
Abstract
Alzheimer’s disease (AD) and progressive supranuclear palsy are two common neurodegenerative tauopathies, and the most common cause of progressive brain dementia in elderly affecting more than 35 million people. The tauopathies are characterized by abnormal deposition of microtubule associated protein tau into intracellular neurofibrillary tangles composed mainly of the hyperphosphorylated form of the protein. The diagnosis of tauopathies is based on the presence of clinical features and pathological changes. Over the last decade, there has been an intensive search for novel biochemical markers for clinical diagnosis of AD and other tauopathies. In the present study, we used transgenic rat model for tauopathy expressing human truncated tau protein (aa 151–391/4R) to analyze the cerebrospinal fluid (CSF) peptidome using liquid chromatography – matrix assisted laser desorption/ionization mass spectrometry (LC-MALDI TOF/TOF). From 345 peptides, we identified a total of 175 proteins. Among them, 17 proteins were significantly altered in the CSF of transgenic rats. The following proteins were elevated in the CSF of transgenic rats when compared to the control animals: neurofilament light and medium chain, apolipoprotein E, gamma-synuclein, chromogranin A, reticulon-4, secretogranin-2, calsyntein-1 and -3, endothelin-3, neuroendocrine protein B72A, alpha-1-macroglobulin, and augurin. Interestingly most of the identified proteins were previously linked to AD and other tauopathies, indicating the significance of transgenic animals in biomarker validation.
INTRODUCTION
The lack of specific and sensitive diagnostic tests for many human and animal diseases raises the demand for new biomarkers that provide a dynamic view of the health status of patients in different stages or types of a disease [1, 2]. New biomarkers may not only be useful for the diagnosis of the disease but also for toxicological and drug development platforms [2]. Bioactive peptides have been studied for more than one hundred years; however, most studies were predominantly focused on a specific peptide or peptides, and not on the whole peptidome. Following the development of protein analysis using mass spectrometry techniques in the 1990 s, the peptidomic analysis gained popularity.
Peptidomics is an emerging field derived from proteomics, which deals with the qualitative and quantitative analysis of endogenously produced peptides present in biological samples [3], which may be either beneficial or harmful. Identifying potential biomarkers is one of the most frequent applications of peptidomics. Peptidomics is widely used in the search for biomarkers for cancer [4], cardiovascular diseases [5], acute/chronic rejection and operational tolerance in organ transplantations [6], allergy and asthma [7], chronic kidney disease [8], diabetes, osteoporosis [2], and male fertility studies [9], and for biomarkers in neurodegeneration diseases includingtauopathies [10].
Tauopathies represents a group of about 20 diseases characterized by deposition of misfolded protein tau. There is no disease modifying therapy in the market, and the development is limited due to the lack of sensitive biomarkers for the diseases. Cerebrospinal fluid (CSF) occupies the subarachnoid space in the ventricular system around the brain and lacks a tightly regulated barrier with the brain. When compared to plasma or urine, the CSF is the most informative fluid for diagnosis and prognosis of neurodegenerative disorders [11]. The proteins, peptides, and metabolites found in CSF reflect brain activities and could therefore serve as good biomarkers or surrogate markers forneurodegeneration.
Here we present the data from the CSF peptidome study conducted on the transgenic rat model for tauopathies expressing human truncated tau line SHR72 (aa 151–391/4R). We identified altered levels of several peptides and proteins in CSF. The identified proteins are linked to different biological processes and pathways, and reflect tau induced neurodegeneration in the brain. To our knowledge, this study is the first to investigate the peptidome of CSF in a tau transgenic model.
MATERIALS AND METHODS
Chemicals and reagents
α-cyano-4-hydroxycinammic acid (HCCA, MALDI grade, part no. 201344) and MALDI mass calibrant (Peptide mass calibration standard II, part no. 222570) were purchased from Bruker Daltonics (Brno, Czech Republic). LC/MS grade acetonitrile, formic acid, water, and trifluoroacetic acid were obtained from Sigma Aldrich (Lambda Life, Bratislava, Slovakia). Deionized water used in this work was prepared by the water purification system Milli-Q (Merck Millipore, Bratislava, Slovakia). MWCO (Microcon 0.5 mL, 30 Kda) filters were purchased from Merck Millipore (Merck Millipore, Bratislava, Slovakia).
Animals
The generation and characterization of a transgenic rat model for tauopathy expressing human truncated tau (aa 151–391/4R; line SHR72) is described elsewhere [12]. For this study, we used heterozygous transgenic rats (5–7 months old; n = 13) and non-transgenic SHR age-matched controls (n = 15). All animals were housed under standard laboratory conditions with free access to water and food, and were kept under diurnal lighting conditions (12 h light/dark cycles with light starting at 7:00 a.m.). All experiments on animals were performed in accordance to the institutional animal care guidelines conforming to international standards (ARRIVE guidelines) and were approved by the State Veterinary and Food Committee of Slovak Republic (#Ro-1101/14-221C) and by the Ethics Committee of Institute of Neuroimmunology. Efforts were made to minimize the number of animals utilized and to limit discomfort, pain, or any other suffering of the experimental animals used in this study.
Collection of cerebrospinal fluid
CSF was collected from the cisterna magna. Animals were anesthetized with tyletamin/zolazepam/xylazine mixture, fixed in a head holder, and a midline incision in the skin was made up to the head area to permit easy access to the cisterna magna. Approximately 80 μl of CSF was collected from each animal. After a brief centrifugation (3 min at 5000× g, 4°C), all CSF samples were immediately flash frozen in liquid nitrogen and stored at –80°C until used.
Sample preparation
To 80 μL of CSF, acetonitrile was added (20% final v/v) and samples were centrifuged at 4°C for 10 min (20,000 × g). Then the CSF samples were centrifuged through the MWCO filters according to the manufacturer’s instructions. All steps were performed on ice without addition of protease inhibitors. The filtrate containing the peptide fraction was dried using the SpeedVac concentrator (Pragolab, Bratislava, Slovakia) and then stored at –80°C until analyzed.
Liquid chromatography
The Ultimate 3000 nano-LC system (Dionex, Bruker Daltonics Brno, Czech Republic) with Acclaim PepMap RSLC C18 column (75 mm, 150 mm, Thermo Scientific) and Acclaim PepMap100 C18 trap column (75 mm, 20 mm, Thermo Scientific) was used for peptide separation. Mobile phase A consisted of trifluoroacetic acid (Merck, Bratislava, Slovakia) in water (0.05% v/v). Mobile phase B consisted of 80% acetonitrile with 0.05% trifluoroacetic acid (v/v). The elution started at 10% B (0–15 min), increasing to 70% B (15–79 min), to 95% B (80–100 min), returning to 10% B and re-equilibrating at 100 to 120 min. The column temperature was 40°C. The flow rate was 0.3 μL/min and the injection volume was eight μL. A liquid dispensing robot Proteineer fc II (Bruker Daltonics, Brno, Czech Republic) was connected to the nano-LC device and was used to dispense fraction from the separation onto a MTP Anchor Chip 384 MALDI sample plate (Bruker Daltonics, Brno Czech Republic). The effluent from the nano-LC was mixed with HCCA matrix solution and dispensed onto the target plate within 192 spots.
Mass spectrometry
The fractionated samples were analyzed using a MALDI TOF/TOF (Ultraflextreme, Bruker Daltonics, Brno, Czech Republic) instrument operated in the positive ion mode with mass range 700–5000 and deflection up to m/z 600. The analysis of the MS spectra and selection of compounds for MS/MS analysis was performed using the WARP-LC software (Bruker Daltonics, Brno, Czech Republic). Only compounds with S/N >12 were selected for MS/MS analysis. 4,000 single shot spectra were accumulated from 40 random positions on each sample, irradiating each position with 100 laser pulses. For MS/MS analysis, 4,000 single shot spectra were recorded for the fragment ions.
Data processing and statistics
Statistical analysis was performed in R software [13]. The absolute and relative frequencies of protein prevalence per animal were calculated. The null hypotheses about difference in relative prevalence of each protein between SHR72 and SHR (null hypothesis: the difference is equal to zero against alternative hypothesis: the difference is not equal to zero) were tested by two-sample Score test [14] at a significance level α equal to 0.05. Related differences and empirical 95% score confidence intervals were calculated also. Differentially abundant proteins were subjected to GO classification via the Panther Classification System Database [15] to investigate biological processes, molecular function, and cellular compartment.
RESULTS
CSF sampling and sample preparation
Cisterna magna puncture is a highly effective method in rats that allow sampling of up to 100 μl of CSF [16]. Only CSF samples that contained no visible red blood cell pellet were selected for peptidomic analysis. We used simple sample preparation procedure that consisted of dilution with acetonitrile and ultrafiltration through centrifugal filter units, as described previously [17]. Dilution with acetonitrile is important since many peptides may attach to plastic material and their recovery would be very low. Diluted samples were separated by LC and fractions were spotted onto MALDI targets and analyzed. Figure 1 presents a workflow used in this study.
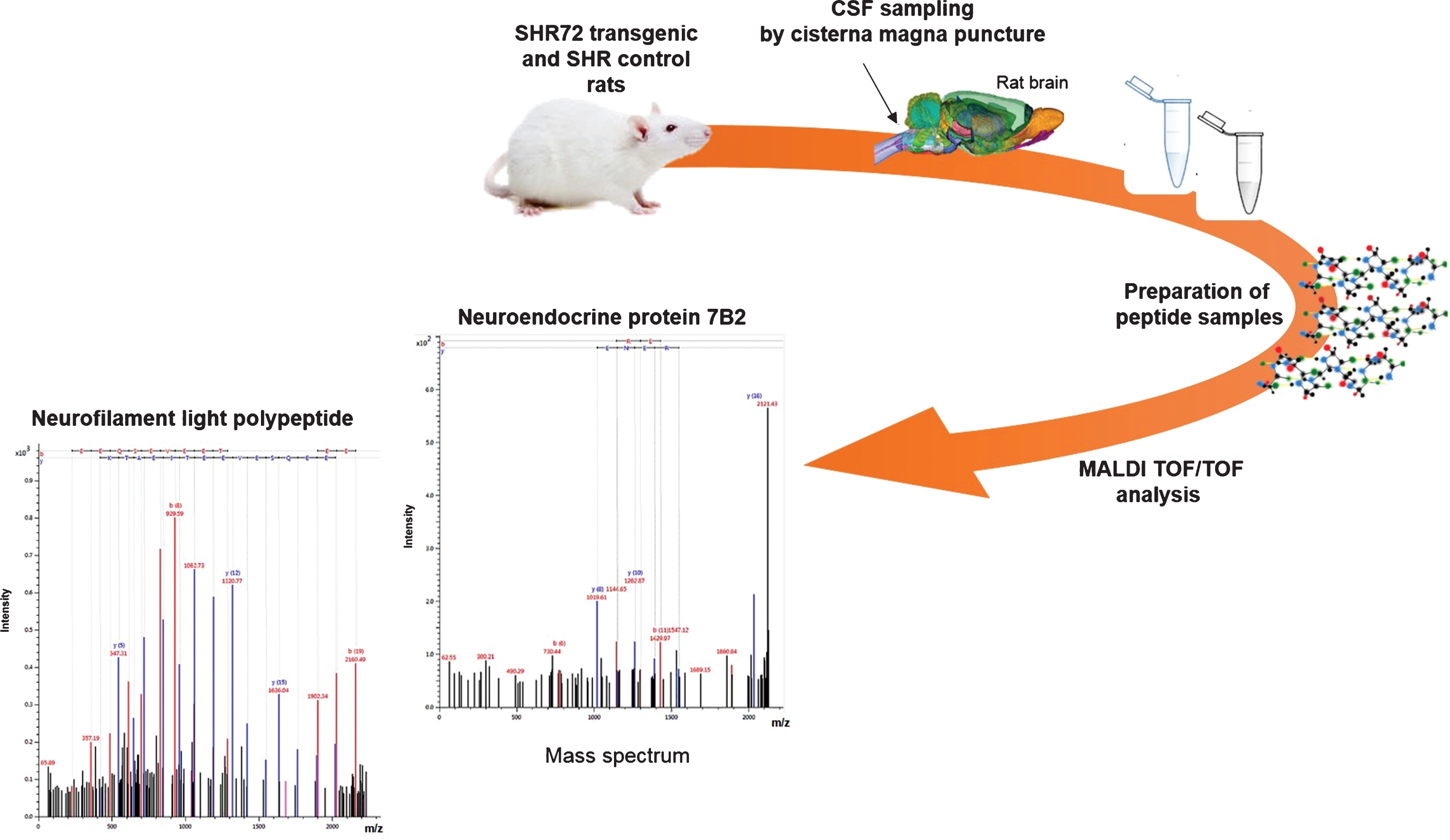
Schematic outline of a workflow used in our study. CSF was collected from cisterna magna. Sample pretreatment steps included dilution and ultrafiltration. Prepared samples were separated by nano-LC and fractions were spotted onto MALDI targets together with matrix. Mass spectra were generated for each fraction and tandem MS (MS/MS) spectra were generated for peptide/protein identification. MALDI, matrix-assisted laser desorption ionization; TOF, time-of-flight, MS, mass spectrometry; m/z, mass to charge ratio, SHR, spontaneously hypertensive rat.
LC-MALDI and peptide identification
For optimal separation of peptides, the gradient elution program was established. Figure 2 displays typical chromatogram and LC-MS profile of rat CSF peptidome. We detected 325 peptides in CSF from control SHR animals (mean; range 325±170; n = 13), and we obtained a total of 491 hits in transgenic SHR72 animals (mean; range 491±172; n = 15). Subsequently, a total of 175 proteins derived from 345 peptides were identified using Mascot search.
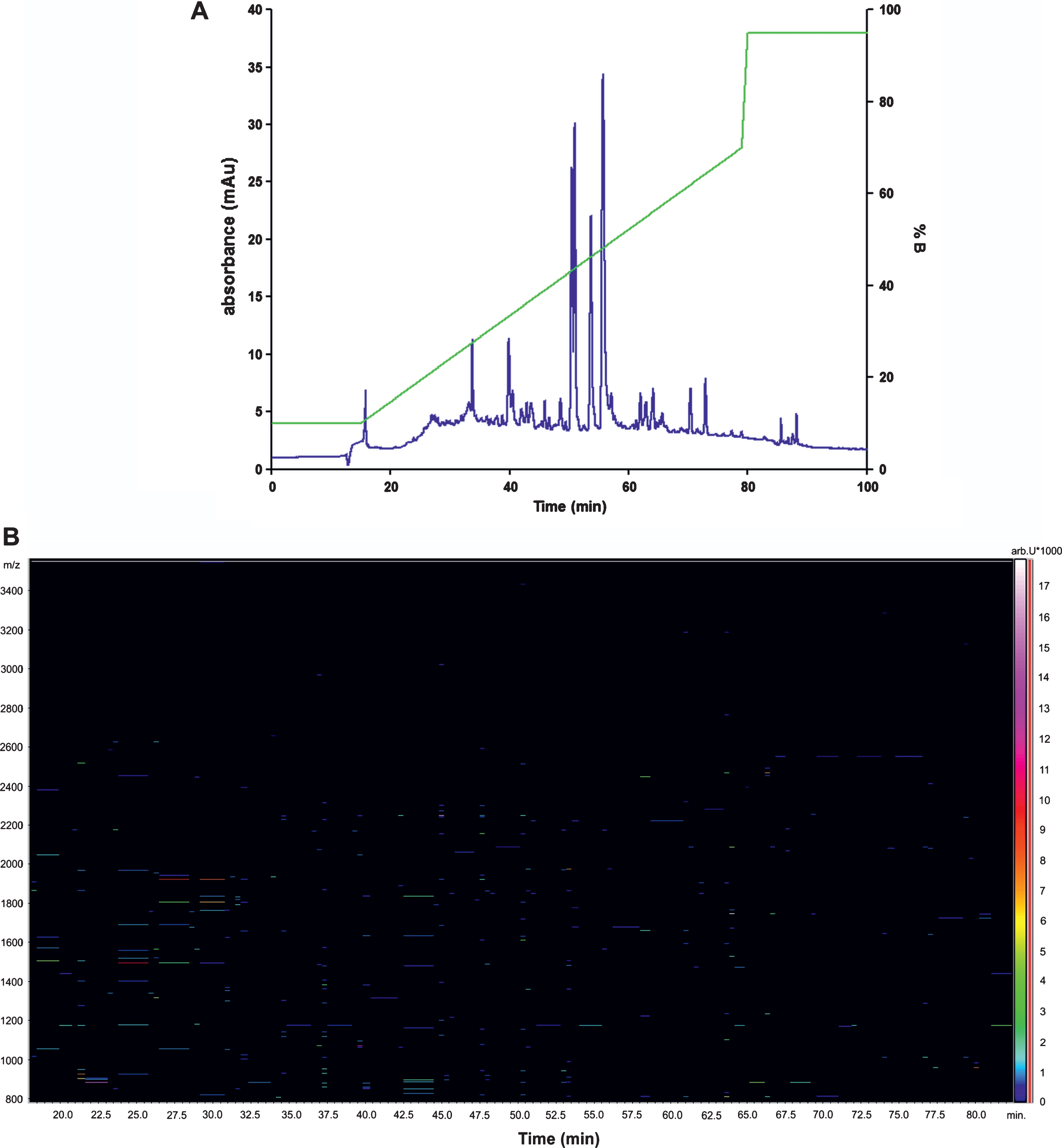
Representative chromatogram and LC-MS profile of rat CSF peptides. A) LC/UV chromatographic profile. Compounds were detected using absorption at 214 nm. Retention time is shown on the x-axis and gradient profile (% B) is shown on right y-axis. B) The LC-MS profile. Retention time is shown on the x-axis and the m/z on the y-axis. m/z, mass to charge ratio.
The results from the first 20 proteins are summarized in Table 1. A table of all identified peptides and proteins is provided in Supplementary Table 1. 17 out of 20 proteins showed significantly altered levels (p < 0.05) in the CSF from transgenic SHR72 animals, when compared to controls.
The list of identified peptides and proteins
amolecular weight in kDa. bincreased or decreased in SHR72 transgenic animals compare to SHR controls. NC, no significant change. ↑Increase.
Our data showed increased levels of neurofilament light and medium polypeptides (Nfs, p = 0.0007; p = 0.0443), alpha-1-macroglobulin (p = 0.0443), apolipoprotein E (ApoE, p = 0.0443), augurin (p = 0.0443), calsyntenin-1 and -3 (p = 0.0216; p = 0.0443), endothelin-3 (p = 0.0216), fibrinogen beta chain (p = 0.049), gamma-synuclein (p = 0.0216), neuroendocrine protein 7B2 (p = 0.008), small nuclear ribonucleoprotein-associated protein N (p = 0.0443), prosaposin receptor (p = 0.0443), receptor-type tyrosine-protein phosphatase zeta (PTPRZ1) (p = 0.0443), reticulon-4 (p = 0.0045), chromogranin-A (p = 0.0101), and secretogranin-2 (p = 0.049).
For Nf light polypeptide, we identified three peptides from C-terminal region of the protein (aa 439–467). For Nf medium polypeptide, we also identified two peptides from C-terminus (aa 441–466). For ApoE, we identified two peptides from N-terminus (aa 19–43). Two peptides from C-terminus of calsyntenin-1 (aa 819–832), and nine peptides corresponding to the mid regions of calsyntenin-3 (aa 156–180) were identified. For other proteins such as gamma-synuclein, we identified two peptides from C-terminal part (aa 94–123) and three N-terminal peptides for endothelin-3 (aa 19–52). Two peptides, both from middle region (aa 42–68), were identified for augurin. For chromogranin-A, we found two peptides, one from C-terminus (aa 438–463) and one from middle region of the protein (aa 342–353). For reticulon-4, we found one peptide from N-terminal of the protein (aa 135–145). Twelve peptides from C-terminal part were identified for neuroendocrine protein 7B2 (aa 180–210). We found three peptides (aa 1198–1204; aa 1625–1648) from mid region of PTPRZ1 and two peptides from N-terminal part of fibrinogen beta chain (aa 18–32). Prosaposin receptor was represented by one peptide from N-terminal part of the protein (aa 53–78) and small nuclear ribonucleoprotein-associated protein N by one peptide from C-terminal region (aa 185–193). Finally, we found twelve peptides from N-terminal (aa 184–216) and C-terminus of secretogranin-2 (aa 493–612).
Analysis of biological processes and functions
Differentially abundant proteins were subjected to GO classification via the Panther Classification System Database to investigate biological processes, molecular functions and cellular compartments (Fig. 3). Proteins significantly enriched in transgenic animals were primarily related to biosynthetic process, nervous system development, protein metabolic processes and system development (Fig. 3A). The predominant molecular functions of proteins found in CSF of transgenic animals included motor activity, peptidase and peptidase inhibitor activity, and receptor activity (Fig. 3B). Moreover, in terms of cellular compartment, transgenic rats-enriched proteins were comprised of a substantially higher percentage ofG-protein complexes, cytosolic, intermediate filament cytoskeleton, extracellular region and space than proteins in control animals (Fig. 3C).
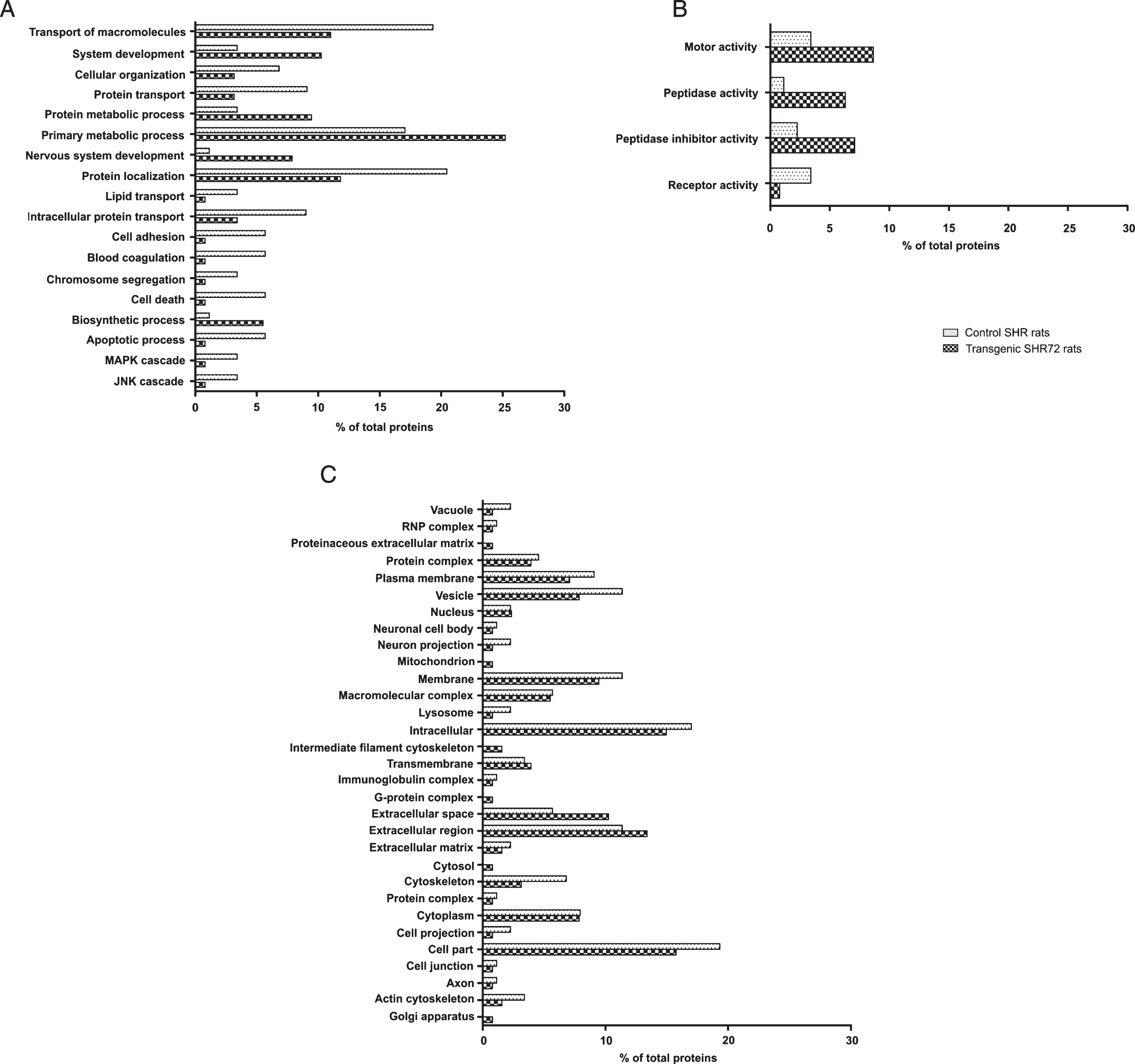
GO analysis of protein differing between control and transgenic rats. Proteins with significant differences between control and transgenic animals were subjected to GO analysis in terms of biological process (A), molecular function (B), and cellular compartment (C).
DISCUSSION
Identification and evaluation of CSF biomarkers in animal models can assist in development and implementation of similar approaches in clinical population of humans. The biomarkers identified in animal studies may also be invaluable in monitoring efficacy of therapeutic compounds in clinical trials. Several genetically modified rodent models with clinical, histopathological, and molecular characteristics of neurodegeneration have been developed. In the present work, we used SHR72 transgenic rat model for tauopathy with progressive age-dependent neurofibrillary degeneration in brainstem, deep cerebellar nuclei, and spinal cord. The neurofibrillary pathology was induced by structurally modified truncated tau protein consisting of four microtubule binding domains (4R) and the proline-rich region (151–391/4R). The model shows many characteristics of tauopathies, such as tau hyperphosphorylation, formation of neurofibrillary tangles, oxidative stress, and neuroinflammation [12, 19].
CSF as a major biological fluid used in human neuroscience research can be easily collected from rats by cisterna magna puncture. Interestingly, CSF can be repeatedly sampled which allows periodic monitoring and evaluation of a disease. In the current study, we analyzed and compared the CSF peptidome profiles of non-transgenic SHR rats and SHR72 transgenic animals. We detected more than 300 peptides derived from 175 proteins. In physiological conditions the protease activity in CSF is low [20]. It is known that during central nervous system (CNS) infections, multiple sclerosis, traumatic brain injury, and Alzheimer’s disease (AD), the protease activity in CSF is increased [21–23]. Therefore, some peptides we detected might be as a result of increased protease activity in CSF of tau transgenic animals. However, the presence of these proteins in CSF suggests neuronal damage and the potential use of the CSF peptides as biomarkers of thedisease.
We identified 17 proteins that are significantly elevated in the CSF of transgenic versus control animals, many of which were previously linked to neurodegeneration in AD and related disorders. Table 2 summarizes the identified proteins based on their biological functions and involvement in the pathogenesis of diseases in the CNS.
Potential biomarkers for tauopathies
AD, Alzheimer’s disease; PD, Parkinson’s disease; ALS, amyotrophic lateral sclerosis; CNS, central nervous system.
The Nf light and medium polypeptides were significantly higher in CSF from transgenic animals compared to the controls. Nfs are neuron-specific intermediate filaments that are composed of four subunits: Nf light, Nf medium, and Nf heavy chain and alpha-internexin. Nfs are assembled with cytoskeletal proteins, transported into axonal compartment, where they are extensively phosphorylated [24]. Phosphorylation regulates their axonal transport, dynamics, and proteolysis [10, 26].
In addition, Nfs as components of AD neurofibrillary pathology are abnormally hyperphosphorylated [27–29]. Nfs are released following axonal damage and neuronal degeneration. The presence of Nfs in the CSF [29–32] and blood has been suggested as a biomarker useful for prediction and monitoring of disease progression in AD and related tauopathies [33, 34]. In our model, the increase of Nfs in CSF is most likely due to the progressive tau induced axonopathy.
Apolipoprotein E (ApoE) has been studied as a potential biomarker in AD, but the levels of ApoE has not been considered diagnostically useful. In the peripheral system, ApoE is involved in the transportation of triglycerides, phospholipids, and cholesterol [35]. Recent studies on ApoE have revealed conflicting results with some groups reporting lower levels of ApoE in CSF from AD patients [36, 37], while others showing increased levels [38, 39]. We found increased levels of ApoE in CSF of transgenic animals. Significantly higher levels of ApoE in CSF reflect neuronal damage and inflammation that is predominant during AD pathogenesis [40] and is one of the characteristic features of the transgenic model.
Peptides from the granins have been discussed as CSF biomarkers for diseases such as AD, multiple sclerosis, and depression [41] The granins (Chromogranin A, Secretogranin-1, -2 and -3, neuroendocrine protein 7B2, proSAAS, and VGF) are involved in the delivery of several factors such as neurotransmitters, hormones and growth factors. The neuroendocrine protein 7B2 acts as a prohormone convertase 2-binding protein and is universally expressed in endocrine, neural and neuroendocrine cells [42]. Its widespread neuronal distribution within the brain, and also in the areas lacking prohormone convertases strongly suggests its physiological functions in neuropeptide synthesis [43, 44]. Previous studies have shown that 7B2 blocks the formation of proPC2 oligomers and aggregates [45], as well as IGF-1 aggregates [46], and demonstrated its function as a post-folding and post-secretion chaperone. In addition, different independent discovery studies identified 7B2 as a potential biomarker of early-onset of neurodegenerative diseases, such as AD, Parkinson’s disease, and amyotrophic lateral sclerosis [47–50].
Chromogranin-A is the major soluble protein in catecholamine storage vesicles and its abnormalities may play a central role in memory deficits in AD. Chromogranin-A was identified as a potential biochemical marker for the synaptic degeneration in AD, and a CSF biomarker for patients with multiple sclerosis [30, 51]. Secretogranin-2 belongs to the granin family of prohormones widely distributed in dense-core secretory granules of endocrine, neuroendocrine, and neuronal cells [52]. These acidic secretory proteins play a role in the morphogenesis of secretory vesicles and are precursor proteins for biologically active peptides. Secretogranin-2 is the precursor of secretoneurin, a peptide playing a role in the modulation of the inflammatory response, which may represent a novel sensory peptide for neuroimmunomodulation [53].
Receptor-type tyrosine-protein phosphatase zeta is a member of the protein tyrosine- phosphatase receptor family [54]. It plays a negative role in oligodendrocyte differentiation in early CNS development, and in remyelination of demyelinating CNS diseases, through the dephosphorylation of substrates such as p190RhoGAP [55]. In contrast to our results, analysis of the CSF proteome using an antibody-based detection method showed significantly lower levels of protein tyrosine- phosphatase receptor in AD patients when compared to non-dementedcontrols [56].
In the CSF of transgenic rats, we also identified significantly increased levels of Reticulon-4 (Nogo-A) that belongs to a super-family of proteins (reticulons) named after their principal subcellular localization at the endoplasmic reticulum [57]. Nogo-A is an important growth cone collapsing and growth inhibitory protein in the adult CNS. Its main role is the inhibition of axon regeneration and compensatory sprouting after CNS injury [58]. Different studies have highlighted the importance of reticulons in the pathogenesis of neurodegenerative diseases. Fluctuations of these proteins lead to the damage of normal brain functions in several neurodegenerative disorders, as well as in schizophrenia [57]. Recently, it has been shown that Nogo-A could also play an important role in AD pathogenesis through the modulation of the amyloid-β protein formation [59].
Gamma-synuclein is an aggregation-prone protein belonging to the synuclein family. It forms atypical intracellular inclusions in different human neurodegenerative diseases. Moreover, elevated expression of gamma-synuclein leads to selective damage and loss of discrete populations of neurons and their axons [60]. Gamma-synuclein also plays a role in neurofilament network integrity [61].
Few other proteins demonstrated a statistically significant elevation in transgenic animals in comparison to the control group (Table 1). Prosaposin was previously described as a potential CSF biomarker for AD neurodegeneration [39]. Prosaposin is a chaperone protein critical for lysosomal catabolism of glycosphingolipids. The increased level of prosaposin can be a result of generalized lysosomal dysfunction during neurodegeneration [62]. Lee et al. demonstrated an elevated level of fibrinogen gamma-A chain precursor protein in the CSF from patients with mild cognitive impairment and AD compared to the age-matched normal subjects [63]. Its expression was prominent in the AD group and correlated with disease severity and progression. However, it cannot be excluded that increased levels of fibrinogen in brain tissue may be due to increased blood levels and/or increased permeability across the blood-CSF barrier. Increase of alpha 1-macroglobulin in CSF could be explained mainly by an increase in permeability through damaged blood-brain barrier under pathological conditions.
Endothelins are endothelium-derived vasoactive peptides involved in variety of biological functions. Endothelins are present at low levels in normal human CSF; however, previous studies have suggested a possible alteration of the endothelin-1 system in the CNS in AD. During AD, it has been reported that the CSF endothelin-1 levels are decreased in patients and its expression has been found to be elevated in frontal and occipital cortex [64, 65]. In contrast to our results, endothelin-3 levels are unchanged in AD.
Calsynthenin-1 and calsynthenin-3, transmembrane proteins that belong to the cadherin superfamily were increased in SHR-72 Tg animals. There are three members of calsynthenins: calsynthenin-1, 2, and 3, which regulate inhibitory synapse formation and function and neuron development [66]. In Parkinson’s disease, the CSF calsynthenin-1 isoform 2 levels were increased [67]. Consistent with our data, the accumulation of calsyntenin-3 fragments has been described in dystrophic neurites surrounding amyloid-β plaques in Tg2576 mice and AD brain [68]. However, patients with familiar mutations in presenilin-1 and amyloid precursor protein genes have decreased calsynthenin-3 in the CSF [69].
We also observed increased augurin protein in the CSF of SHR-72 Tg animals. This is in correlation with previous report showing increased expression of Esophageal Cancer Related Gene-4, the gene that encodes augurin, in tau Tg mice [70]. Augurin is a hormone-like secreted peptide that is present in large concentrations in the choroid plexus epitelia. Following CNS injury, augurin acts as an acute phase response protein and is released into theCSF [71].
In conclusion, using peptidomic analysis of CSF from transgenic rat model of tauopathy we identified deregulation of several proteins that are associated with tau induced neurodegeneration. Few of these proteins have been previously associated with neurodegeneration in humans, including AD. A detailed investigation of molecular pathways behind alterations in levels of these proteins could in the future serve as an important tool for validation of potential biomarkers for human neurodegenerativediseases.