
Abstract
In the last decade, the association between diet and cognitive function or dementia has been largely investigated. In the present article, we systematically reviewed observational studies published in the last three years (2014–2016) on the relationship among dietary factors and late-life cognitive disorders at different levels of investigation (i.e., dietary patterns, foods and food-groups, and dietary micro- and macronutrients), and possible underlying mechanisms of the proposed associations. From the reviewed evidence, the National Institute on Aging–Alzheimer’s Association guidelines for Alzheimer’s disease (AD) and cognitive decline due to AD pathology introduced some evidence suggesting a direct relation between diet and changes in the brain structure and activity. There was also accumulating evidence that combinations of foods and nutrients into certain patterns may act synergistically to provide stronger health effects than those conferred by their individual dietary components. In particular, higher adherence to a Mediterranean-type diet was associated with decreased cognitive decline. Moreover, also other emerging healthy dietary patterns such as the Dietary Approach to Stop Hypertension (DASH) and the Mediterranean-DASH diet Intervention for Neurodegenerative Delay (MIND) diets were associated with slower rates of cognitive decline and significant reduction of AD rate. Furthermore, some foods or food groups traditionally considered harmful such as eggs and red meat have been partially rehabilitated, while there is still a negative correlation of cognitive functions with saturated fatty acids and a protective effect against cognitive decline of elevated fish consumption, high intake of monounsaturated fatty acids and polyunsaturated fatty acids (PUFA), particularly n-3 PUFA.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is the most common form of dementia. Over 5.4 million people are estimated to be affected in the Unites States [1]. In the last 20 years, most of the therapeutic approaches were directed against the production and accumulation of amyloid-β (Aβ), one of the neuropathological hallmarks of AD, and, up to now, have failed to show clinical efficacy [2].
Currently, available drugs for the treatment of AD have only symptomatic effects [2], and there is an unmet need of preventing AD onset and delaying or slowing disease progression from mild cognitive impairment (MCI) in the absence of disease-modifying therapies. Therefore, at present, the management of potential risk factors is believed to be the most effective way of preventing dementia, AD, and MCI. In the last decade, one of the most intriguing and appealing links hypothesized was the association between lifestyle factors such as diet and dietary habits and the occurrence of AD and dementia [3–7]. Dietary factors may affect the risk of cardiovascular disease (CVD), also influencing the risk of AD and dementia [8–10]. A growing body of evidence suggests that certain diets are associated with a lower incidence of AD, so maintaining a healthy diet may have an impact on many of these possible risk factors for cognitive decline, and the model to follow seems to be the Mediterranean diet (MeDi) [4, 9]. Therefore, elevated dietary monounsaturated fatty acids (MUFA) and n-3 polyunsaturated fatty acids (n-3 PUFA) and high fish consumption [10, 11], alongside high levels of antioxidants from fruit and vegetables [12, 13], and moderate alcohol and coffee consumption [14–16] may have a beneficial effect on the risk of dementia. Nonetheless, diet should be considered as a “whole”, consisting of a complex of nutritional principles, foods, micronutrients and macronutrients interacting with each other. In fact, the combinations of foods and nutrients into certain patterns may act synergistically to provide stronger health effects than those conferred by their individual dietary components. This hypothesis has addressed the interest in the role of dietary patterns rather than of the individual components of the diet against CVD, neurodegenerative diseases, and all-cause mortality [17, 18]. Some recent systematic reviews and meta-analyses of pooled studies on this issue found that a higher adherence to the MeDi was associated with a reduced risk of cognitive impairment, MCI and AD, as well as the transition from MCI to AD [19–22]. However, in the last few years, some changes have emerged in approaching the relationship between diet and cognitive impairment. In fact, the National Institute on Aging–Alzheimer’s Association (NIA-AA) guidelines for AD and cognitive decline due to AD pathology [23] introduced some evidence suggesting a direct relation between diet and changes in the brain structure and activity, opening the era of brain imaging biomarkers in nutritional epidemiology. The aim of the present systematic review article was to shed light on the relationship among dietary patterns, foods, food groups, micro-, and macronutrients and late-life cognitive disorders considering the results of observational studies published in the last three years (2014–2016), as well as possible underlying mechanisms of the proposed associations.
METHODS
In the present systematic review article, we followed the Preferred Reporting Items for Systematic reviews and Meta-Analyses (PRISMA) guidelines, adhering to the PRISMA 27-item checklist [24]. This systematic review was based upon searches of US National Library of Medicine (PubMed), Ovid MEDLINE, EMBASE, Google Scholar, Web of Science, and Scopus databases, picking the following terms to identify the risk exposure (dietary patterns OR foods OR food groups OR micronutrients OR macronutrients) combined with terms to determine the outcomes of interest [cognitive AND (impairment OR decline OR disorders) OR cognition OR Alzheimer’s disease OR dementia OR mild cognitive impairment OR vascular dementia]. There were no language restrictions on the search. To be included in this systematic review, studies were limited to observational studies (including both cross-sectional and longitudinal population-based studies) published between January 1, 2014 and December 31, 2016. We choose these time limits on the basis of a pre-search without time limits that included a very high number of identified articles to review (36104 articles). Studies were further required to meet the following inclusion criteria: (1) studies conducted in humans aged 45 years or older; (2) studies that provided a description of the tools used for collecting the adherence to the different dietary patterns and the intake of foods, food groups, micro- and macronutrients (e.g., validated semi-quantitative food frequency questionnaires, 7-day dietary records, or 24-h dietary recall) or that evaluated nutrient consumption from the values of biochemical markers (e.g., serum concentration or red blood cells levels); (3) studies assessing cognitive functions according to the diagnostic criteria for the diagnoses of MCI [Petersen criteria and their revision/modifications, International Working Group on Mild Cognitive Impairment criteria, European Alzheimer’s Disease Consortium (EADC) criteria, NIA-AA criteria for MCI due to AD, and DSM-5 criteria for Mild Neurocognitive Disorder), AD [National Institute of Neurological and Communicative Disorders and Stroke and the Alzheimer’s Disease and Related Disorders Association (NINCDS-ADRDA) criteria, NIA-AA criteria for dementia due to AD, International Working Group (IWG)-1 criteria for AD, and IWG-2 criteria for AD], vascular dementia (VaD) (NINCDS-AIREN), unspecified dementia [Diagnostic and Statistical Manual for Mental Disorders (DSM)-III-R criteria, DSM-IV criteria, DSM-IV-TR criteria, DSM-5 criteria for Major Neurocognitive Disorder, International Classification of Diseases (ICD), 9th Revision, Clinical Modification (CM), and ICD-10-CM] or that provided the neuropsychological tools used for defining late-life cognitive impairment/decline also in non-demented older subjects. The studies included had to present original data.
Figure 1 shows the stages in obtaining studies for inclusion in the present report (PRISMA Four-phase Flow Diagram). From 9,074 articles identified with multiple electronic searches, we screened titles and abstracts of the citations downloaded from the searches and identified 1,877 potential relevant articles chosen for a closer review. We excluded 1,798 articles not meeting inclusion criteria and obtained full copies of the 79 potentially suitable reports for further assessment. After inclusion of 4 articles of interest from the reference lists of the selected articles and exclusion of another 36 articles, 47 studies met study eligibility criteria and were finally included in the overall systematic review (Tables 1–3) [25–71]. We did not use formal methods of assessment of the quality of the studies included in the present systematic review, given the lack of randomized clinical trials, but we described in depth study design and sample together with dietary and cognitive assessment and principal results of the included studies in Tables 1–3 [25–71]. Finally, we used a narrative synthesis to summarize the findings of the included studies, subdividing the articles for the four principal diet-based approaches (dietary patterns, foods and food groups, micronutrients, and macronutrients), specifying sample size and study design and the cognitive outcomes of the included studies (late-life cognitive impairment/decline or dementia, AD, and non-AD dementias), when possible.
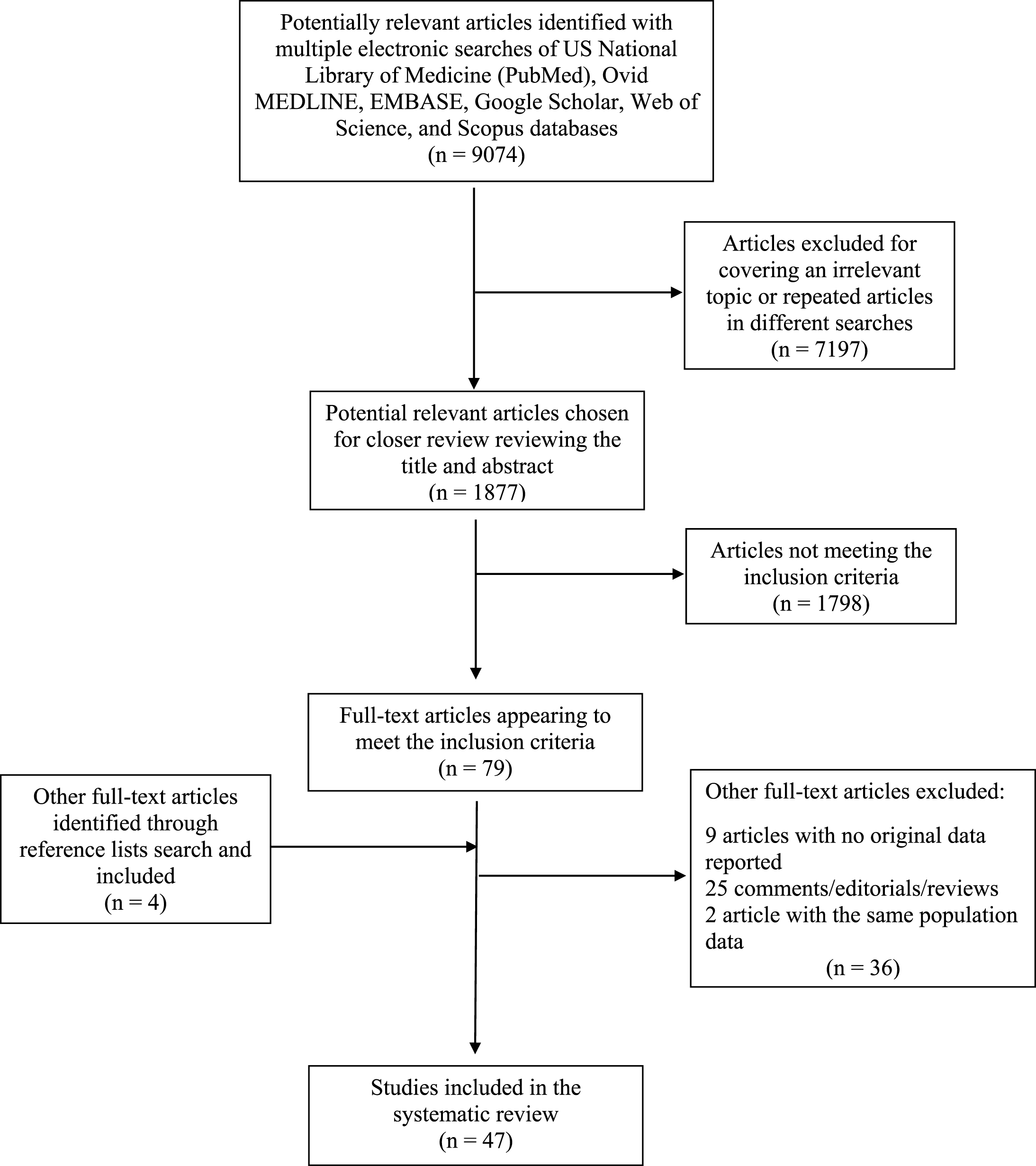
PRISMA Four-phase Flow Diagram of retrieved and selected articles showing associations among dietary patterns, foods, food groups, micro- and macronutrients, and late-life cognitive disorders considering the results of observational studies published in the last three years (2014–2016).
Observational studies on the relationship among dietary patterns and late-life cognitive disorders (2014–2016)
NHANES, National Health and Nutrition Survey; MABAT ZAHAV, Israeli National Health and Nutrition Survey; MeDi, Mediterranean diet; MDS, Mediterranean diet score; OR, odds ratio; CI, confidence interval; FFQ, Food Frequency Questionnaire; TICS-m, Telephone Interview of Cognitive Status-modified; MMSE, Mini-Mental State Examination; SU, standard units; AusMeDi, Australian-style Mediterranean diet; DASH, Dietary Approach to Stop Hypertension; MAP, Memory and Aging Project; AD, Alzheimer’s disease; MIND, Mediterranean-DASH diet Intervention for Neurodegenerative Delay; CDR, Clinical Dementia Rating scale; HR, hazard ratio; MRI, magnetic resonance imaging; CT, cortical thickness; ROI, region of interest; PET, positron emission tomography; PiB, 11C-Pittsburgh compound-B; FDG, 18F-fluorodeoxyglucose; METglc, glucose metabolism; TGMV, total gray matter volume; MUFA, monounsaturated fatty acids; PUFA, polyunsaturated fatty acids; SFA, saturated fatty acids; TBV, total brain volume; TWMV, total white matter volume; mCT, mean cortical thickness; SD, standard deviation; SE, standard error; GM, gray matter; WM, white matter; mAHEI, modified Alternative Healthy Eating Index; IL-6, interleukin-6; IDP, inflammatory dietary pattern.
Observational studies on the relationship among foods, food-groups, and late-life cognitive disorders (2014–2016)
TICS-m, Telephone Interview of Cognitive Status modified; CI, confidence interval; FFQ, Food Frequency Questionnaire; AD, Alzheimer’s disease; PUFA, polyunsaturated fatty acids; LDQ, Lifetime Diet Questionnaire; MCI, mild cognitive impairment; MoCA, Montreal Cognitive Assessment; CSI-D, Community Screening Instrument for Dementia; OR, odds ratio; HVLT IR, Hopkins Verbal Learning Test immediate recall; SU, standard units; HR, hazard ratio; CERAD, Consortium to Establish A Registry for Alzheimer’s Disease; AMR, arithmetic mean ratio; MMSE, Mini-Mental State Examination; BVRT, Benton Visual Retention Test; CVLT, California Verbal Learning Test; VFT-L, Verbal Fluency Test - Letter; VFT-C, Verbal Fluency Test - Categorical; TMT A, Trail Making Test Part A; TMT B, Trail Making Test Part B; DS-F, Digit Span Forward test; DS-B, Digit Span Backward test; NAS, Nutrient adequacy score; CSF, cerebrospinal fluid; Aβ, amyloid-β; EADC, European Alzheimer’s Disease Consortium; SU.VI.MAX, Supplémentation en Vitamines et Minéraux Antioxydants; CDP, carotenoid-rich dietary pattern; RRR, reduced rank regression.
Observational studies on the association of micro- and macronutrients with late-life cognitive disorders (2014–2016).
MMSE, Mini-Mental State Examination; CLVT-List A, California Verbal Learning Test immediate free recall List A; CVLT-DFR, California Verbal Learning Test immediate Delayed Free Recall; DS-F, Digit Span Forward; DS-B, Digit Span Backward; BVRT, Benton Visual Retention Test; CVLT, California Verbal Learning Test; AF, Animal Fluency test; BTA, Brief Test of Attention; TMT A, Trail Making Test Part A; TMT B, Trail Making Test Part B; CDT, Clock Drawing Test; CR, Card Rotations; IP, Identical Pictures; CES-D, Center for Epidemiologic Studies Depression Scale; SCI, subjective cognitive impairment; MCI, mild cognitive impairment; AD, Alzheimer’s disease; 25(OH)D, 25-hydroxyvitamin D; CSF, cerebrospinal fluid; Aβ, amyloid-β; IWG-MCI, International Working Group on Mild Cognitive Impairment; OR, odds ratio; CI, confidence interval; MRI, magnetic resonance imaging; PET, positron emission tomography; PiB, (11)C-Pittsburgh Compound-B; FDG, (18)F-fluorodeoxyglucose; FFQ, Food Frequency Questionnaire; PUFA, polyunsaturated fatty acids; 3MS, modified MMSE; DSST, Digit Symbol Substitution Test; HR, hazard ratio; SE, standard error; CDR, Cognitive Drug Research; RI-48, rappel indicé-48 items; DST, Digit Symbol Test; KOLT, Kendrick Object Learning Test; COWAT, Controlled Oral Word Association Test; MMSE-KC, Korean version of MMSE in the Korean version of the Consortium to Establish a Registry for Alzheimer’s disease Assessment Packet (CERAD-K); BN, Boston Naming; WF, Word Fluency; WLM, Word List Memory; CR, Constructional Recall; WLR, Word List Recognition; CPT, Constructional Praxis Tests; TUP, total urinary polyphenols; TDP, total dietary polyphenols; IST, Isaac’s set test; CERAD, Consortium to Establish a Registry for Alzheimer’s disease; hs-CRP, high-sensitivity C reactive protein; DHA, docosahexaenoic acid; RBC, red blood cell; SD, standard deviation.
RESULTS
Dietary patterns and late-life cognition
Table 1 shows selected observational studies published in the last three years that evaluate the association of dietary patterns with late-life cognitive disorders [25–41]. Among these diets, MeDi is a typical dietary pattern of Mediterranean countries, characterized by high consumption of fruits, vegetables, legumes and cereals, olive oil as the main added lipid, moderate consumption of alcohol (mainly wine and during meals) and low consumption of red meat and dairy products. It is doubtless the most analyzed dietary pattern, and accumulating evidence support a potential protective role against cognitive decline and dementia, although there are still inconsistencies in the reported data. In particular, the findings from prospective studies and very recent systematic reviews and meta-analyses suggested that adherence to the MeDi fulfilling the whole-diet approach may affect not only the risk of AD, but also of predementia syndromes and their progression to overt dementia [19]. A cross-sectional analysis from the US National Health and Nutrition Survey (NHANES) 1999–2002 and from the Israeli National Health and Nutrition Survey (MABAT ZAHAV) 2005–2006 suggested that cognitive and physical functions were significantly better among older subjects with the higher adherence to the MeDi [25] (Table 1). In the Spanish prospective cohort of the SUN project, in an 8-year follow-up, a higher cognitive decline was observed among participants with low or moderate baseline adherence to the MeDi than among those with better adherence [26] (Table 1). In the European Prospective Investigation into Cancer and Nutrition (EPIC) study, in a cohort of Greek elderly population that still adhered to the traditional MeDi, it was demonstrated that closer adherence to MeDi was associated with less decline in Mini-Mental State Examination (MMSE) performance over a period of about 7 years, especially in individuals aged 75 years or older [27] (Table 1). The association between dietary patterns and cognitive decline was also recently investigated in non-Western populations [28, 29] (Table 1). In fact, in a prospective Chinese cohort study on 1,650 Chinese adults≥55 years of age, the subjects in the highest tertile of a modified MeDi score or a wheat-based diverse diet with similar features of the adapted MeDi had a slower rate of cognitive decline [28]. Moreover, in the population-based Australian Imaging, Biomarkers and Lifestyle Study of Ageing, higher adherence to the Australian-style MeDi was associated with better performance in the executive function, while higher Western diet adherence was associated with greater decline in the visuospatial cognitive domain [29].
Other emerging dietary patterns are the Dietary Approach to Stop Hypertension (DASH) and the Mediterranean-DASH diet Intervention for Neurodegenerative Delay (MIND) diets. The DASH diet is characterized by low consumption of saturated fat and commercial pastries and sweets, and higher intake of dairy than in the MeDi. In the last three years, in the Memory and Aging Project (MAP) study, a prospective study on older adults with 4 years of follow-up, the DASH pattern was associated with slower rates of cognitive decline. In particular, a 1-unit-higher DASH score, was equivalent of being at least 4.4 years younger [30] (Table 1). These results were in line with those of Morris and colleagues, in the same MAP study, in which higher adherence to DASH diet was related with greater reduction of incident AD rather than higher adherence to MeDi (54% and 39% reduction, respectively) [31] (Table 1).
The MIND diet was based on the dietary components of the MeDi and DASH diet with modifications that highlight the foods and nutrients shown to be associated with dementia prevention. Among the MIND diet components, there are 10 brain healthy food groups (green leafy vegetables, other vegetables, nuts, berries, beans, whole grains, seafood, poultry, olive oil, and wine) and five unhealthy food groups (red meats, butter and stick margarine, cheese, pastries and sweets, and fried/fast food). Hence, MIND diet uniquely specifies consumption of berries and green leafy vegetables and does not specify high fruit consumption (both DASH and MeDi), high dairy (DASH), high potato consumption, or >1 fish meal per week (MeDi). Other recent findings from the MAP study suggested that higher MIND diet score was associated with slower decline in cognitive abilities [32] (Table 1). The rate reduction for persons in the highest tertile of diet scores compared with the lowest tertile was the equivalent of being 7.5 years younger. MIND diet score was also more predictive of cognitive decline than either of the other diet scores (DASH and MeDi) [32]. Furthermore, in a follow-up of 4.5 years of the MAP study, participants with higher and moderate adherence to MIND diet had statistically significant reduction in AD rate compared with those with lower adherence (53% and 35% respectively) [31] (Table 1). Instead, only the highest tertiles of the DASH and MeDi scores were significantly associated with incident AD reduction [31].
Despite the promising results of these two diets, to date, we have brain imaging data only on the correlation with the MeDi (Table 1). The few cross-sectional studies carried out on cognitively normal people showed that higher adherence to MeDi was related to greater magnetic resonance imaging (MRI)-based cortical thickness in AD-vulnerable regions and larger brain volumes. MeDi effects on MRI biomarkers were significant in the left, but not in the right hemisphere, and were most pronounced in entorhinal cortex, orbito-frontal cortex and posterior cingulate cortex [33] (Table 1). In the same sample of cognitively normal people, 5 different nutrients patterns (NPs) were identified and associated with major brain AD biomarkers. NP4 scores (vitamins B12, D, and zinc) were positively associated with glucose metabolism investigated with 18F-fluorodeoxyglucose positron emission tomography (PET) scans and MRI-based total gray matter volume, and negatively associated with PET- 11CPittsburgh compound-B (PiB) retention in AD-vulnerable regions. Glucose metabolism and total gray matter volume were also positively associated with NP2 scores (vitamin E, MUFA, and PUFA), and negatively associated with NP5 scores [saturated fatty acids (SFA), trans unsaturated fatty acids, cholesterol and sodium], and glucose metabolism was positively associated with higher NP3 scores (vitamin A, vitamin C, carotenoids, and dietary fibers) [34] (Table 1). Higher adherence to a Mediterranean dietary pattern was associated with larger MRI measures of cortical thickness and with several individual region of interests (ROIs) that undergo age-related or AD-related neurodegeneration, was marginally associated with temporal and AD signature cortical thickness, and was not associated with hippocampal volume [35] (Table 1). This finding may be explained with the observation from the Alzheimer’s Disease Neuroimaging Initiative in which presymptomatic individuals had significantly reduced cortical thickness in AD-vulnerable regions compared to controls but did not differ in regard to hippocampal volume [72]. In the Washington Heights–Inwood Community Aging Project (WHICAP), higher MeDi adherence was associated with less brain atrophy (larger total brain volume, total gray matter volume, total white matter volume), with an effect similar to 5 years of aging [36] (Table 1).
In the last three years, only one prospective imaging-diet study on older adults was conducted [37] (Table 1), confirming other results coming from cross-sectional studies. In fact, Jacka and colleagues, in the Personality and Total Health Through Life Study, found that a healthy “prudent” dietary pattern characterized by intake of fresh vegetables, salad, fruit and grilled fish was associated with a larger left hippocampal volume on MRI over 4 years of follow-up [37] (Table 1). In particular, every one standard deviation (SD) increase in healthy “prudent” dietary pattern was associated with a 45.7 mm3 larger left hippocampal volume [37]. While higher consumption of an unhealthy Western dietary pattern characterized by intake roast meat, sausages, hamburgers, steak, chips, crisps, and soft drinks was independently associated with a 52.6 mm3 smaller left hippocampal volume [37]. The difference in hippocampal volume between those classified with a healthy and or unhealthy diet was 203 mm3, a difference which corresponds to 62% of the average decline in left hippocampal volume observed over the 4-year period. It was found no interaction between right hippocampus volumes and the two dietary factor scores [37]. Other studies suggested a strong impact of healthy diets on structural connectivity in older subjects, rather than gray and white matter volumes. In fact, through diffusion tensor imaging (DTI) at MRI examination was seen that higher adherence to the MeDi was associated with preserved white matter microstructure in multiple brain areas and appeared to delay cognitive aging by up to 10 years [38] (Table 1). None of the individual components was strongly associated with DTI parameters, supporting the hypothesis that overall diet quality may be more important to preserve brain structure than any single food. These results suggested the involvement of vascular pathways rather than neurodegenerative mechanisms in the link between the MeDi and lower risks of cognitive decline and related diseases [38].
The importance of components of prudent dietary pattern (vegetables, fruit, cooking/dressing oil, cereals and legumes, whole grains, rice/pasta, fish, low-fat dairy, poultry and water) was confirmed by the observation that the MMSE decline associated with Western diet may be attenuated by high adherence to prudent pattern [39] (Table 1). In fact, the decline became less pronounced (53.5%) and non-significant among people who had a high adherence to both the prudent and Western patterns. Furthermore, Western dietary pattern score was significantly associated with all-cause mortality in the older age cohorts [39]. Instead, people followed healthiest diet was slightly older, more active, less likely to smoke, had a lower body mass index, normal serum creatinine, and had higher MMSE score [40] (Table 1). The healthiest diet was associated with a reduction of about 24% in risk of cognitive decline and in particular was shown a significant association between higher diet quality and reduced risk of decline in 4 components of the MMSE including copying, attention and calculation, registration, and writing [40]. The brain damage related to an unhealthy diet may be based on a pro-inflammatory mechanism. Ozawa and colleagues detected an inflammatory dietary pattern characterized by higher intake of red meat, processed meat, peas and legumes, and fried food, and lower intake of whole grains which correlated with elevated interleukin (IL)-6 [41] (Table 1). It was related with greater decline in reasoning and in global cognition and, in a cross-sectional analysis at baseline, a two times greater risk of having a decline of 3 points or more in MMSE [41].
Foods, food groups, and late-life cognitive disorders
Fish and seafood
Table 2 shows selected observational studies published in the last three years and evaluating the relationship among foods, food-groups, and late-life cognitive disorders [42–54]. In particular, the emerging data from the last studies on the correlation between fish and seafood consumption and cognitive decline are conflicting. Significant correlations were found in some particular population subgroups [≥65 years and apolipoprotein E (APOE) ɛ4 carriers]. Age significantly modified the association between fish consumption and cognitive change [42] (Table 2). In fact, no association was observed among adults aged 55–64 years. Conversely, adults aged≥65 years, that consuming≥1 servings/week fish (i.e., 100 g) had a reduction of cognitive decline rate [42]. Compared with individuals who consumed <1 serving/week fish, the mean annual rate of global cognitive decline was reduced by 0.35-point equivalent to the disparity associated with 1.6 years of age. Removing shellfish and/or preserved fish from the total fish did not appreciably alter the results [42].
Interestingly, Morris and colleagues showed that, in APOE ɛ4 carriers, seafood consumption≥β1 meals/week was correlated with lesser burden of brain AD neuropathology, including lower density of neuritic plaques, less severe and widespread neurofibrillary tangles, and lower neuropathologically defined AD [43] (Table 2). Furthermore, some studies demonstrated an association between fish consumption and MRI biomarkers [34, 35] (Table 1). In the Mayo Clinic Study of Aging, higher fish intake was associated with larger cortical thickness summary measures for parietal and average lobar cortical thickness and marginally associated with AD signature cortical thickness, temporal and frontal cortical thickness, and also associated with several individual cortical thickness measures: precuneus, superior parietal, posterior cingulate, supramarginal, middle temporal, and inferior parietal, and marginally associated with fusiform CT [34]. Higher fish consumption was also related with larger total gray matter volume [35]. Fish consumption was also associated with a slower decline in composite and verbal memory scores [42] (Table 2). Of note, Danthiir and colleagues demonstrated that more frequent consumption of total fish (oily and white) was associated with slower cognitive speed for the constructs of inhibition, simple/choice reaction time, reasoning speed, and memory scanning [44] (Table 2). More frequent consumption of oily fish was significantly associated with worse inhibitory processes; similarly, consumption of white fish significantly and negatively predicted simple/choice reaction time [44]. Danthiir and colleagues hypothesized that the negative trends observed between cognitive performance and fish consumption were due to neurotoxic contaminants in fish, such as methylmercury [44]. However, as seen above, Morris and colleagues found that higher brain levels of mercury were not correlated with brain neuropathology [43] (Table 2). Other studies did not suggest evidence that higher fish intake may impact positively cognitive performance in older adults with cognitive impairment [45, 46] or in those cognitively normal [46, 47] (Table 2). However, Dong and colleagues found that cognitively normal Chinese older subjects consumed more fish than MCI subjects [45], and Zhao and colleagues suggested that higher consumption of marine product was associated with a significantly decreased odd of suffering from MCI [47], although in these two Chinese studies, MCI was diagnosed only on the basis of a screening cognitive test (the Montreal Cognitive Assessment - MoCA) and not with clinical criteria (Table 2).
Fruits and vegetables
In Greece, in the EPIC study, among the components of MeDi, only vegetable consumption exhibited a significant inverse association with cognitive decline [27] (Table 1). The diet low in fruit and vegetable might increase the risk of cognitive function decline in older adults [45] (Table 2). In fact, adherence to WHO recommendations for daily intakes of fruit and vegetable, that are eating 5 or more portions of fruit and/or vegetables a day (≥400 g/day), were significantly associated with a 47% decreased prevalence of cognitive impairment [45]. In contrast to these findings, Xu and colleagues found that among older adults (≥68 y) being vegetarian (not eating meat), the risk for cognitive impairment increased almost 4-fold [48] (Table 2). Imaging data in older cohort showed that higher intake of total vegetables was associated with larger dorsolateral prefrontal and superior parietal cortical thickness, while vegetables without legumes were associated with larger middle temporal, superior parietal, and dorsolateral prefrontal cortical thickness [35] (Table 1). In contrast, fruit consumption was negatively associated with inferior parietal, supramarginal, superior parietal, parietal, and precuneus cortical thickness [35]. These findings are in keeping with result of another study in which higher fruit intake was associated with lower temporal and hippocampal volumes [36] (Table 1). This is probably due to high content of simple sugars and a high glycemic index of several fruits and so the effects of carbohydrate component on increased risk of MCI [73]. In older adults, fruit intake would benefit name ability and attention level, while vegetables intake would benefit orientation ability [45] (Table 2). Finally, consumption of green vegetables was independently associated with better memory function and among older elderly (≥68 y) it reduced the risk for cognitive impairment by almost 20% [48] (Table 2).
Nuts
Nuts are rich in PUFA (n-3 and n-6) and MUFA, and also contain a significant amount of minerals, such as phosphorus, potassium, magnesium, calcium, iron, and sulfur, and vitamins, such as B1, B2, B6, and E. It was found the nut intake of MCI patients was less than that of cognitively normal subjects [45] (Table 2). In fact, a study performed on older women found that higher total nut intake (i.e.,≥5/week) over the long term was associated with modestly better cognitive performance [49] (Table 2). Increasingly higher total nut intake was related to increasingly better overall cognition at older age. Considering that one year of age was associated with a mean decline of 0.04 standard units on both the global and verbal composite scores, the mean differences comparing the highest to lowest categories of nut intake were equivalent to approximately two years of cognitive aging [49]. In the same study, it was found a suggestion that those who consumed walnuts 1 to 3 times per month had better cognition than those who consumed walnuts less than once per month, but there was no overall trend of increasingly better cognitive performance with increasing walnut intake [49]. Dong and colleagues also showed that nut intake benefited delayed memory [45] (Table 2).
Coffee and caffeine intake
A recent systematic review suggested that several cross-sectional and longitudinal population-based studies suggested a protective effect of coffee, tea, and caffeine use against late-life cognitive impairment/decline, although the association was not found in all cognitive domains investigated and there was a lack of a distinct dose-response association, with a stronger effect among women than men [74]. The findings on the association of coffee, tea, and caffeine consumption or plasma caffeine levels with incident MCI and its progression to dementia were too limited to draw any conclusion [74]. Furthermore, for dementia and AD prevention, some studies with baseline examination in midlife pointed to a lack of association, although other case-control and longitudinal population-based studies with briefer follow-up periods supported favorable effects of coffee, tea, and caffeine consumption against AD [74]. Recent findings from the Italian Longitudinal Study on Aging (ILSA) suggested that cognitively normal older individuals who increased their coffee consumption had a higher rate of developing MCI, while a constant in time moderate coffee consumption was associated to a reduced rate of the incidence of MCI [50] (Table 2). Among older adults in Brazil, coffee consumption was associated with better cognitive performance on memory and efficiency of searching in long-term memory (drinking 2–3 cups of coffee per day was associated with about a 3% increase in the mean number of words remembered on the learning, recall, and word recognition tests) [51] (Table 2). Also, drinking≥3 cups/day of coffee was associated with an increase of about 1.23 words in the mean number of words pronounced in the semantic verbal fluency test [51]. However, in this Brazilian study, Araujio and colleagues did not find indication of a dose response relationship in these associations [51]. In the Chinese study on cognitively normal and MCI adults of Dong and colleagues, no significant association was detected between drinking of coffee and cognitive function [45] (Table 2). Another aspect of coffee assumption is the role of its component such as the caffeine. Coffee is a rich source of caffeine, which acts as a psychoactive stimulant. In a cross sectional analysis, Beydoun and colleagues found that caffeine intake was associated with better global cognitive function (MMSE) at baseline for patients≥70 years [52] (Table 2). However, in a study that evaluated the association of caffeine consumption with the cerebrospinal fluid (CSF) biomarkers, particularly Aβ, in AD and MCI patients, no significant difference was found in daily consumption of caffeine between MCI and AD patients, with no correlation between caffeine consumption and Aβ1-42 in the CSF [53] (Table 2). In the same study, theobromine, xanthine formed upon caffeine metabolism and also directly ingested from chocolate products, was associated with a favorable Aβ profile in the CSF [53]. Interestingly, theobromine in the CSF did not correlate with caffeine consumption, theobromine consumption, or the levels of caffeine and other xanthines in the plasma, but instead it correlated with levels of caffeine, theophylline, and paraxanthine in the CSF, suggesting that it may be formed by central metabolic pathways [53].
Eggs
Eggs have a high content of proteins and lipids, in particular cholesterol. For this reason, they are traditionally considered an unhealthy food. However, eggs have also a significant amount of vitamins A, B6, B12, riboflavin, folic acid, choline, iron, calcium, phosphorus and potassium. In a recent study, higher daily intake of eggs reduced of about 3% the odds of suffering from MCI [47] (Table 2). Instead, in the Chinese study of Dong and colleagues, no significant association was detected between intake of eggs with cognitive function in normal and MCI adults [45] (Table 2).
Tofu
Tofu is a common food in most of the Far East. It is obtained from curdling of the juice extracted from soybeans. It has a high protein and PUFA content. Higher weekly intake of tofu was associated with worse memory performance, furthermore among older elderly (≥68 y), high tofu intake increases the risk (of almost 30%) of cognitive impairment indicative of dementia [48] (Table 2).
Meat
Red meat is a classical element of Western diet that, as mentioned previously, was associated with worse cognitive performance in several imaging studies [37, 41] (Table 1). Consistent with these findings, a negative association of red meat with inferior and superior parietal cortical thickness was found [35] (Table 1). However, this concept should be partially reviewed. In fact, in the last years, eating meat (not being vegetarian) was independently associated with better memory function and in older age (≥68 y) with a four-fold decrease in risk of possible dementia [48] (Table 2). Furthermore, Staubo and colleagues also observed that higher red meat intake was associated with larger entorhinal cortical thickness [35] (Table 1). This effect could also be related to some beneficial components of lean red meat (iron, protein, MUFA, PUFA, cobalamine) and beneficial effects in increasing satiety and reducing weight gain. In the Chinese study of Dong and colleagues, no significant association was detected between intake of light or red meat with cognitive function in normal and MCI adults [45] (Table 2).
Oil
Dong and colleagues, in their Chinese cohort, found that oil intake of MCI patients was less than the normal subjects (29.76 versus 35.20 mL cooking oil per day), and, in particular, would have a positive impact on visual-spatial ability [45] (Table 2). Vegetable oils are rich in carotenoids, and in the Supplémentation en Vitamines et Minéraux Antioxydants (SU.VI.MAX) study carotenoids were associated with higher cognitive performance [54] (Table 2). Extra-virgin olive oil (EVOO) is one of the main elements of MeDi, and clinical trials and population studies indicated that this dietary pattern and its main lipid component EVOO could have a protective role against AD [75].
Legumes
Dong and colleagues, in their Chinese cohort, showed that normal subjects consumed more legumes and legume products than MCI subjects, demonstrating that intake of legumes and legume products would benefit overall cognition level [45] (Table 2). These data were confirmed by imaging biomarkers; in fact, Staubo and colleagues also found that higher intake of legumes was associated with larger parietal and occipital cortical thickness, and with larger thickness in ROIs for superior parietal, inferior parietal, precuneus, and lingual cortical thickness [35] (Table 1).
In their imaging biomarker study, Staubo and colleagues also showed that intake of whole grains or cereals was associated with larger temporal pole and superior temporal cortical thickness [35] (Table 1). Conversely, lower intake of whole grains was associated with higher inflammatory markers (IL-6) and accelerated cognitive decline at older age in the Whitehall II prospective cohort study [41] (Table 1). However, in the Chinese cohort of Dong and colleagues, no significant association was detected between intake of whole grains and cognitive function in normal and MCI adults [45] (Table 2).
Alcohol
Recent findings from the Baltimore Longitudinal Study of Aging suggested that alcohol intake was associated with slower improvement on letter fluency and global cognition among those aged <70 years at baseline [52] (Table 2). Conversely, alcohol intake was associated with better attention and working memory performance, particularly among men and individuals≥70 years at baseline [52]. Compared with moderate consumption (14 to 28 g/d), individuals with higher alcohol intake (>28 g/d) had faster decline or slower improvement on the MMSE, particularly among women and in the older group. Overall, among men, and for those aged≥70 years, lower alcohol intake (<14 g/d) compared with moderate consumption (14 to 28 g/d) was associated with poorer performance in working memory [52]. In the younger group, consuming <14 g/day was associated with slower decline or faster improvement in the letter fluency compared with a moderate intake of 14 to 28 g/day. Similar pattern was showed also for attention and executive functioning [52].
Relation of micro- and macronutrients with cognition in older age
Vitamin E
Table 3 shows selected observational studies published in the last three years and evaluating the association of micro- and macronutrients with late-life cognitive disorders [55–71]. Vitamin E is a potent antioxidant and seems to slow the effects of aging. It is found mainly in nuts, seeds, and oils. Vitamin E is composed by four tocopherols (α-, γ, δ, and β-tocopherols) and four corresponding tocotrienols, and it is known that α- and γ-tocopherols found in foods were linked to slower rate of cognitive decline over a 6-year period [76]. More recently, in the HANDLS study, Beydoun and colleagues investigated the effects on cognitive domains of only one of its isoform, α-tocopherol, showing that vitamin E was positively associated with performance in verbal memory and fluency in the total population, particularly among the younger age group (<48 y) and psychomotor speed among women [55] (Table 3).
Vitamin D
Vitamin D deficiency is a common condition in older adults. Furthermore, the detection of hydroxylases for vitamin D activation and vitamin D receptors in neurons and glia suggested that vitamin D might have a role in cognition. In fact, serum 25-hydroxyvitamin D [25(OH)D] was cross-sectionally associated with reduced Aβ1-42 concentrations in CSF [56] (Table 3). These findings were on the same line of Mosconi and colleagues showing an association of higher intake of vitamin D with lower Aβ load in AD regions on PET with 11C-Pittsburgh Compound-B (PiB) [57] (Table 3). Vitamin D deficiency was also associated with greater decline in global cognitive function over 4 years of follow-up [58] (Table 3) and with a substantially increased risk of all-cause dementia and AD over a mean follow-up of 5.6 years [59] (Table 3). In particular, vitamin D deficiency was related with significantly faster declines in both episodic memory and executive function performance, the two cognitive domains strongly associated with AD dementia [60] (Table 3). However, Granic and colleagues found a U-shaped relationship between 25(OH)D and global cognitive function and attention in the very old [61] (Table 3). In fact, both low and high season-specific quartiles of 25(OH)D were associated with higher odds of prevalent cognitive impairment, poorer attention reaction times/processing speed and focused attention/concentration, and greater attention fluctuation [61]. Therefore, it could be hypothesized that the neuroprotective effects of vitamin D may be attained only at moderate but not at low or high 25(OH)D concentrations. On the other hand, very old subjects taking vitamin D supplements could be probably only recently advised by their physician to take vitamin D, perhaps due to an underlying vitamin D deficiency condition [77]. Finally, findings from the SU.VI.MAX study suggested a positive association between higher 25(OH)D concentrations and better short-term and working memory only among individuals with low education [62] (Table 3).
Vitamins B and folate
Vitamin B12 deficiency has been associated with neurological and psychiatric symptoms including cognitive impairment [78]. Additionally, vitamin B12, along with folate and vitamin B6, are key actors in the homocysteine cycle, and the disruption of this cycle can lead to elevated homocysteine, which has been found among individuals with dementia [79] and has been shown to predict dementia prospectively [80]. Despite potential biological mechanisms through which B vitamins could play a role in the onset of dementia, results from observational and intervention studies have been inconsistent. In the last three years, in imaging studies, higher intake of folate was associated with higher glucose metabolism on (18)F-fludeoxyglucose (FDG)-PET [57] (Table 3) and greater superior temporal cortical thickness [35] (Table 1). Positive associations were also found between vitamins B1 and B2 and superior temporal and temporal pole cortical thickness, and vitamin B6 and superior temporal and middle temporal cortical thickness [35]. Higher intake of vitamin B12 was also associated with lower Aβ load in AD regions on PiB-PET. The association of vitamin B12 with PiB retention was independent of gender, APOE, and family history [57].
Data collected on 1,354 subjects from three Australian cohorts suggested that participants with low serum vitamin B12 (<250 pmol/L) and high red cell folate (>1,594 nmol/L) levels were more likely to have impaired cognitive performance when compared to participants with biochemical measurements that were within the normal ranges [63] (Table 3). In the same cohorts, participants with high folate levels, but normal serum vitamin B12, were also more likely to have impaired cognitive performance [63]. Furthermore, in the Hordaland Health Study, plasma vitamin B12 concentrations in the lowest quartile (<274 pmol/l) combined with plasma folate concentrations in the highest quartile (>18·5 nmol/l) were associated with a reduced risk of cognitive impairment compared with plasma concentrations in the middle quartiles of both vitamins [64] (Table 3).
In a cross-sectional study on older Koreans, in the AD group, positive associations were observed between vitamin B2 intake and global cognitive status, language, verbal fluency, and verbal and praxis memory; between vitamin B6 intake and global cognitive status, language, verbal fluency, verbal and praxis memory, and constructional praxis; between vitamin B12 intake and global cognitive status, language, praxis memory, and constructional praxis; and between folate intake and praxis memory [65] (Table 3). In the MCI group, vitamin B2 intake was positively associated with global cognitive status and language, vitamin B6 intake was positively associated with language, and folate intake was positively associated with global cognitive status and verbal memory. No associations were observed in the normal group [65]. Finally, in the Women’s Health Initiative Memory Study, folate intake below the recommended daily allowance at study baseline was associated with increased risk of incident MCI/probable dementia, after controlling for multiple confounders [66] (Table 3). There were no significant associations between vitamins B6 or B12 and MCI/probable dementia, nor any evidence of an interaction between these vitamins and folate intake [66].
Polyphenols
Polyphenols are natural bioactive compounds that have been identified in foods and beverages. They are associated with neuroprotective activity probably due to their antioxidant and anti-inflammatory properties. In the inCHIANTI study, in older adults without dementia, high concentrations of total urinary polyphenols were associated with an approximately 47% lower risk of substantial cognitive decline in global cognitive function and an approximately 48% lower risk of substantial cognitive decline in attention over a 3-year period, but not with executive function. No significant association was found between total dietary polyphenols and any cognitive test [67] (Table 3).
Carotenoids
Carotenoids are ubiquitous colored pigments, found mainly in fruits and vegetables such as kiwi, carrots, and tomatoes or in green leafy vegetables. Among carotenoids, positive association was found between higher intake of β-carotene and dorsolateral prefrontal and temporal pole cortical thickness [35] (Table 1) and with higher glucose metabolism on FDG-PET [57] (Table 3). However, the β-carotene levels were related with reduced glucose metabolism for women, APOE ɛ4-carriers, and participants with positive AD family history [57].
Among carotenoids, findings from the Three-City-Bordeaux cohort suggested that higher lutein concentration, considered as a function of plasma lipids, was consistently significantly associated with a decreased risk of all-cause dementia and AD [68] (Table 3). Moreover, in the SU.VI.MAX study, a carotenoid-rich dietary pattern (CDP) strongly correlated with the plasma status of β-carotene, α-carotene, β-cryptoxanthin, and lutein, was positively correlated with the consumption of green-colored fruits and vegetables, vegetable oils, orange-colored fruits and vegetables and soup, and was negatively correlated with that of beer-cider and wine. CDP score was negatively associated with alcohol and protein intake and positively associated with carbohydrate intake [54] (Table 2). Furthermore, CDP score was positively associated with the composite cognitive performance score assessed 13 years later. More specifically, higher CDP scores were related to better episodic memory, semantic fluency, working memory, and executive functioning [54].
Capsaicin
Capsaicin is a micronutrient acting as an active component of chili peppers. In recent studies, it has been shown to have favorable effects on various diseases including atherosclerosis, CVD, cancer, obesity, gastrointestinal diseases, hypertension, and stroke, and also reducing total and cause-specific mortality [81, 82]. Liu and colleagues, in a cross-sectional study collecting the dietary habits of participants in 12 months, found that capsaicin-rich diet was associated with MMSE and serum Aβ40 levels, but not with serum Aβ42 levels [69] (Table 3). A possible explanation may be that neurons produce more Aβ40 than Aβ42 and serum Aβ42 levels have higher cerebral tropism with increased capacity of aggregation in the brain, so it is less prone to efflux from the brain to blood.
Macronutrients: Fatty acids
Many epidemiological studies have demonstrated that dietary fatty acids may play a key role in several pathological conditions. In particular, an increasing body of epidemiological evidence suggested that elevated SFA could have negative effects on MCI [11], while a clear reduction of risk for cognitive decline has been found in population samples with elevated fish consumption, high intake of MUFA and PUFA, particularly n-3 PUFA [11]. In fact, it is of extreme nutritional importance the intake of essential fatty acids from both the n-3 and n-6 series of PUFA. In the n-3 series, eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) are considered the most important. In particular, DHA might affect the neuronal survival. In agreement with this hypothesis, Baierle and colleagues showed that cognitive function was positively associated with DHA, nervonic acid (MUFA), and total n-3 PUFA, while was inversely related to myristic acid (SFA), palmitic acid (SFA), palmitoleic acid (MUFA), the n-6 PUFA/n-3 PUFA ratio, and homocysteine levels [70] (Table 3). In this study, elevated levels of homocysteine were also inversely associated with total n-3 PUFA which may lead to vascular disease and, thus, may be involved in the risk of cognitive decline and other brain dysfunctions. Therefore, balanced intake of both n-3 and n-6 fatty acids may be essential for health by avoiding a harmful permanent proinflammatory state. These data are supported also by recent imaging studies. In fact, positive associations (higher intake with larger cortical thickness) were observed for n-3 PUFA and linolenic acid (n-3 PUFA) with superior temporal cortical thickness and for linoleic acid (n-6 PUFA) with superior temporal, precuneus, middle temporal, and parietal cortical thickness [35] (Table 1). Furthermore, in postmenopausal women study, n-3 index [red blood cell (RBC) EPA+ DHA levels] was directly associated with the total brain and hippocampal volumes. In particular, a 2 standard deviation (SD) (3.2% absolute) higher n-3 index was associated with a 0.48% (4.2 cm3) larger total brain volume measured 8 years later [71] (Table 3). A marginally significant association between a 1-SD increase in DHA with total and normal brain volume has also been found, instead no association was found for EPA [71]. Furthermore, Pottala and colleagues showed that higher levels of RBC EPA+ DHA were associated with 50 mm3 larger hippocampal volumes [71]. Furthermore, higher intake of n-3 PUFA was associated with lower Aβ load in AD regions on PiB-PET, independently of gender, APOE, and family history [57] (Table 3). On the contrary, in another study, in normal older adults, higher proportions of long-chain n-3 PUFA did not give benefits on cognitive performances [44] (Table 2). Higher concentrations of EPA were associated with poorer perceptual and reasoning speed in females, but not in males, before controlling for current fish intake [44]. Therefore, it is plausible that a physiologically different effect of long-chain n-3 PUFA in relation to gender may lead to differential associations with cognition [44].
Mechanisms underlying the relationship among dietary factors and late-life cognitive disorders
Dietary patterns and late-life cognitive disorders: Possible mechanism models
We need to pursue studies which will improve the knowledge of the biochemical mechanisms underlying the pathophysiological processes and, in a public health perspective, will examine dietary patterns. These mechanisms could be searched at the different levels of investigation of dietary factors linked to late-life cognitive disorders used in the present review article (i.e., dietary patterns, foods and food-groups, and dietary micro- and macronutrients). Consumption of the western diet contributes to the development of chronic conditions, such as atherosclerosis, diabetes, obesity, hypertension, and hyperlipidemia, which in turn results in metabolic, inflammatory, and microvascular changes that induce injury to the white matter of the brain [83, 84]. Subsequently, white matter disease promotes the development and progression of cognitive decline and dementia [85].
There is converging evidence that composite dietary patterns, such as the MeDi, are related to lower risk for CVD, dyslipidemia, hypertension and coronary artery disease, several forms of cancer, cognitive decline, and overall mortality [86, 87]. Several mechanisms have been proposed underlying the suggested protective role of the MeDi against age-related changes in cognitive function, predementia syndromes and dementia: vascular variables and non-vascular biological mechanisms such as metabolic, oxidative, and inflammatory processes [88]. Therefore, MeDi may act on cognition through cerebrovascular mechanisms. Alternatively, AD almost certainly has its pathological origins in midlife, and it is possible that components of the MeDi could affect the metabolism of Aβ or tau protein [89]. Moreover, the model of “gene environment interaction” may be also considered, in light of the recently proposed Latent Early Life Associated Regulation model, in which environmental agents, such as heavy metals and toxins, and dietary factors, could perturb gene regulation in a long-term fashion, beginning at early developmental stages, but these perturbations do not have pathological results until significantly later in life [90].
The DASH is a diet that has been shown by randomized clinical trials in the US to protect against many cardiovascular risk factors of dementia [91]. Some of the protective effects of DASH included lower blood pressure [92] and blood low-density lipoprotein (LDL) cholesterol levels [93], weight reduction [94], reduced oxidative stress and inflammation [95], improved insulin sensitivity and reduced incidence of diabetes mellitus [96]. DASH diet and the MeDi have many similar components, such as emphasis on natural plant-based foods and limited consumption of red meat, but there are differences. The MeDi is a cultural diet that uniquely emphasizes daily intake of olive oil (MUFA), fish, and potatoes, and moderate wine consumption, whereas DASH uniquely emphasizes dairy consumption and restricted intake of sodium, commercial sweets, and saturated fat.
The MIND diet is a refinement of the extensively studied cardiovascular diets, the MeDi and DASH diets, with modifications based on existing studies relevant to nutrition and associated with dementia prevention. MIND diet is not specific to the underlying AD pathology, but perhaps better overall functioning and protection of the brain. A number of prospective cohort studies found that higher consumption of vegetables was associated with slower cognitive decline with the strongest relations observed for green leafy vegetables [12]. Green leafy vegetables of MIND diet are sources of folate, vitamin E, carotenoids, and flavonoids, micronutrients that have been related to lower risk of dementia and cognitive decline [31, 32].
Foods, food groups, and late-life cognitive disorders: Possible mechanism models
Fish consumption has been associated with lower risk of AD because it could have a preventive effect against dementia through its anti-thrombotic and anti-inflammatory properties in addition to its specific effect on neural functions, an effect thought to be mediated through altering gene expression [97]. Furthermore, the favorable effects of fish oil on cerebrovascular function are known, in particular for the impact of DHA on endothelial nitric oxide synthase (eNOS) expression and a lower risk for ischemic stroke [98]. It was hypothesized that some negative epidemiological findings observed for the impact of fish consumption on cognitive performance may be due to neurotoxic contaminants in fish, such as methylmercury [44]. However, as seen in one of the reviewed studies, higher brain levels of mercury were not correlated with brain neuropathology [43]. Diet may affect trace metal concentrations in the brain of subjects with normal cognition [99] or AD [100–102], and transition metals, though crucial to many biochemical neuronal processes, were abnormally aggregated and distributed in AD [103, 104]. Dyshomeostasis in aluminum, silicon, lead, mercury, zinc, iron, and copper have been reported in AD brains, where the abnormal accumulation and distribution of the latter three may elicit oxidative stress and macromolecular damage, impeding cellular function [104–107]. However, considering recent advances in neurobiology of metals, the complex correlation between biometal metabolism, genetics, and the environmental and the pathophysiology of many neurodegenerative diseases [106, 107] warrants further investigation.
In recent years, numerous preclinical studies involving fruits, nuts, and vegetables have identified potential beneficial effects to brain health and cognition resulting from their nutrient and non-nutrient phytochemical content. These phytochemicals have the ability to alter cellular function by modulating transcription factors and altering the expression of genes, cellular metabolism, and cellular signaling [108]. In vitro and in vivo studies have firmly established the mechanistic effects of these phytochemicals on cognitive function, with many studies demonstrating their ability to counter oxidative stress [109], inflammation [110], as well as promote neuronal signaling and regulate transcription [111]. In the past five years, there has been an increasing interest in the role that gut microbiota may play in the modulation of the bidirectional signaling underlying the gut–brain axis [112, 113]. In particular, in an Irish study, there was an association between indicators of general health, including frailty and MMSE, and microbiota composition only in long-stay subjects [114]. It has been also suggested that diet and specific nutrients can affect the composition of the gut microbiota [115] that might influence the production or aggregation of Aβ protein in AD pathogenesis [116]. Bacteria populating the microbiome have been shown to produce amyloids, lipopolysaccharides, and other immunogenic compounds [117, 118] that might contribute to the regulation of signaling pathways implicated in neuroinflammation, brain Aβ deposition, and AD pathogenesis [116]. Plant-derived nutrients and phytocompounds, by ameliorating gut inflammation and dysbiosis, might stimulate a positive modulation of the gut–brain axis, reduce neuroinflammation, and retard or regress cognitive impairments associated with AD [119]. Additionally, high intake of plant foods and specific plant-derived compounds have been shown to reduce Aβ aggregation and, for this reason, are currently considered suitable for the prevention of AD, as showed in some selected studies of the present systematic review [34, 57].
Caffeine, as an antioxidant substance, was revealed to reduce oxidative stress [120] and also to protect against the disruptions of the blood-brain barrier [121]. Most of the biological effects of caffeine, including those on the brain and the central nervous system, are mediated through antagonism of the adenosine receptors, specifically the A1 and A2A receptors [122]. Other complementary mechanisms of caffeine action that could contribute to its cognitive benefits in AD mice are the ability of caffeine to decrease hippocampal levels of pro-inflammatory cytokines [e.g., tumor necrosis factor-α, interleukin (IL)-12(p70), and interferon-γ) [123], caffeine’s beneficial effects on signal transduction factors is involved in neuronal plasticity and survival [124], and the ability of caffeine to enhance brain mitochondrial function [125].
Micronutrients, macronutrients, and late-life cognitive disorders: Possible mechanism models
Optimal micronutrient status would moderate the deterioration in brain integrity. Autophagy is probably a crucial and a major process in the preservation of brain integrity. Micronutrients (vitamins, trace-elements, and also antioxidants) most likely affect brain integrity by normalizing efficient autophagy [126]. Multiple lines of evidence indicate that oxidative stress not only strongly participates in an early stage of AD prior to cytopathology, but it plays an important role in inducing and activating multiple cell signaling pathways that contribute to the lesion formations of toxic substances and then promote the development of AD. Exogenous anti-oxidants (such as vitamin E and C, carotenoids, flavonoids) decrease free radical mediated damage caused by toxic chain reactions in neuronal cells and reduce the toxicity of Aβ in in vitro studies of brains of patients with AD [127].
There is a vast literature demonstrating neuroprotection of the brain by vitamin E, rich sources of which are vegetable oils, nuts, and whole grains [128]. In particular, vitamin E might exert a regulatory effect on cell proliferation and a beneficial effect in improving glucose transport and insulin sensitivity [129]. Most of the known actions of flavonoids are related to their antioxidant properties that can include suppression and scavenging of reactive oxygen species, and upregulation of antioxidant defenses, the control of neuronal survival, death, and differentiation, long-term potentiation, and memory as well as the effect on gene expression and interactions with mitochondria [130]. Therefore, flavonoids, lead to increased NO bioavailability in the vasculature of the hippocampus and subsequent angiogenesis and neurogenesis.
It has now been proven that vitamin B6, vitamin B12, and folate deficiency, as co-factors in the methylation of homocysteine, are associated with increased homocysteine concentrations. Supraphysiological concentrations of homocysteine or deficits in folate and vitamin B12 should promote Aβ and tau protein accumulation and neuronal death, and also have a direct effect on cognitive decline. The potential mechanisms whereby homocysteine might mediate cognitive decline and dementia include: neurotoxicity induced by activation of N-methl- D-aspartate receptors, promotion of apoptosis, vascular injury from promotion of atherogenesis making vascular endothelium prothrombotic and proliferation of smooth muscle cells, platelet activation, and increased burden of ischemic strokes and white matter lesions [131].
Epidemiologic investigations have revealed a beneficial role of vitamin D in muscle function, cardiovascular health, diabetes, and cancer prevention. Low levels of serum 25(OH)D are also associated with increased odds of prevalent cognitive dysfunction, AD and all cause dementia in a number of studies. Vitamin D contributes to neuroprotection by modulating the production of nerve growth, neurotrophin, glial cell derived neurotrophic factor, nitric oxide synthase and choline acetyl transferase [132], and neuroprotective mechanisms including vasoprotection and Aβ phagocytosis and clearance [133, 134].
Several mechanisms have been postulated for the possible protective role of n-3 PUFA in dementia [77]. Supplementation with n-3 PUFA may have cognitive benefits based on animal studies showing neuroprotection through anti-oxidant, anti-inflammatory and anti-amyloid effects. DHA, the predominant n-3 PUFA in the brain, is a key component of membrane phospholipids in the brain and adequate n-3 PUFA status, coming from dietary consumption, may help maintain integrity and neuronal function [135]. Secondly, n-3 PUFAs appear to have effects on dopaminergic and serotonergic systems [136]. The oxidative products of PUFA act as key cellular mediators of inflammation, allergy and immunity, oxidative stress, bronchial constriction, vascular response, and thrombosis, and may thereby influence risk especially for vascular dementia. DHA may be directly involved in enhancing neuronal health in the aging brain through a range of potential mechanisms. In particular, it increases cell viability via neuroprotective and antiapoptotic mechanisms, while also promoting dendritic arborization and synaptogenesis [137]. DHA may modify the expression of genes that regulate a variety of biological functions potentially important for cognitive health, including neurogenesis and neuronal function [138]. Long-chain n-3 PUFA have been shown to reduce Aβ formation and oxidative damage [139]. Higher intakes of saturated and trans fat and lower intakes of mono- and polyunsaturated fat can contribute to insulin resistance and an atherogenic lipid profile [140]. Moreover, insulin resistance, high insulin levels, and cholesterol are all implicated in Aβ accumulation in the brain, the pathologic hallmark of AD [141]. In a very recent neuropathological metabolomic-based study, in the brains of patients with varying degrees of AD pathology (healthy brains, brains with AD pathology but without cognitive symptoms, and AD brains), significant differences in the abundance of six unsaturated fatty acids were identified in three brain regions both vulnerable and resistant to classical AD pathology, with gradations in these metabolites being related to both severity of neuropathology at death as well as domain-specific cognitive performance during life [142]. These findings suggested that dysregulation of unsaturated fatty acid metabolism may play a role in driving AD pathology, providing with further evidence for the metabolic basis of AD pathogenesis.
DISCUSSION
In the last decade, the association between diet and cognitive function or dementia has been largely investigated. In the present article, we systematically reviewed observational studies published in the last three years (2014–2016) on the relationship among dietary factors and late-life cognitive disorders using different levels of investigation (i.e., dietary patterns, foods and food-groups, and dietary micro- and macronutrients). In the present systematic review, considering separately the different cognitive outcomes in older age, we included studies focusing on dementia, AD, MCI, and different models of late-life cognitive impairment/decline, but we did not find studies focusing in particular on VaD. From the reviewed studies, it appeared to be apparent that the NIA–AA guidelines for AD and cognitive decline due to AD pathology introduced some evidence suggesting a direct relation between diet and changes in the brain structure and activity. Several studies focused on the role of the dietary patterns on late-life cognition, with accumulating evidence that higher adherence to a Mediterranean-type diet was associated with decreased cognitive decline, although the MeDi combines several foods, micronutrients, and macronutrients already separately proposed as potential protective factors against dementia and MCI. Moreover, also other emerging healthy dietary patterns, such as the DASH and the MIND diets, were associated with slower rates of cognitive decline and significant reduction in AD rate.
Furthermore, some foods or food groups traditionally considered harmful such as eggs and red meat have been partially rehabilitated, while there is still a negative correlation of cognitive functions with saturated fatty acids and a protective effect against cognitive decline of elevated fish consumption, high intake of MUFA and PUFA, particularly n-3 PUFA. In fact, while a Western diet was associated with worse cognitive performance in several imaging studies selected in the present systematic review [37, 41]; however, for red meat, a classical element of this dietary pattern, the epidemiological findings were controversial [35, 48], also including studies outside the time limits of the present systematic reviews [143, 144]. Multi-country ecological studies of dietary meat and increased risk of cancer [145] have been strongly supported by trends in cancer risk in Asian countries undergoing the nutrition transition [146, 147], as well as in observational studies in Uruguay [148]. Thus, it would be expected that they would also be reliable for AD. In 2014, an ecological study linked the nutrition transition in Japan to the greatly increased risk of AD [149], with increased meat consumption as an important factor. Interestingly, rates of VaD have been relatively unchanged for many years [149]. Recent observational and ecological findings added support for dietary advanced glycation end products as an important risk factor for AD [150], with meat cooked at high temperatures in a dry environment as an important source. Another recent ecological study compared dietary factors with prevalence rates of AD in ten countries with recent prevalence determinations, showing that dietary supply of meat or animal products less milk 5 years before AD prevalence had the highest correlations with AD rates [151]. The three countries with highest rates of AD and dietary meat, Brazil, Mongolia, and the U.S., are considerably different in many respects, adding confidence that the finding was not due to confounding factors [151]. Noteworthy, MeDi falls in the middle of the regression line for meat and AD prevalence [151]. In these ecological studies, countries with high consumption of rice and low consumption of meat had the lowest rates of AD, as did Japan prior to the nutrition transition [149]. Also, eggs and fish added to meat gave similar results as meat [151]. Fish that does not have PUFA n-3 and vitamin D may act like meat and eggs in terms of AD risk.
However, some limitations should be reported for this systematic review article. Heterogeneity exists in the quantification of individual items as well among the different diet backgrounds of the populations investigated, especially in view of different geographical areas, setting of dietary patterns, such as the Mediterranean countries in which a large segment of the population still adhered to MeDi. Heterogeneity in time between the two assessments, among studies using paired assessments (or a single assessment) several years after study population enrollment. Nevertheless, these data represent a brick in the construction of the building of the causal link between dietary habits and cognitive impairment.
The absence of causal etiological therapies against AD leads to seek multimodal alternative strategies, increasing the interest in the potential for prevention of dementia by targeting modifiable risk factors. It is now evident that dietary habits influence diverse cardiometabolic risk factors, including not only obesity and low-density lipoprotein cholesterol, but also blood pressure, glucose-insulin homeostasis, lipoprotein concentrations and function, oxidative stress, inflammation, endothelial health, hepatic function, adipocyte metabolism, pathways of weight regulation, visceral adiposity, and the gut microbiome. Whereas decades of dietary recommendations focused on dietary fat and single vascular risk factors (e.g., hypertension, blood cholesterol, etc.) and current dietary discussions are often worried about total calories and obesity, the full health impact of diet extends far beyond these pathways. Considering strategies of prevention of AD could be complicated and take to negative results. A second key lesson is the importance of pointing out specific foods and overall diet patterns, rather than single isolated nutrients, for cognitive impairment. No single micronutrient or macronutrient, no single antioxidant or vitamin, especially in their synthetic forms, may be able to show a dramatic effect on dementia prevention, due to the complexity of biochemistry. Even a mixture of fixed beneficial compounds applied to everybody will not show dramatic effects against late-life cognitive disorders due to interindividual and intraindividual response. Natural nutrition may exert better effects than single nutrients due to bioavailability and biochemical synergisms. However, also the healthiest natural nutrition alone, such as MeDi, could not exert dramatic effects against cognitive disorders in older age if not part of diet intended as lifestyle. A food-based approach also better facilitates public guidance and minimizes industry manipulation. Nevertheless, the complexity of the stake, the correction of modifiable risk factors to expect “the compression of cognitive morbidity” still remains a desirable goal of public health. Healthy diets and lifestyles could be not very powerful if applied in fixed amounts regardless of individualized multidimensional needs and resources. Larger observational studies with longer follow-up periods should be encouraged, addressing other potential bias and confounding sources, so hopefully opening new ways for diet-related prevention of dementia and AD.
DISCLOSURE STATEMENT
Authors’ disclosures available online (http://j-alz.com/manuscript-disclosures/17-0248r1).