Republic. Serum samples were obtained at AD Center, Charles University in Prague, Department of Neurology or Memory Clinic, Czech Republic. Serum samples were collected from 186 participants who were divided into four groups. The first group of non-demented controls consisted of 86 neurological patients with normal Mini-Mental State Examination (MMSE) [30] and normal basic CSF findings when patients were willing to undergo this examination [31]. The second group consisted of 18 patients with mild cognitive impairment due to Alzheimer’s disease (MCI-AD) [32], the third group consisted of 14 patients with mild cognitive impairment (MCI) not fulfilling the criteria for MCI-AD [33], and the fourth group consisted of 68 patients with dementia due to AD according to the NIA-AA criteria [34]. The patients had an established diagnosis of AD from an experienced neurologist. The diagnosis of patients with cognitive impairment was based on objective evidence of a progressive decline in cognition, functional and neuropsychological assessments, hippocampal atrophy seen on brain magnetic resonance imaging (or computer tomography in case of contraindications), temporoparietal hypoperfusion seen using single photon emission computed tomography, or increased total or phosphorylated tau protein and/or decreased Aβ concentrations in CSF using cut-offs established in our previous studies [35–37] when patients could undergo or were willing to undergo these examinations.
Serum samples were collected, centrifuged, aliquoted in 1 mL polypropylene tubes and stored (on average within 1.5 h of sampling) at –80°C until analysis. The specimens were thawed just before measurements.
Determination of tau oligomers and conformers by ELISA assay
Tau oligomers/conformers were measured by the semi-quantitative indirect ELISA assay using specific antibody T22/TTC-99, respectively. The wells of Polysorp NUNC microplates (Thermo Fisher) were coated overnight at 4°C with 20 μl of 1 : 20 diluted subject’s sera using 0.05 M bicarbonate buffer pH 9.5. The plates were washed one time with 1×Tris-buffered saline with 0.1% Tween-20, pH 7.4 (TBST) and blocked with 10% nonfat dried milk in TBST for 2 h at room temperature. Blocked plates were washed one time and treated with T22 (1 : 100) or TTC-99 (1 : 100) diluted in 5% nonfat milk in TBST for 1 h at room temperature. Subsequently, the plates were washed three times and incubated with 100 μl of horseradish peroxidase (HRP)-conjugated anti-rabbit IgG (GE Healthcare) diluted 1 : 3,000 in 5% milk TBST for 1 h at room temperature. Finally, plates were washed three times with TBST and incubated with 100 μl TMB+ substrate chromogen (Dako) for 6 min at room temperature in the dark. The reaction was stopped by 100 μl of 2 M HCl, and the plates were read at 450 nm in a POLARstar Omega plate reader (BMG Labtech). In vitro prepared tau oligomers [38] were included on each plate as a positive control and used for generation of the standard curve (Fig. 1-A,B). However, the precise determination of oligomer concentration was difficult due to variations of dye binding. Therefore, the ELISA is indicated as semi-quantitative with arbitrary units (AU/ml). The interassay variation between plates was calculated from 18 measurements for each antibody. The variation was 10% and 13% for T22 and TTC-99 antibody, respectively. All samples were measured in duplicates. Negative control (primary and secondary antibodies added to non-coated blocked wells) was measured on each plate. Randomly selected samples from each group of participants were probed only with secondary antibody to control non-specific signal. The non-specific signal was below the limit of detection (O.D. 0.107).
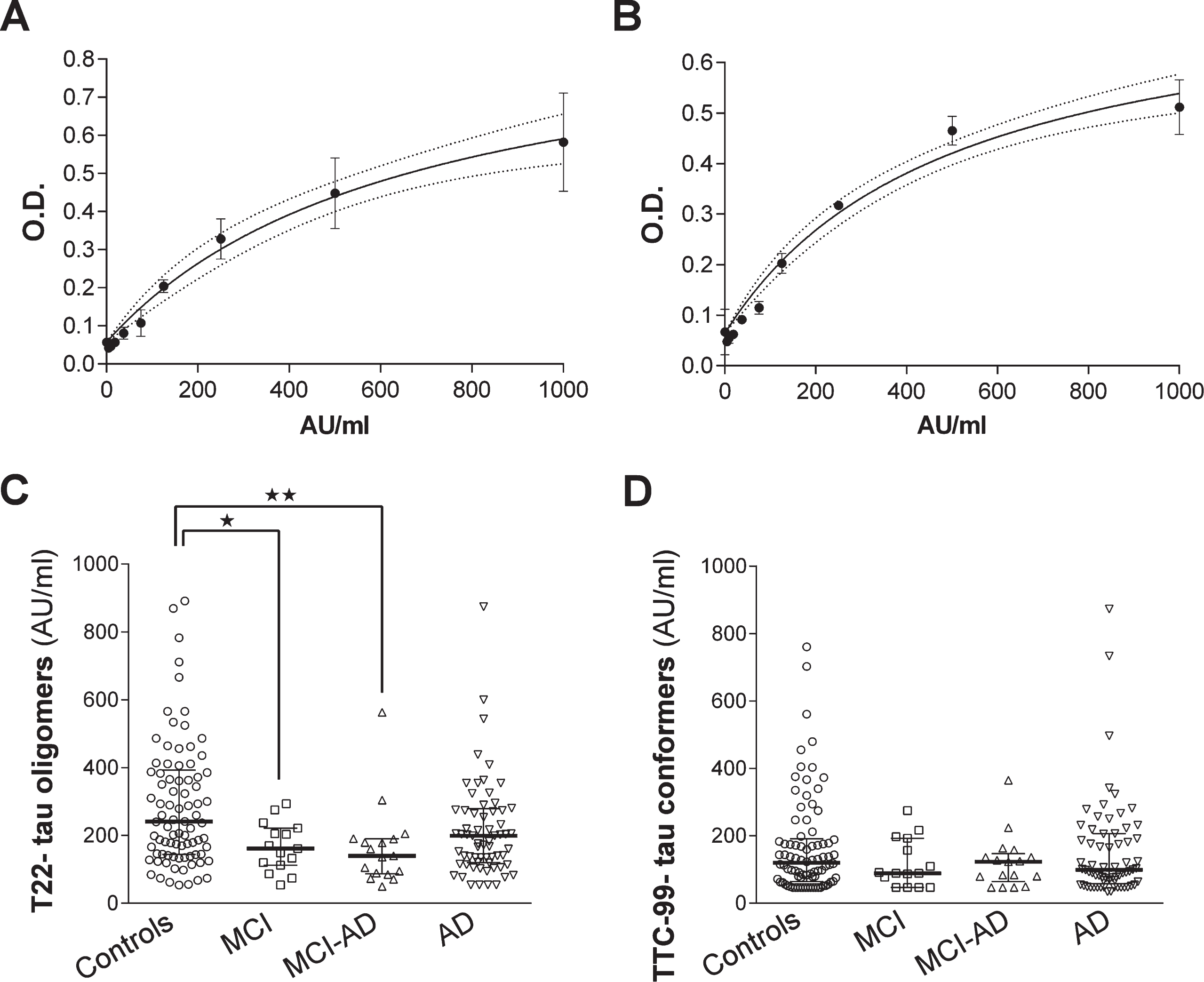
Serum levels of tau oligomers/conformers measured by ELISA. The standard curves of T22 (A) or TTC-99 (B) reactivity to recombinant tau oligomers were used for interpolation of tau oligomers/conformers concentrations in serum samples. Data are presented as a scatter plot of interpolated concentrations of T22- reactive tau oligomers (C) and TTC-99- reactive tau conformers (D). Concentrations are presented as medians with 25th-75th percentiles. Statistical significance was calculated with respect to controls and age as a covariate (Generalized linear model analysis with the covariate of age and contrast statement or Mann-Whitney test. *p < 0.05, **p < 0.01,***p < 0.001).
ELISA assays for CSF biomarkers of AD
The concentrations of recent CSF biomarkers of AD [total tau protein, phosphorylated tau at Thr 181 (phospho-tau181) and amyloid-β 1–42 peptide (Aβ42)] were measured by Fujirebio ELISA kit for some of the participants according to the manufacturer’s instructions.
Western blot analysis of sera samples
Western blot analysis of randomly selected sera samples was carried out to see the distribution of tau forms detected by antibody T22. Samples were prepared under non-reducing conditions in Novex NuPAGE LDS sample buffer (Invitrogen). 50 μg of total protein per lane was loaded on precast NuPAGE 10% Bis-Tris Gels for SDS-PAGE electrophoresis (Invitrogen) and subsequently transferred onto nitrocellulose membrane. After blocking overnight at 4°C with 10% nonfat dried milk in TBST, membranes were probed for 1 h at room temperature with T22 (1 : 150) and anti-albumin antibody (1 : 2,000; Bethyl) diluted with 5% nonfat dried milk. T22 was detected with HRP-conjugated anti-rabbit IgG (1 : 3,000; GE Healthcare) and anti-albumin antibody with HRP-conjugated IgG anti-goat secondary antibody (1 : 3,000; Southern Biotech). ECL Plus (Pierce) was used for signal generation. For protein quantification, the densitometry of each band was measured using ImageJ software and normalized with albumin. All densitometry results represent the mean±SD.
Statistical analysis
Data were analyzed with GraphPad Prism 6 (GraphPad Software) and STATISTICA software (StatSoft). Nonparametric Kruskal-Wallis test for global and Mann-Whitney test for pairwise comparisons were calculated for group characteristics and results of tau conformers obtained by ELISA. Generalized linear model (GLM) analysis with the covariate of age and contrast statement for intergroup comparisons was performed for analyzing results of logarithmically transformed tau oligomers obtained by ELISA with T22 antibody. Correlations coefficients were calculated using a Spearman correlation. Statistical analysis of Western blot quantification results was performed using the two-tailed Student’s t-test. The results are presented as the mean±SD for clinical variables of groups and median with interquartile range for results of ELISA assays. A value of p < 0.05 was consideredsignificant.
RESULTS
By ELISA assays, we measured tau oligomers/conformers in 186 sera samples of subjects divided into groups of patients with different stages of cognitive impairment and one group of healthy subjects. Table 1 shows demographic and cognitive characteristics of individuals in groups including measured levels of current CSF biomarkers (total tau, phospho-tau181, and Aβ42) by commercial ELISA kits if they were available. Statistical analysis revealed expected significant drops in MMSE score in all groups with cognitive impairment when compared to controls. The concentration of total tau was higher in groups of patients with dementia due to AD. Levels of a phospho-tau181 protein were higher in cognitively impaired groups in contrast to controls. Aβ42 levels showed significant difference only between the groups of patients with dementia due to AD and cognitively healthy subjects. By comparing the average age of subjects in groups, we found that individuals in MCI-AD (Kruskal-Wallis test, p < 0.001) and dementia due to AD (Kruskal-Wallis test, p < 0.001) groups were older than cognitively normal subjects. Therefore, we analyzed if age is an important variable in the analysis of levels of tau oligomers/conformers. After we interpolated the levels of tau oligomers/conformers from calibration curves (Fig. 1A, B), we found that the age of participants correlated with the serum tau oligomers (GLM, p = 0.005) levels, but not serum tau conformers (GLM, p = 0.167). The statistical comparison between groups for serum tau oligomers was, therefore, calculated with respect to age as a covariate. The levels of serum tau oligomers were decreased in MCI group (p = 0.033) and MCI-AD group (p = 0.006) with respect to healthy subjects (Fig. 1C). In sera of patients with dementia due to AD, we observed an elevation of tau oligomers levels in comparison to MCI and MCI-AD groups, but it did not reach statistical significance. Tau conformers did not show any difference between groups (Kruskal-Wallis test, p = 0.486, Fig. 1D). We performed a correlation analysis of serum T22- tau oligomers with basic groups’ characteristics and TTC-99- tau conformers (Table 2). The results of the correlation analysis indicated a positive correlation between aging and levels of tau oligomers in the group of controls suggesting increasing levels of tau oligomers with aging. We have found strong positive links between tau oligomer and tau conformer levels measured by ELISA assays in all groups except MCI group.
Demographic, cognitive, and cerebrospinal fluid characteristics of subjects enrolled for measurement of tau oligomers
Data are presented as the mean±SD. Statistically significant differences highlighted in bold were calculated with respect to controls (Mann-Whitney test, **p < 0.01, ***p < 0.001). Data of CSF biomarkers (total tau, phospho-tau181, and Aβ42) were obtained from 40 controls, 11 MCI subjects, 17 patients with MCI due to AD, and 45 AD patients. MCI, mild cognitive impairment; AD, Alzheimer’s disease; MMSE, Mini-Mental State Examination.
Correlation analysis of serum T22- tau oligomers levels with cognitive and CSF characteristics and TTC-99- tau conformers levels
Data were collected from 186 subjects (86 non-demented controls, 14 individuals with mild cognitive impairment (MCI), 18 patients with mild cognitive impairment due to AD (MCI-AD), 68 AD patients (AD). Data of CSF biomarkers (total tau, phospho-tau181, and Aβ42) were obtained from 40 controls, 11 MCI subjects, 17 patients with MCI due to AD and 45 AD patients. Correlation coeficients (CC) were evaluated via a Spearman correlation for non-normally distributed data. Finally, permutation analysis was performed to compare groups.
Distribution of T22 reactive proteins in sera samples assayed by western blot
To visualize tau oligomers in human serum that react with the T22 antibody, western blot analysis of serum samples from three patients with dementia due to AD and three controls was performed (Fig. 2A). T22- immunoreactive tau oligomers higher than 250 kDa were observed in the serum samples of patients with dementia due to AD but also in some of the control samples. To ensure the same amount of protein from each sample was loaded into each lane, we used anti-albumin antibody as the loading control (Fig. 2A). Densitometric quantifications of higher molecular weight (HMW) bands (>250 kDa) normalized to albumin revealed significantly increased tau oligomers in patients with dementia due to AD (p = 0.012, Student’s t-test) (Fig. 2B). When the density of all protein bands was compared between groups, the patients with dementia due to AD had significantly more tau oligomers detected with T22 when compared to controls (p = 0.038, Student’s t-test) (Fig. 2C). We attribute the bands around 120–150 kDa to the tau dimers/trimers and non-specific immunoglobulins.
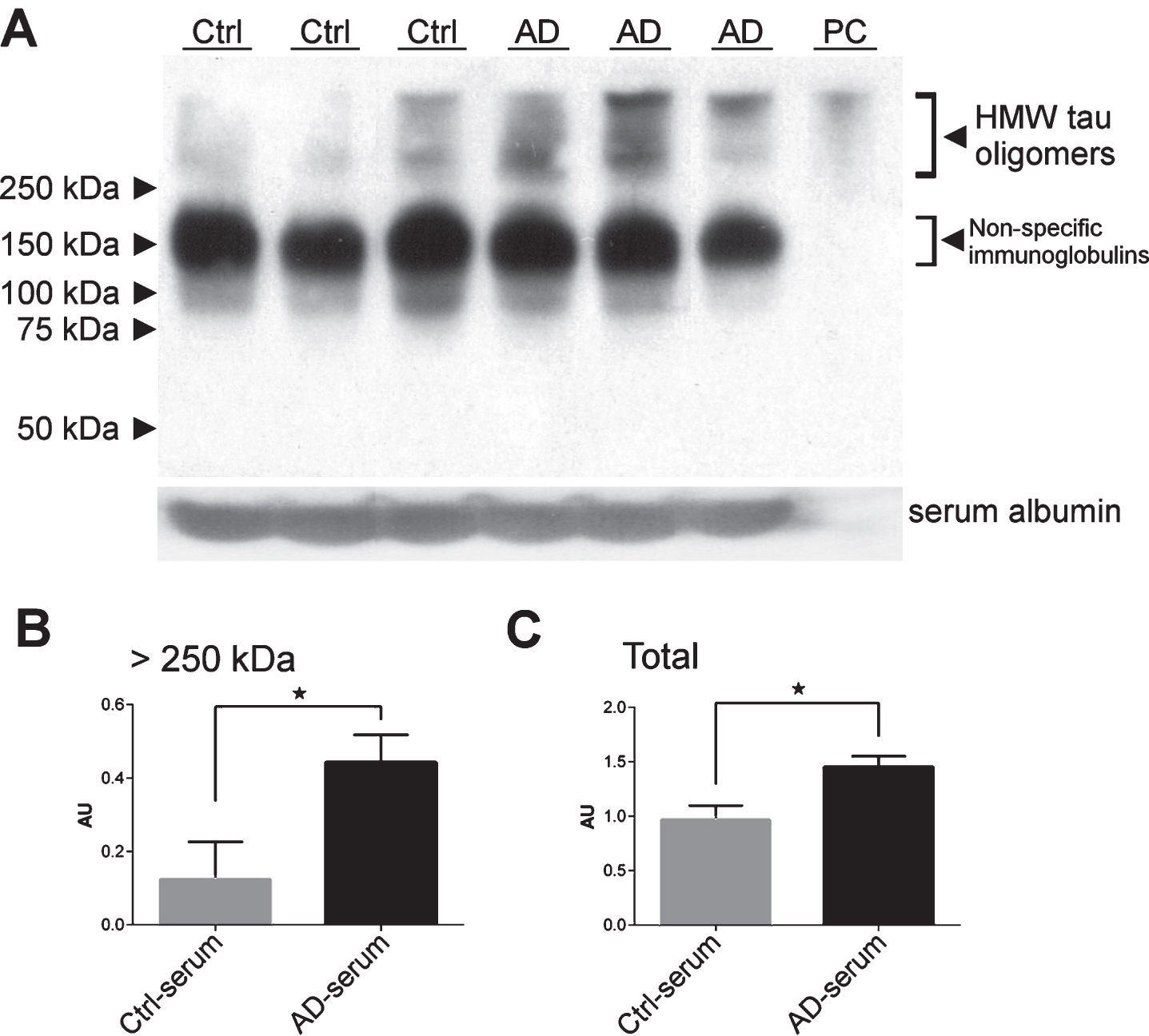
Western blot analysis of serum samples (50 μg of total protein/lane) from three patients with dementia due to Alzheimer’s disease (AD) and three cognitively normal controls (Ctrl) using rabbit polyclonal anti-oligomer antibody T22. In vitro prepared recombinant tau oligomers were used as a positive control (PC). Detection of serum albumin in each sample was used as a loading control (A). Tau oligomers detected by antibody T22 were quantified by densitometry in the higher molecular weight (HMW) bands (B) and the whole lane (C). Densitometric quantifications (AU, arbitrary unit) were normalized to albumin. Bars represent mean±SD.
DISCUSSION
There are studies concerning the occurrence of tau protein and its modified forms in CSF because this fluid is in direct contact with the brain parenchyma. Lower CSF Aβ42 levels and elevated total tau, and phospho-tau181 are considered as the core markers of AD pathology. Measurement of other CSF phospho tau like p-tau 231–235,or p-tau 396–404 have also been shown in AD diagnosis [18, 19]. However, detection of tau protein forms circulating in the blood could be a valuable diagnostic marker of AD. Several studies have reported plasma and serum levels of tau protein, but the results are contradictory. While one study showed lowered levels of plasmatic tau proteins in AD patients [39], Zetterberg et al. on the contrary, found higher plasma levels of tau protein in AD patients using their digital array technology [40]. The serum levels of tau protein were reported to be elevated after mild traumatic brain injury (TBI) [41] and in acute ischemic stroke [42]. Recent studies reported that release of endogenous tau protein from neurons into the interstitial fluid (ISF) is a physiological process which is mediated by neuronal activity [43, 44]. They suggest that released tau proteins can function as a signal for adjacent cells and may be taken up by post-synaptic neurons. However, the basal levels of tau have to be restored upon release. The secreted tau could be washed out of ISF through glymphatic pathway into the blood where it can be subjected to fast degradation by proteases or targeted by antibodies.
In our study, we detected T22- reactive tau oligomers in the human serum, interestingly their levels positively correlate with aging. However, the exact cause of this aggregation remains unidentified. It is reported that metabolism slows down and coagulation activity in blood increases with aging [45]. Therefore, the increased tau oligomer burden detected in the serum of patients with dementia due to AD could be a consequence of the higher accumulation of tau, thrombin and clotting agents in the blood. The observation of tau oligomers detected in the serum was also supported by western blot assay. The detection of HMW tau oligomers in serum of AD patients and to some extent in control individuals in our study is in agreement with the study of Neumann et al. and Farías et al. where the researchers observed similar HMW forms of tau protein in the platelets of both AD patients and control subjects [46, 47].
The impaired clearance of tau protein from the ISF through glymphatic pathway appearing after TBI has recently been described in a TBI mouse model [48], which is an established risk factor for the development of neurodegeneration, including AD. In addition, the physiological efflux of proteins from CSF to blood through the choroid plexus was described as a clearance pathway for Aβ peptide [21, 50]. No studies describing clearance of tau protein through this barrier have been published thus far [50]. However, pathological changes in the choroid plexus related to neurodegeneration were described as enhancing tau oligomerization [23, 52] and that may be reflected later on in the course of the disease as an increase of stable tau oligomers in blood. On the other hand, the accumulation of HMW oligomers could correlate to the pathophysiology of the disease in both the CNS and the peripheral blood cells. The results obtained in this study will need to be validated with larger sample size. Although, the mechanisms involved in the clearance of tau are not completely understood when compared to Aβ clearance, the changes in the levels of tau oligomers with the manifestations of cognitive decline could be a valuable tool for future diagnostics. Additionally, similar to fibrillar structures, tau oligomers can exhibit different conformers whose aggregation stage might reflect the state of the disease. A recent study has demonstrated that Aβ oligomers from AD brain are characteristically different than those collected from CSF in their seeding property [53]. Therefore, extending our study to better analyze these tau oligomeric species will enhance our existing understanding of the heterogeneity of different tau oligomer conformers from different sources like brain, CSF and blood serum/plasma. Thus, further research on heterogeneous tau oligomer population will provide more insight into the understanding of tau trafficking and its role in AD.
Footnotes
ACKNOWLEDGMENTS
This study was supported by the grants from National Institute of Health RO1AG054025, RO1NS094557 project Nr. LO1611 with a financial support from the MEYS under the NPU I program, GACR 13-26601S, PROGRES Q35, The Alzheimer Foundation Czech Republic (AVASTipendium for the human brain), grant Mobility of Charles University in Prague and the Mitchell Center for Neurodegenerative Diseases.