
Abstract
Aging is a risk factor for Alzheimer’s disease (AD). There are changes of brain metabolism and biometal fluxes due to brain aging, which may play a role in pathogenesis of AD. Positron emission tomography (PET) is a versatile tool for tracking alteration of metabolism and biometal fluxes due to brain aging and AD. Age-dependent changes in cerebral glucose metabolism can be tracked with PET using 2-deoxy-2-[18F]-fluoro-D-glucose (18F-FDG), a radiolabeled glucose analogue, as a radiotracer. Based on different patterns of altered cerebral glucose metabolism, 18F-FDG PET was clinically used for differential diagnosis of AD and Frontotemporal dementia (FTD). There are continued efforts to develop additional radiopharmaceuticals or radiotracers for assessment of age-dependent changes of various metabolic pathways and biometal fluxes due to brain aging and AD with PET. Elucidation of age-dependent changes of brain metabolism and altered biometal fluxes is not only significant for a better mechanistic understanding of brain aging and the pathophysiology of AD, but also significant for identification of new targets for the prevention, early diagnosis, and treatment of AD.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative disease accounting for more than 80% of dementia in the aged population. Senile plaques derived from deposits of Amyloid β (Aβ) and neurofibrillarytangles (NFT) of hyperphosphorylated Tau aggregates are two neuropathological hallmarks of AD [1 –4]. Tremendous efforts have been made to explore use of these two biomarkers as a target for prevention, diagnosis, and treatment of AD [5, 6]. Positron emission tomography (PET) is a nuclear medicine technique with high sensitivity for noninvasive detection of Aβ or NFT deposition in vivo. Following administration of a radiotracer, deposits of Aβ or NFT could be visualized by computer analysis of 510 keV gamma rays emitted by the electron–positron annihilation process after binding of a positron emitting Aβ or NFT radiotracer to brain deposits of Aβ or NFT. Several radiopharmaceuticals were developed for noninvasive detection of Amyloid β plaques with PET, including N-methyl-[11C]2-(40 methylaminophenyl)-6 hydroxybenzothiazole [11C]Pittsburgh Compound-B or [11C]PiB [7, 8], and fluorine-18 labeled amyloid β radiopharmaceuticals 18F-florbetapir, (18F-AV-45) [9, 10], 18F-flutemetamol [11, 12], 18F-florbetaben [13, 14]. Recently, there have been significant progression in development of Tau-targeting radiopharmaceuticals for detection of NFT in AD patients with PET, including 11C-PBB3 [15, 16], 18F-FDDNP[17], 18F-THK-523 [18, 19], 18F-THK-5105 [19],18F-THK-5117 [19], 18F-T807, or 18F-T808[20 –22]. AD is a multi-factor disease, and neuroinflammation may play a role in the pathogenesis of AD. There were efforts to develop radiopharmaceuticals targeting 18 kDa translocator protein (TSPO), previously known as peripheral benzodiazepine receptor, on activated microglial cells for assessment of neuroinflammation in AD with PET, such as 11C-(R)-PK11195 (11C-radiolabeled N-butan-2-yl-1-(2-chlorophenyl)-N-methylisoquinoline-3-carboxamide) [23 –25], 11C-PBR28 (N-acetyl-N-(2-[11C]methoxybenzyl)-2-phenoxy-5-pyridinamine) [26, 27], 11C-ER176 (11C-(R)-N-sec-butyl-4-(2-chlorophenyl)-N-methylquinazoline-2-carbo-xamide) [28], 18F-FEPPA (N-Acetyl-N-(2-[18F]fluoroethoxybenzyl)-2-phenoxy-5-pyridinamine)[29], and 18F-FEMPA (N-2-[2-18F-Fluoroethoxy]-5-methoxybenzyl-N-[2-(4-methoxyphenoxy)pyri-dine-3-yl]acetamide) [30]. Investigating biodistribution of 11C-(R)-PK11195 uptake in the brains of healthy children and adults, Kumar et al. demonstrated increase of 11C-(R)-PK11195 uptake with age, without hemispheric asymmetry and change of the overall pattern of brain 11C-(R)-PK11195 uptake [25]. Wiley et al. found no differences in brain 11C-(R)-PK11195 uptake when subjects were grouped by clinical diagnosis or the presence or absence of brain Aβ deposits as indicated by analyses of [11C]Pittsburgh Compound-B uptake with PET [24]. Recently, PET quantitative analysis using new generation TSPO-binding ligands revealed increased uptake of 11C-PBR28 [26, 27], 18F-FEPPA [29], or 18F-FEMPA [30] in the brains of AD patients compared with physiological low brain uptake of these TSPO-binding radiotracers in healthy controls, supporting a longitudinal study with a larger number of human subjects for the role of neuroinflammation in brain aging and pathogenesisof AD.
Aging is one of the major risk factors for development of AD [31]. In addition to age-related morphological changes, such as brain volume loss or atrophy, metabolic changes associated with brain aging play a role in shifting from healthy brain aging to pathological brain aging in AD [32, 33]. Using a radiolabeled metabolite or a substrate specific for an enzyme or product of a metabolic pathway as a tracer, brain metabolic changes could be assessed non-invasively and quantitatively by PET measurement of radiotracer concentration or activity which reflects metabolic activity within a brain structure. Although the spatial resolution of PET is lower than those of computed tomography (CT) and magnetic resonance imaging (MRI), recent development of modern hybrid PET/CT or PET/MRI scanners makes it possible to achieve accurate anatomical localization of radiotracer activity within a specific brain region. Using a PET-CT or PET-MRI scanner, three-dimensional mapping of cerebral radiotracer activity can be accomplished with PET and CT or MRI performed on the patient during the same session. PET/CT or PET/MRI techniques are valuable tools for longitudinal assessment of metabolic changes and altered biometal fluxes in brain aging and AD. Multiple positron emitting radionuclides are available for radiolabeling of analogues of metabolic precursors or a component of a metabolic pathway in the preparation of PET radiopharmaceuticals or radiotracers (Table 1).
Radionuclides for preparation of radiotracers to assess alteration of metabolism and biometal fluxes in brain aging and AD with PET
Age-dependent metabolic changes in brain aging and AD can be assessed with PET non-invasively and quantitatively (Fig. 1). Using a positron emitting radionuclide produced with a cyclotron or generator, radiopharmaceuticals were prepared according to a standard protocol of good manufacturing practice (GMP) production and delivered to an imaging facility for functional imaging of brain metabolic changes with PET. For instance, alteration of glucose metabolism in brain aging and AD can be tracked with PET using 2-deoxy-2-[18F]-fluoro-D-glucose (18F-FDG), a radiolabeled glucose analogue, as a tracer [34 –36]. Following intravenous administration of 18F-FDG, brain glucose metabolic activity can be evaluated by visual assessment of PET images and quantitative analysis of 18F-FDG radioactivity localized within a brain region of interest(ROI).

Schematic representation of non-invasive assessment of age-dependent changes of metabolism and biometal fluxes in brain aging and AD with PET. Using positron emitting radioisotopes produced with a cyclotron or generator, radiopharmaceuticals can be prepared in a cyclotron/radiochemistry facility according to standard good manufacturing practice (GMP) production protocol. Following administration of a radiotracer, biodistribution of the radiotracer within the human body can be analyzed by visual assessment of PET images acquired with a PET or PET/CT scanner. PET quantitative analysis can be performed to determine concentration of the radiotracer within different regions of the human brain. Using radiotracers specific for various metabolic pathways or biometals, age-dependent changes of brain metabolism and biometal fluxes can be determined by altered brain distribution and concentration of the radiotracers with PET. ROI, region of interests. SUV, standardized uptake value.
In this short review, we will discuss advances in the development of radiopharmaceuticals for assessment of age-dependent changes of metabolism and biometal fluxes in brain aging and AD with PET (Fig. 2, Table 2).
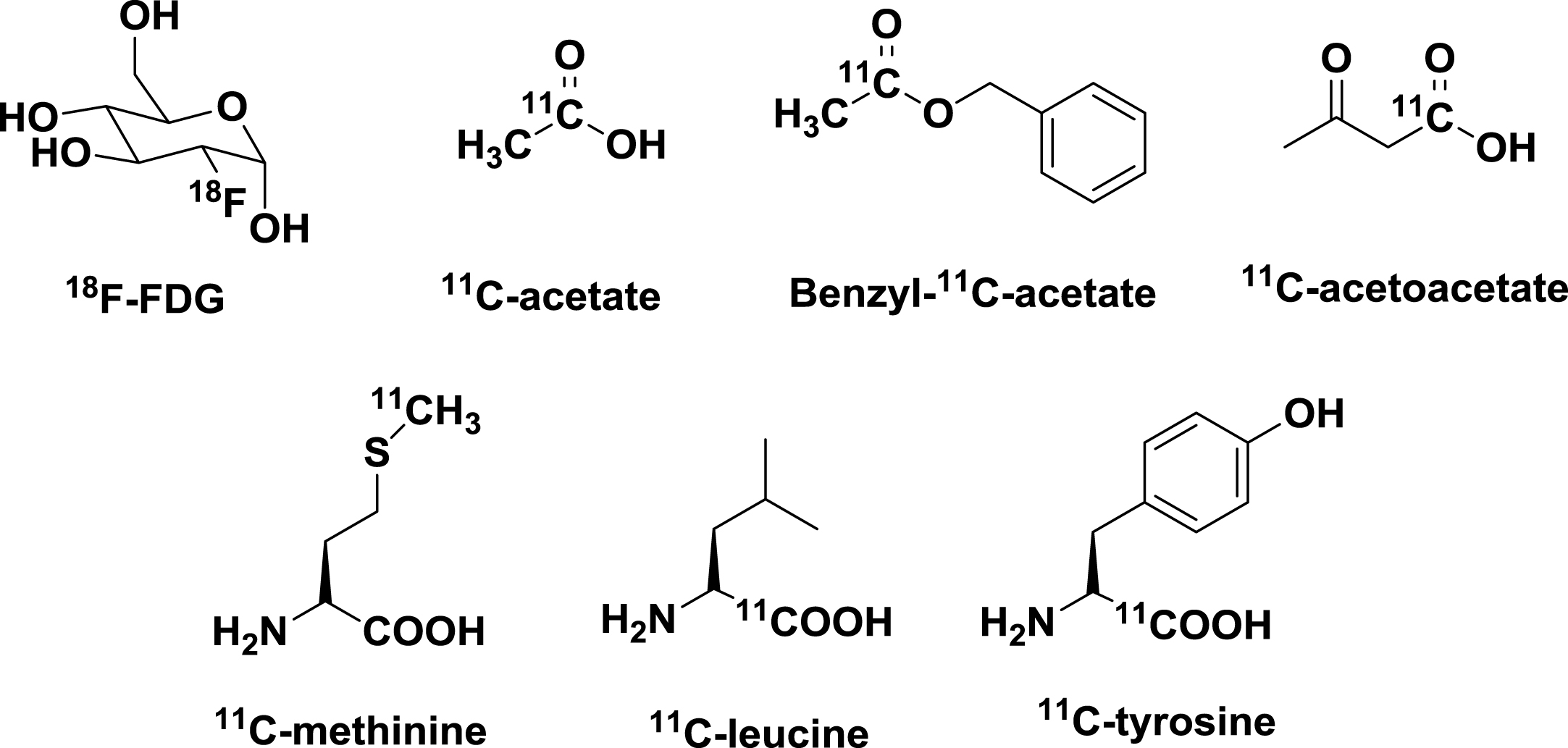
Structure of radiopharmaceuticals for assessment of metabolic changes in brain aging and AD with PET.
Radiopharmaceuticals for assessing alteration of metabolism and biometal fluxes in brain aging and AD with PET
GLUCOSE METABOLISM
There is an alteration of cerebral glucose metabolism in the pathophysiology of AD [35, 36]. The molecular mechanism of glucose metabolic changes in AD is not fully understood, but it is likely related to malfunction of neuron glucose transporters or glycolytic enzymes, or dysregulation of systemic glucose metabolism [37]. Mild cognitive impairment (MCI) is considered as a prodromal stage of AD [38]. Although detection of glucose hypometabolism in the parieto-temporal lobes and frontal lobes often signified advanced AD and loss of opportunity for early intervention, glucose hypometabolism in the posterior cingulate cortex (PCC) could be detected in the early stages of AD with PET [39 –43]. Glucose hypometabolism may be a useful biomarker for anti-AD drug development for selection of human subjects with MCI for clinical trials and tracking therapeutic effects of these agents on glucose metabolic changes with PET.
LIPID METABOLISM
Alteration of lipid metabolism plays a role in the pathogenesis of AD [44, 45]. Apolipoprotein Eɛ4 (Apo-Eɛ4) genotype is associated with increased risk for AD [46, 47]. However, little is known about changes in brain endogenous lipid metabolism in AD, which may be partially related to a lack of radiopharmaceuticals for real-time assessment of neuron lipid metabolism with PET. Acetate is a molecule that enters the tricarboxylic acid cycle in lipid metabolism involving energy production and synthesis of the cell membrane. There were attempts to use radioactive 11C-acetate to study brain energy production and oxidative stress related to brain aging. In 2009, Wyss et al. investigated astrocytic oxidative metabolism using 11C-acetate as a radiotracer and demonstrated stimulation-induced increases of astrocytic oxidative metabolism in rats and humans [48]. In 2013, there was also an attempt to investigate astrocytic oxidative metabolism using benzyl [1-11C]acetate, an acetate analog with better brain uptake, with PET [49]. A longitudinal PET study of age-dependent changes in lipid metabolism in brain aging using 11C-acetate is expected to be informative on the role of altered lipid metabolism in the pathophysiology of AD. The brain meets a portion of its energy requirements from ketone bodies when there is impairment of glucose metabolism. Using radioactive 11C-acetoactate, one of three water-soluble ketone body molecules (acetoacetate, beta-hydroxybutyrate, and acetone), Castellano et al. found normal 11C-acetoacetate metabolism in mild AD dementia patients who had low brain 18F-FDG uptake [50]. Recently, an inverse relationship between brain glucose and ketone metabolism in adults during short-term moderate dietary ketosis was demonstrated with a dual-tracer PET study using 18F-FDG and 11C-acetoacetate [51]. Targeting deficiencies in pyruvate dehydrogenase complex, aconitase, an alpha-ketoglutarate dehydrogenase complex in AD, radiolabeled pyruvate, citrate, or alpha-ketoglutarate molecules might be used as a radiotracer to assess altered brain metabolism in AD with PET. Development of radiopharmaceuticals specific for various pathways of brain lipid and ketone body metabolism is significant for delineation of the role of altered lipid metabolism in the pathophysio-logy of AD.
AMINO ACID TRANSPORT AND PROTEIN SYNTHESIS
Deficits in amino acid transport and protein function play a significant role in the pathogenesis of AD. In addition to radiopharmaceuticals used for evaluation of Aβ and Tau molecules, there are a few amino acid-based radiopharmaceuticals which might be useful for evaluation of altered amino acid transport and protein synthesis in brain aging and pathophysiology of AD with PET(Fig. 2).
To test the hypothesis that local rate of perikaryal protein synthesis can be used as a biomarker for assessment of neuronal loss in AD, Salmon et al, used [11C]methionine as a radiotracer to study changes in amino acid transport and protein synthesis in AD, in conjunction with the use of 18F-FDG for assessment of altered glucose metabolism with PET [52]. It was found that there was about a 20% decrease in [11C] methionine accumulation in the temporo-parietal lobe in AD patients compared with normal controls, along with a 45% decrease in temporo-parietal lobe glucose metabolism in AD. In contrast, there was no significant difference for [11C]methionine radiotracer activity in the cerebellum between AD patients and normal controls. No correlation of a regional decrease in [11C]methionine accumulation with severity of AD symptoms was observed. Two amino-acid based radiotracers, L-[11C]-tyrosine and L-[1-11C]-leucine, were used for studies of protein synthesis in the brain [53, 54]. However, there was no report of using these two radiotracers for evaluation of altered amino acid transport or protein synthesis in AD. It would be interesting to test whether L-[11C]-tyrosine or L-[1-11C]-leucine can be used as a radiotracer for assessment of altered amino acid transport and protein synthesis in brain aging and AD. L-arginine, a semi-essential amino acid, is a precursor of nitric oxide produced by NO synthesis. It was reported that over-expression of human wild type beta amyloid precursor protein (βAPP) caused changes in cellular L-arginine metabolism [55]. It remains to be determined whether altered metabolism of L-arginine or other amino acids play a role in Aβ-related pathology in AD. In view of the important role of amino acid transport and protein synthesis in brain function, there is an urgent need for dedicated efforts to develop more amino acid-based radiotracers to assess age-dependent changes of amino acid transport and protein synthesis in brain aging and the pathogenesis of AD.
COPPER
Copper is a transition metal essential for brain development and function, but excess copper may be harmful. There has been significant progress in the delineation of a delicate network of copper chaperones and transporters which function together to maintain cellular copper homeostasis. However, there is limited understanding of the systemic regulation of copper fluxes to maintain brain copper homeostasis, which is partly due to lack of a tool for real-time tracking of systemic and brain copper fluxes. It was postulated that disruption of copper homeostasis might play a role in the pathogenesis of AD [56, 57]. PET is a useful tool for real-time tracking of brain copper fluxes or trafficking using a radioactive copper tracer. It was demonstrated that radioactive glyoxalbis(N4-methyl-3-thiosemicarbazonato)-64Cu(II) complex, 64Cu(II)-GTSM, could cross the blood brain barrier (BBB) and release 64Cu after intracellular reduction of 64Cu(II)-GTSM to an unstable 64Cu(I)-GTSM [58]. Using 64Cu(II)-GTSM as a probe, Torres et al. examined changes to brain copper trafficking in TASTPM transgenic AD mice carrying both the Swedish mutation in APP and the M146V mutation in PSEN1 with PET [59]. Altered 64Cu trafficking in the brains of TASTPM transgenic AD mice was demonstrated by detection of increased 64Cu concentration and faster brain 64Cu clearance, compared to age-matched wild type controls. There was heterogeneous 64Cu distribution within the brains of TASTPM transgenic mice, but regional brain distribution of 64Cu did not correlate with amyloid-β plaque distribution based on the results of ex vivo autoradiography conducted at 24 h post injection of 64Cu(II)-GTSM. The findings suggest potential application of 64Cu-GTSM as a radiopharmaceutical for diagnostic imaging of AD with PET. Although an increase of intracranial delivery of 64Cu with 64Cu-GTSM made it possible to obtain PET images for visual assessment of intracranial copper trafficking, it remains to be determined whether altered distribution and clearance of 64Cu observed in the brains of TASTPM transgenic AD mice injected with 64Cu-GTSM reflected brain copper trafficking when brain copper fluxes are regulated by a network of copper transporters and chaperons [60, 61]. There is a need for new radiotracers which can be used for assessment of altered copper fluxes in brain aging and AD under systemic regulation of copper fluxes, including gastrointestinal absorption and whole body biodistribution of copper, and subsequent control of copper fluxes to maintain brain copper homeostasis. Copper-64 chloride (64CuCl2) was tested as a radiotracer for assessing copper metabolism imbalance in Wilson’s disease (WD) with PET, using an Atp7b –/– knockout mouse model of WD [62, 63]. Age-dependent changes of brain copper fluxes in Atp7b –/– knockout WD mice was demonstrated by a longitudinal PET study using orally administered 64CuCl2 as a radiotracer [64]. Moreover, 64CuCl2 was successfully used as a radiotracer for diagnostic imaging of prostate cancer [65] and brain tumors [66] in humans with PET, based on previous use of 64CuCl2 as a tracer for localization of human prostate cancer xenograft tumors in rodents [67]. The findings from these studies support use of 64CuCl2 as a radiotracer for quantitative analysis of age-dependent changes of copper fluxes in brain aging and pathophysiology of AD with PET[68 –70].
IRON
Iron is one of the most abundant metals in the human body [71]. Iron is essential for maintenance of dopaminergic and GABAergic neurotransmission [72], but an excess of ferrous iron may be harmful due to generation of reactive oxygen species [73]. Bruehlmeier et al. used 52Fe-citrate as a tracer for tracking iron metabolism in WD and demonstrated significantly increased brain 52Fe-transferrin uptake in WD patients following rapid conversion of intravenously injected 52Fe-citrate to 52Fe-transferrin immediately after intravenous administration [74]. There is an increase of iron in the brain with aging [75], and increased iron levels were observed along with deposits of senile plaque in AD. It was found that 52Fe increased significantly in the temporal regions of 8 AD patients compared with 52Fe in the brains of 10 age matched healthy controls [76]. It was postulated that iron might serve as a catalyst for free radical formation inducing aggregation of Aβ [77]. A longitudinal study of age-dependent changes of iron metabolism in brain tissue with PET is expected to be informative in delineation of the iron’s role in the pathogenesis of AD.
ZINC
Zinc is a metal required for brain function [78]. Zinc levels stay relatively constant in the brain throughout adult life. There are several zinc transporter families which function to maintain zinc homeostasis [79]. Zinc is released into the synaptic cleft, where it modulates N-methyl-D-aspartate receptor (NMDA) and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors [80]. Altered concentration of zinc in the brains of AD patients was reported, with reduction of zinc concentration in hippocampus, amygdala, and inferior parietal lobe compared with zinc concentration in the brains of control subjects. Radioactive 63Zn-zinc citrate was used as a radiotracer for tracking changes in zinc transport in vivo. A biodistribution study of 63Zn revealed prominent uptake of 63Zn in the liver (about 50% dose/organ), along with moderate 63Zn uptake in the pancreas, kidney, spleen, and intestines and low uptake in bone marrow and the brain. There was no significant difference of 63Zn uptake in the brains of AD patients and healthy controls. However, kinetic analysis demonstrated low clearance of 63Zn (27.5 to 97.5 min) in multiple brain regions in AD patients compared to clearance of 63Zn in the brains of healthy control group (p < 0.05). The regions of low 63Zn clearance in AD include cingulate, precuneus, left and right prefrontal cortices, left parietal, left lateral temporal, orbitofrontal cortex, primary visual cortex, and the occipital cortex. Interestingly, the brain regions with slow 63Zn clearance corresponded to the regions of amyloid-β deposits visualized on the11C-PIB-PET scans [82]. Although the number of human subjects was small, the findings of this study invite further investigation of altered zinc homeostasis in brain aging and the pathophysiology of AD, in conjunction with assessment of altered homeostasis of other biometals in brain aging and AD.
MANGANESE
Manganese (Mn) is a micronutrient metal, but excess manganese is neurotoxic. It was found that the serum Mn levels were lower in AD patients compared with the serum Mn levels in healthy controls [83]. Mn deficiency may be a risk factor for AD. It was reported that positron emitting [52Mn]MnCl2 radiotracer was prepared with a cyclotron, and biodistribution of this radionuclide in mice was determined by quantitatively analysis with PET [84]. The findings from this proof-of-concept study suggested that [52Mn]MnCl2 might be a useful radiotracer for study of biodistribution and neurotoxicity of manganese in human body with PET. This may be informative to determine whether there is alteration of dynamic flow of Mn in AD patients and whether altered Mn fluxes play a role in the pathogenesis of AD.
PERSPECTIVES
Although the cause of AD is not fully understood, aging is a significant risk factor for AD and metabolic changes associated with brain aging play a role in the pathogenesis of AD. There are advances in developing 18F-FDG and several other radiopharmaceuticals for assessment of metabolic changes in brain aging and the pathophysiology of AD. However, there is an unmet need of radiopharmaceuticals for assessing changes in various metabolic pathways in brain aging and AD, particularly metabolic changes and altered biometal fluxes related to neurotransmission, function of synapses, and neuron plasticity. It is important to develop radiopharmaceuticals and conduct a longitudinal study to assess age-dependent changes of brain metabolic pathways and biometal homeostasis related to cognition and memory function. Elucidation of the role of metabolic changes and altered biometal fluxes in brain aging and pathogenesis of AD is critical for identification of therapeutic targets and development of effective agents for prevention, early diagnosis, and treatment of AD.