
Abstract
The pathological aggregation of tau is a common feature of most of the neuronal disorders including frontotemporal dementia, Parkinson’s disease, and Alzheimer’s disease. The inhibition of tau aggregation is considered to be one of the important strategies for treating these neurodegenerative diseases. Curcumin, a natural polyphenolic molecule, has been reported to have neuroprotective ability. In this work, curcumin was found to bind to adult tau and fetal tau with a dissociation constant of 3.3±0.4 and 8±1 μM, respectively. Molecular docking studies indicated a putative binding site of curcumin in the microtubule-binding region of tau. Using several complementary techniques, including dynamic light scattering, thioflavin S fluorescence, 90° light scattering, electron microscopy, and atomic force microscopy, curcumin was found to inhibit the aggregation of tau. The dynamic light scattering analysis and atomic force microscopic images revealed that curcumin inhibits the oligomerization of tau. Curcumin also disintegrated preformed tau oligomers. Using Far-UV circular dichroism, curcumin was found to inhibit the β-sheets formation in tau indicating that curcumin inhibits an initial step of tau aggregation. In addition, curcumin inhibited tau fibril formation. Furthermore, the effect of curcumin on the preformed tau filaments was analyzed by atomic force microscopy, transmission electron microscopy, and 90° light scattering. Curcumin treatment disintegrated preformed tau filaments. The results indicated that curcumin inhibited the oligomerization of tau and could disaggregate tau filaments.
INTRODUCTION
The formation of neurofibrillary tangles with tau aggregates is a characteristic of several neurodegenerative diseases like Alzheimer’s disease [1 –3], frontotemporal dementia, Pick’s disease, and progressive supranuclear palsy [4]. These pathological conditions are termed as tauopathies. Tau has a crucial role in microtubule formation and their stabilization [2 , 5–8]. Microtubules are the main component in human neuronal cells and play important roles in axonal transport and growth of axon. Tau binds to microtubules by its three or four microtubule binding regions present at theC-terminus of the protein. Several post-translational modifications including phosphorylation, acetylation, glycation, ubiquitylation, sumoylation, and nitration are reported to regulate tau function [9]. For example, the microtubule-binding ability of tau is highly dependent on its phosphorylation level [10]. The longest isoform of tau has 80 potential phosphorylation sites with mostly serine and threonine residues [11]. A recent study revealed that the acetylation regulates the tau binding to microtubules. The acetylation neutralizes the charges on the microtubule binding region of tau, which interferes with the microtubule and tau interaction, therefore leading to pathological tau aggregation [12]. Tau acetylation on Lys 280 hinders the tau-microtubule interactions, and cytosolic tau is available for aggregation [13]. In neurodegenerative disorders, the hyper-phosphorylated or acetylated tau detaches from the microtubules, changes its structure and aggregates to form paired helical filaments (PHFs) and subsequently neurofibrillary tangles. In addition, some stress molecules, such as amyloid-β (Aβ), are reported to redistribute tau into dendrites and spines, causing a toxic effect on synapse [14]. This leads to microtubule breakdown, defects in intracellular trafficking, and disruption of the dendritic spine [15].
Several therapeutic approaches have been reported for treating neurodegenerative diseases, especially tauopathies, which include tau phosphorylation inhibitors [11], microtubule stabilizers [16], tau aggregation inhibitors [1 , 18], and immunotherapy [19]. The present study examines the interaction of a natural polyphenolic compound, curcumin (diferuloylmethane), with tau. Curcumin is found in the rhizome of turmeric, Curcuma longa L. (Zingiberaceae), and it has antioxidant, anti-angiogenic, anti-inflammatory, and neuroprotective properties [20]. It reduces the oxidative damage and amyloid pathological symptoms in an Alzheimer’s transgenic mouse [21]. It inhibits both the oligomerization and fibrilization of Aβ protein and induces disaggregation of Aβ in transgenic mice with Alzheimer’s disease [22]. Further, curcumin was reported to inhibit α synuclein aggregation and to increase the solubility of the protein [23]. It decreased the level of soluble tau dimers and elevated heat shock proteins involved in tau clearance in transgenic mice [24]. The heat shock proteins, HSP 70 and HSP 90, were reported to enhance tau solubility and promote tau-microtubule interaction [25]. However, the effect of curcumin on the aggregation of tau is not known. In this work, we report that curcumin binds to adult tau more strongly than the fetal tau. Curcumin inhibited the aggregation and fibrilization of tau in vitro. The study also provides a possible mechanism of the inhibition of tau aggregation by curcumin.
MATERIAL AND METHODS
Materials
Piperazine-1,4-bis(2-ethanesulfonic acid)(Pipes); 4-(2-hydroxyethyl)-1-piperazineethanesulfonicacid (HEPES); phenylmethanesulfonyl fluoride (PMSF); ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid (EGTA); 1,4-dithiothreitol (DTT); β-mercaptoethanol (β-ME); curcumin; thioflavin S, Isopropyl β-D-1-thiogalactopyranoside (IPTG); and dimethyl sulphoxide (DMSO) were purchased from Sigma-Aldrich. Arachidonic acid (AA) was purchased from MP Biomedical.
Expression and purification of recombinant tau
4R0N tau used in this study contains 383 amino acids, and it is predominantly found in Alzheimer’s disease PHFs, with other tau isoforms [26, 27]. 4R0N tau is expressed in the adult human brain while 3R0N tau is expressed in fetal brain [2]. 4R0N tau contains 4 microtubule-binding repeats (MTBR), whereas 3R0N tau has 3 MTBR repeats [2]. 3R0N tau does not contain the second (R2) repeat domain [2]. The 4R0N and 3R0N tau expression clones were kind gifts from Prof. S. Feinstein, University of California Santa Barbara. Both proteins were expressed in an E. coli BL-21 DE-3 strain and purified as described previously with minor modifications [7, 8]. Briefly, bacteria were grown in LB media with 100 μg/mL ampicillin for 16 h. This primary culture was inoculated to 1 L fresh LB media with antibiotics, and the cells were grown at 37°C in an incubator shaker. After the O.D.600 reached 0.8–1.0, the culture was incubated with 0.5 mM of IPTG at 37°C for 16 h. Cells were harvested and homogenized in PEM buffer (pH 6.8, 50 mM PIPES, 1 mM EGTA, 1 mM MgCl2, 2 mM PMSF, 0.1% 2-mercaptoethanol, 1X protease inhibitor cocktail, 1 mg/mL lysozyme). The suspension was sonicated using a probe sonicator at 40% amplitude with pulse 2 s on/2 s off. Each sonication cycle was 1 min, and 12 cycles were given to lyse the cells. The lysate was boiled in a water bath for 15 min, and then centrifuged at 26,234× g. Next, the clear supernatant was applied to a phosphocellulose column pre-equilibrated with PEM buffer. Then, the column was washed with PEM buffer, and the protein was eluted with a step gradient of NaCl (0.2 M–0.8 M). Tau was found to elute at 0.3 M NaCl. The purified fractions were pooled and concentrated using 10 kDa cut off protein concentrator and dialyzed in HBSE buffer (pH 7.4, 10 mM HEPES, 100 mM NaCl, 3 mM EGTA, 1 mM DTT) overnight. The purity of the protein was analyzed by SDS-PAGE. The extent of purity was ≥95%. The concentration of tau was determined by UV absorption at 280 nm [28, 29]. Tau was stored at –80°C in small aliquots.
Determination of the dissociation constant of tau-curcumin interaction
4R0N tau (5 μM) in 50 mM PIPES buffer (pH 6.8) was incubated with different concentrations (0.1, 0.2, 0.5, 1, 2, 3, 5, 7, 10, 15, 20, 25, and 30 μM) of curcumin at 25°C for an h. 3R0N tau (5 μM) was incubated with different concentrations (1, 2, 3, 5, 7, 10, 15, 20, and 25 μM) of curcumin at 25°C for an h. Then, the fluorescence spectra (450–550 nm) were monitored by exciting the samples at 425 nm using a 0.3 cm path length cell. The observed intensities were corrected for the inner filter effect using the following equation:
Tau aggregation reaction
In the preliminary experiments, Tau (5 μM) in 10 mM HEPES (pH 7.4), 100 mM NaCl, 5 mM dithiothreitol (HBSE buffer) was incubated without and with either 20 μM curcumin or Methylene Blue for 20 min at 37°C. AA (reconstituted in 100% ethanol) was used to induce the aggregation of tau [30, 31]. The aggregation of tau was induced by adding 75 μM AA. Aggregation reaction had 4% ethanol as a vehicle; hence, the control samples without AA were adjusted to 4% ethanol. All reaction samples were incubated at 37°C for 24 h. Then, the aggregation of tau was monitored by Thioflavin S fluorescence. Thioflavin S (20 μM) was added to the reaction mixtures and the fluorescence spectra were monitored using 440 nm as the excitation wavelength. The fluorescence intensities at 521 nm were noted. The aggregation of tau was also monitored by 90° light scattering at 400 nm using a JASCO fluorescence spectrophotometer.
To determine the half-maximal concentration (IC50) of curcumin required to inhibit tau aggregation, tau (5 μM) in HBSE buffer was incubated without or with different concentrations of curcumin (1, 5, 8, 10, 15, 20, and 30 μM) for 20 min. The aggregation was induced using 75 μM AA for 24 h. The aggregation was monitored by ThS assay and 90° light scattering as mentioned above. The IC50 value of curcumin was calculated using Graph Pad Prism 5 software.
Dynamic light scattering
Curcumin in HBSE buffer (pH, 7.4) was filtered by syringe filter (0. 22 μm, Millipore) to remove undissolved curcumin particles. The concentration of curcumin was then determined by measuring absorbance at 425 nm. Tau (5 μM) was incubated without or with 20 μM curcumin for 20 min. Then, the reaction mixtures were incubated with 75 μM AA for 24 h at 37°C. The aggregation controls were only tau (5 μM) and tau (5 μM) in a presence of AA (75 μM). The DLS measurements of samples were performed with a DLS particle size analyzer (Brookhaven 90 plus). Samples were thermally equilibrated at 25°C for 5 min. Particle size was obtained as an average of four independent experiments with 25 runs each and expressed as numberdistribution.
Effect of curcumin on the rate of tau aggregation
Tau (5 μM) in HBSE buffer (pH 7.4) was incubated without or with 20 μM curcumin for 20 min. Then, the tau was incubated with 75 μM AA at 37°C. At different time intervals (0, 2, 6, 9, 25 h), an aliquot of the sample was withdrawn, and 20 μM Thioflavin S was added to the reaction mixture. Then, the fluorescence of Thioflavin S was monitored. The experiment was performed three times.
Circular dichroism (CD) spectroscopy
Tau (10 μM) in HBSE buffer (2 mM HEPES,pH 7.4, 50 mM NaCl, 1 mM EDTA, 3 mM DTT) in the absence and presence of 20 μM curcumin was incubated without or with 75 μM AA at 37°C for 24 h. The far-UV (200–260 nm) CD spectra were monitored using 1 mm path length quartz cuvette in a J-1500 spectropolarimeter (Jasco, Tokyo, Japan). Each spectrum reported was an average of 5 scans. The experiments were repeated three times. The spectra were deconvoluted using Jasco protein secondary structure analysis version 2.1.1.1.
Tau disaggregation reaction
In order to determine the effect of curcumin on tau disaggregation, tau (5 μM) in HBSE buffer(pH 7.4) was incubated with 75 μM of AA at 37°C for 24 h. The preformed tau aggregates were then incubated with different concentrations (0, 5, 10, 20 μM) of curcumin at 37°C for 3 h. Then, the sample was incubated with Thioflavin S for 5 min, and the fluorescence spectra (460–600 nm) were recorded using an excitation wavelength of 440 nm.
Atomic force microscopy
Tau fibrils were formed when tau was incubated with heparin for several days [32]. We also did not find large tau filaments when 4R0N tau was incubated with AA for 1–3 days. Therefore, tau was incubated with AA for 16 days to form large filaments. Tau (20 μM) was incubated with 75 μM AA in HBSE buffer (pH 7.4) in the presence and absence of 20 μM curcumin for 16 days. The reaction mixtures were incubated at 37°C for 24 h to obtain tau oligomers while the reaction mixtures were incubated for 16 days to prepare large tau filaments. The sample was adsorbed on freshly cleaved atomic force microscopy grade mica sheet for 10 min. Then, the sample was washed thrice with milli-Q water. Images were taken using Asylum MFP-3D Bio-AFM. The heights of oligomers/filaments were measured by Asylum MFP-3D Bio-AFM analysissoftware.
For the de-filamentation reaction, tau (20 μM) in HBSE (pH 7.4) buffer was incubated with 75 μM AA at 37°C for 16 days to obtain filaments. The preformed tau filaments were subsequently incubated with 20 μM curcumin at 37°C for 24 h then; the samples were viewed using atomic force microscopy. The de-filamentation was quantified by counting a number of filaments per field from 10 individual fields.
Electron microscopy
Tau (20 μM) in HBSE (pH 7.4) buffer was incubated with 75 μM AA at 37°C for 16 days in the presence and absence of 20 μM curcumin. The protein sample was placed on the 200-mesh copper grid and allowed to adsorb for 10 min. Then grid was stained with 2% uranyl acetate for 60 s and washed thrice with milli-Q water. Samples were viewed using the JEOL JEM-1220 electron microscope at 120 kV. For the de-filamentation reaction, the preformed tau filaments (as mentioned above in the atomic force microscopy analysis) were incubated with 20 μM curcumin at 37°C for 24 h. Then, the samples were analyzed using an electron microscope. The disaggregation was quantified by counting a number of filaments per field from 10 individualfields.
Homology modeling of tau
The sequence of tau protein with accession number NP_058518.1 was obtained from the NCBI database. The homology model of tau was generated usingI-TASSER online protein model building server[33, 34]. The templates used by I-TASSER for automated homology modeling were Yeast fatty acid synthase (2pff, sequence identity 15%), RHDV capsid protein (3zue, sequence identity 22%), SMN protein (4nl6, sequence identity 13%), tau (267–312) protein (2mz7, sequence identity 12%), Properdin (1w0r, sequence identity 20%), B19 parvovirus capsid (1s58, sequence identity 22%), and TLR domain (1ziw, sequence identity 16%). The server generated a large ensemble of structural conformations at the initial stages of modeling. Subsequently, clustering of all conformations was performed based on structural similarity by SPICKER program built in I-TASSER server. Finally, the server created five homology models of tau, which were representatives of five structure clusters. The structures generated by I-TASSER were analyzed on the basis of confidence score (C-score) and the estimated TM-score. Generally, the C-scores of a predicted model are in the range of –5 to 2, where higher values represent models with higher confidence. The model of tau with the best C-score (–0.68), TM-score (0.63±0.14) and cluster density (0.2503) was selected for furtherstudies.
Molecular docking of curcumin on tau
The three-dimensional coordinates of curcumin with identity number 969516 were obtained from PubChem database. The covalent structure of curcumin is shown in Fig. 1A. The best homology model (C-score = –0.68) of tau generated by I-TASSER server was used for molecular docking. V255A, V292A and double mutant (V255A and V292A) of tau were created separately in silico using COOT [35]. First, the tau model and curcumin structure were converted into the pdbqt format using MGL tools 1.5.6 [36]. Polar hydrogens and gasteiger charges were added to tau and curcumin structure while converting the coordinates to pdbqt format. The molecular docking tool Auto Dock vina [37] was used to dock curcumin on tau. Initially, blind docking of curcumin on wild-type tau was performed. For blind docking, the entire tau molecule was covered in a grid box with dimensions of 92 Å×80 Å×70 Åwith a grid spacing of 1 Å, keeping tau as a rigid and curcumin as a flexible molecule. Five sets of blind docking were performed with 100 exhaustiveness. Each set of autodock vina produced 9 docked conformations, among which 7 conformations were bound to one site of tau and that particular site was chosen for the local docking. The local docking was performed with both wild type and mutant tau models to obtain the correct orientation of curcumin at the binding pocket. For local docking, a grid box having dimensions of 24 Å×40 Å×36 Åwith a grid spacing of 1 Åwas centered at the position of 89.10 Å×80.77 Å×90.04 Åto cover binding site obtained by blind docking. Five sets of docking were performed, and the docked conformations with minimum binding energies were selected (Supplementary Table 1). The obtained data were analyzed by MGL tools 1.5.6 [36]. The interaction of curcumin with tau residues was analyzed using PyMol [38].
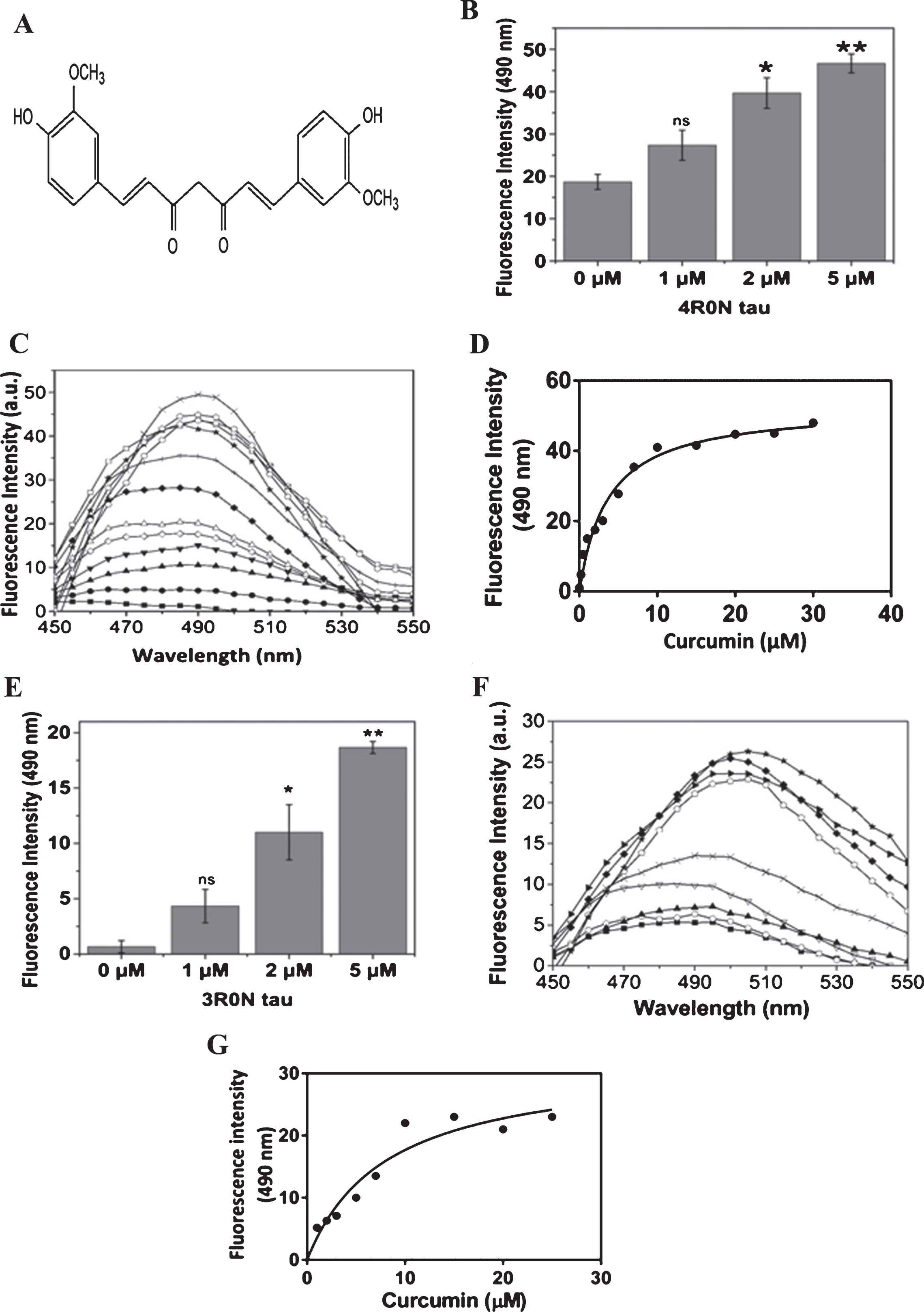
Curcumin binds to recombinant tau. A) The structure of curcumin. B) 4R0N tau (1, 2, 5 μM) was incubated with curcumin (1 μM) for an hour and the binding was monitored using curcumin fluorescence. The fluorescence was noted at 490 nm (n = 4, ns: non-significant, * p < 0.05 and ** p < 0.01). C) The fluorescence spectra of 0.1 (■); 0.2 (●); 0.5 (▴); 1 (▾); 2 (◊); 3 (Δ); 5 (♦); 7 (+); 10 (□); 15 (★); 20 (⌂); 25 (○); and 30 (×) μM of curcumin in the presence of 5 μM of 4R0N tau are shown. D) The change in the fluorescence intensity of curcumin in the presence of 4R0N was plotted against curcumin concentrations. (n = 3). E) 3R0N tau (1, 2, 5 μM) was incubated with curcumin (1 μM) for an hour and the binding was monitored using curcumin fluorescence. The fluorescence was determined at 490 nm (n = 4, ns: non-significant, * p < 0.05 and ** p < 0.01). F) The fluorescence spectra of 1 (■); 2 (○); 3 (▴); 5 (▿); 7 (×); 10 (►); 15 (♦); 20 (⌂); and 25 (★) μM of curcumin in the presence of 5 μM of 3R0N tau are shown. G) The change in the fluorescence intensity of curcumin in the presence of 3R0N tau was plotted against curcumin concentrations. (n = 3).
The 3-dimensional structure of PE859 [39] was generated using Chemdraw. For blind docking, the entire tau molecule and PE859 were covered in a grid box with dimensions of 92 Å×74 Å×72 Åwith a grid spacing of 1 Åand exhaustiveness of 100. Among 9 docked conformations, 8 conformations were docked at MTBR site of tau. For local docking, a grid box having dimensions of 38 Å×40 Å×40 Åwith a grid spacing of 1 Åwas centered at the position of 80.227 Å×80.452 Å×80.568 Åto cover the binding site indicated by the blind docking. Five sets of docking were done and the docked conformation with minimum binding energy was used to determine tau residues interacting with PE859. The analysis of PE859 docking on tau was done using MGL tools 1.5.6 [36].
Statistical analysis
Graphs were plotted using either Graph Pad Prism or Origin Pro 8.0. The results were represented as±SEM. The significance of the data was determined by unpaired t-test where p < 0.05 was taken as significant.
RESULTS
Curcumin binds to tau in vitro
The binding of curcumin (Fig. 1A) to 4R0N tau and 3R0N tau was determined by monitoring curcumin fluorescence (Fig. 1). The fluorescence intensity of curcumin enhanced with increasing concentration of 4R0N tau, indicating that curcumin binds to tau (Fig. 1B). In a separate experiment, a fixed concentration of 4R0N tau was incubated with different concentrations of curcumin (Fig. 1C). In the presence of 4R0N tau, the fluorescence intensity of curcumin increased with increasing concentration of curcumin. The increase in the fluorescence intensity of curcumin upon binding to tau was plotted in Fig. 1D. A dissociation constant for the interaction of curcumin and 4R0N tau was estimated to be 3.3±0.4 μM. The binding of curcumin to 3R0N tau was also similarly examined using the increase in the fluorescence of curcumin upon binding to 3R0N tau (Fig. 1E). The dissociation constant for the binding of curcumin to 3R0N tau was determined to be 8±1 μM (Fig. 1F and G). The binding data suggested that curcumin binds more strongly to 4R0N tau than 3R0N tau. Since 3R0N tau lacks the R2 repeat from the MTBR domain, the finding indicated a possible role of MTBR region in the binding of curcumin to tau.
Curcumin binds to microtubule binding region of tau
The crystal structure of tau is not determined yet. Hence, homology modeling of 4R0N tau was done using I-TASSER server. The predicted tau structure is dominated by the random coil, but it also has 11 helices and 4 β strands (Fig. 2A). The N-terminal region consists of the 2 β strands and 5 helices while C-terminal part consists of 3 helices. The MTBR region consists of 3 helices and 2 β strands. Also, the P148 to R151 (PGSR) part of 4R0N tau forms helical turn conformation same as full length tau model as mentioned earlier [40]. The microtubule binding region (MTBR) of 4R0N tau consists of 116 amino acids that starts from D194 and ends at G309. The MTBR region consists 3 helices and 2 β strands. Five sets of blind docking were performed. In the 80% docked models, curcumin docked in the MTBR region of tau with moderate binding energy (–8.0 to –6.9 kcal/mol), indicating that the MTBR region might be the putative binding site for curcumin.A docking analysis of a new curcumin derivative, PE859 [39], on tau indicated that the compound may bind to the MTBR region of tau with a binding energy of –9.7 kcal/mol (Supplementary Figure 1A). The residues of tau, which are interacting with PE859, are shown in Supplementary Figure 1B. The putative binding pocket of curcumin is dominated by a random coil structure (Fig. 2A). An analysis of the curcumin binding pocket of tau revealed that the binding of curcumin is supported by hydrophobic and polar residues of tau (Fig. 2B). K285, D194, D225, and S258 residues of tau form hydrogen bonds with curcumin. The –NH group from the main chain of D194 forms a strong hydrogen bond with third ketone oxygen of hepatadine chain of curcumin, while –COO group from the side chain of the same residue forms polar interaction with oxygen from methoxyphenyl group. The carboxylic oxygen from the side chain of D225 acts as a proton acceptor with both 3rd and 5th ketone oxygen of heptadine chain of curcumin and forms a hydrogen bond. The 5th ketoneoxygen from heptadine chain forms a hydrogen bond with the – NH group from the side chain of K285. The hydroxyl oxygen of S258 acts as a proton donor for methoxyphenyl group of curcumin and forms a hydrogen bond with the oxygen of methoxy group. The V255, V292, L195 and V305 residues form hydrophobic interactions with curcumin (Fig. 2B). Here L195, V255, and V305 form the hydrophobic interaction with methoxyphenyl rings of curcumin while heptadine ring forms hydrophobic interactions with V292. The binding site of curcumin in tau was further analyzed by performing several in silico mutations in the curcumin-binding pocket of tau. In silico, V255A, V292A, and double (V255A and V292A) mutations were introduced in tau. Then, curcumin was docked on the structures of these mutants (Fig. 2C-E). The putative binding sites of curcumin on wild-type tau and tau mutants were found to be similar (Fig. 2F). The binding energy of curcumin to different tau mutants indicated that curcumin has less affinity towards these variants as compared to the native tau (Supplementary Table 1). These findings indicated that curcumin may bind to the MTBR region of tau (Fig. 2B, G). The MTBR region of tau is considered to be important in the aggregation process [41]. The MTBR regions were stacked upon each other to form β sheet structure. The binding of curcumin to MTBR region may inhibit the stacking of MTBR region and suppress the initial step of aggregation.

A putative binding site of curcumin in tau. A) Homology model of tau represented as cartoon and curcumin is represented as a stick model. The semi-transparent surface of the molecule is also shown in gray color. B) A zoomed view of the curcumin binding site in tau. Curcumin is represented as ball and stick model. The residues of tau are shown as sticks. Hydrogen bonds are shown as black lines while hydrophobic interactions are depicted as a purple dotted surface. C-E) The models showing the interaction of curcumin with V292A mutant (C);V255A mutant (D); double mutant (V255A & V292A) (E). F) The model showing that the putative binding site of curcumin with wild-type tau and tau mutants is same. G) The tau schematic showing the domains of tau and residues interacting with curcumin.
Curcumin inhibits the aggregation of tau
The effect of curcumin on the aggregation of tau was monitored using multiple approaches. In addition, the effect of curcumin on the aggregation of tau was monitored using Thioflavin S dye and 90° light scattering (Fig. 3). Thioflavin S is known to bind to the β sheet of aggregated proteins and produces fluorescence at 521 nm [42]. As reported earlier [43], methylene blue was found to inhibit the aggregation of tau as monitored by Thioflavin S fluorescence (Fig. 3A) and 90° light scattering (Fig. 3B). DMSO had no discernible inhibitory effect on the aggregation of tau (Fig. 3A, B). Curcumin inhibited the aggregation of tau in a concentration-dependent manner (Fig. 3C). The half-maximal aggregation inhibitory concentration (IC50) of curcumin was determined to be 10.6±1 μM (Fig. 3D). Using a light scattering assay, curcumin was also found to inhibit the aggregation of tau (Fig. 3E). The IC50 value was determined to be 9.2±1 μM. Further, curcumin also strongly reduced the rate of tau aggregation (Fig. 3F).

Curcumin inhibits tau aggregation. Tau (5 μM) was incubated without and with either 20 μM Methylene blue (MB), 20 μM curcumin or 0.4% (v/v) DMSO for 20 min. Then, the reaction mixtures were incubated with 75 μM AA at 37°C for 24 h. DMSO was taken as a solvent control. A) The aggregation was monitored by Thioflavin S assay at 521 nm (n = 3, * p < 0.01). B) The aggregation was monitored by 90° light scattering assay at 400 nm. Results are mean±SEM of 3 individual sets (* p < 0.01). C) The fluorescence spectra of Thioflavin S in the presence of tau and AA without (■) and with 1 (▿); 5 (◊); 8 (▴); 10 (□); 15 (★); 20 (⌂); and 30 (×) μM curcumin are shown. The spectra of tau without AA were shown as (●). D) The inhibitory effect (IC50) of curcumin on tau aggregation was determined using Thioflavin S. E). The inhibitory effect (IC50) of curcumin on the aggregation of tau was monitored by 90° light scattering at 400 nm. Results are mean±SEM. (n = 3, R2 = 0.97). F) The time course aggregation of tau (5 μM) in the presence of AA (75 μM) was monitored without and with 20 μM curcumin at 37°C. The aggregation was monitored by the Thioflavin S assay: tau in the presence of AA (▀); tau in the presence of AA and curcumin (●) (n = 3) (** p < 0.01).
The effect of curcumin on tau oligomerization was determined by dynamic light scattering (Fig. 4). Under the conditions used, the mean diameter of tau in the absence and presence of AA was determined to be 56±14 and 400±88 nm, respectively, suggesting that AA induced the formation of tau oligomers (Fig. 4A). However, the mean diameter of tau oligomers was estimated to be 61±15 nm in the presence of curcumin, indicating that curcumin inhibited AA-induced oligomerization of tau (Fig. 4A). The size distribution of tau oligomers/molecules is shown in Fig. 4B, C, and D. Further, AFM images also showed that tau formed oligomers upon incubation with AA for 24 h (Fig. 4E). Curcumin suppressed the oligomerization of tau (Fig. 4F). In the absence of AA, no oligomers were observed in the reaction mixtures containing either only tau (Fig. 4G) or tau with curcumin (Fig. 4H). In addition, an analysis of the AFM peaks of tau molecules in the presence of AA indicated that the height of 50% molecules was 16–30 nm. In contrast, 65% and 35% of tau molecules were found to have a height of 1–5 and 6–15 nm, respectively in the presence of curcumin (Fig. 4I). The findings, along with the DLS data, strongly suggested that curcumin inhibited the oligomerization of tau.

Curcumin inhibited tau oligomerization. Tau (5 μM) was incubated without and with 20 μM curcumin for 20 min. Then, the reaction mixtures were incubated without and with 75 μM AA for 24 h at 37°C. A-D) Tau oligomerization was analyzed by dynamic light scattering. A) The mean diameter of oligomers was determined using DLS. (* p < 0.05). B-D) The size distribution of tau molecules are shown in the absence of AA (B), in the presence of AA without curcumin (C) and with 20 μM curcumin (D). E-I) Atomic force microscopic images of 24 h tau oligomers under different conditions are shown; E) tau in the presence of AA, F) tau in the presence of AA and 20 μM curcumin, G) tau in the absence of AA, H) tau without AA in the presence of 20 μM curcumin. Scale bar = 2 μm. I) AFM histogram of tau + AA (■) and tau + AA + curcumin (●). The p-value for, bin height 1–5 nm: **7.4×10–6; 6–10 nm: non-significant; 11–15 nm: non-significant; **16–20 nm: 0.009; 21–25nm: **0.004; 26–30: *0.02, (n = 4).
Curcumin induces disaggregation of tau aggregates
Tau (5 μM) was incubated with AA for 24 h; the preformed tau aggregates were subsequently incubated with different concentrations of curcumin for 3 h. The effect of curcumin on the disaggregation of preformed tau aggregates was monitored by Thioflavin S fluorescence. Curcumin reduced the level of aggregated tau (Fig. 5A). For example, 20 μM curcumin caused 75±10% disaggregation of tau aggregates (Fig. 5B).
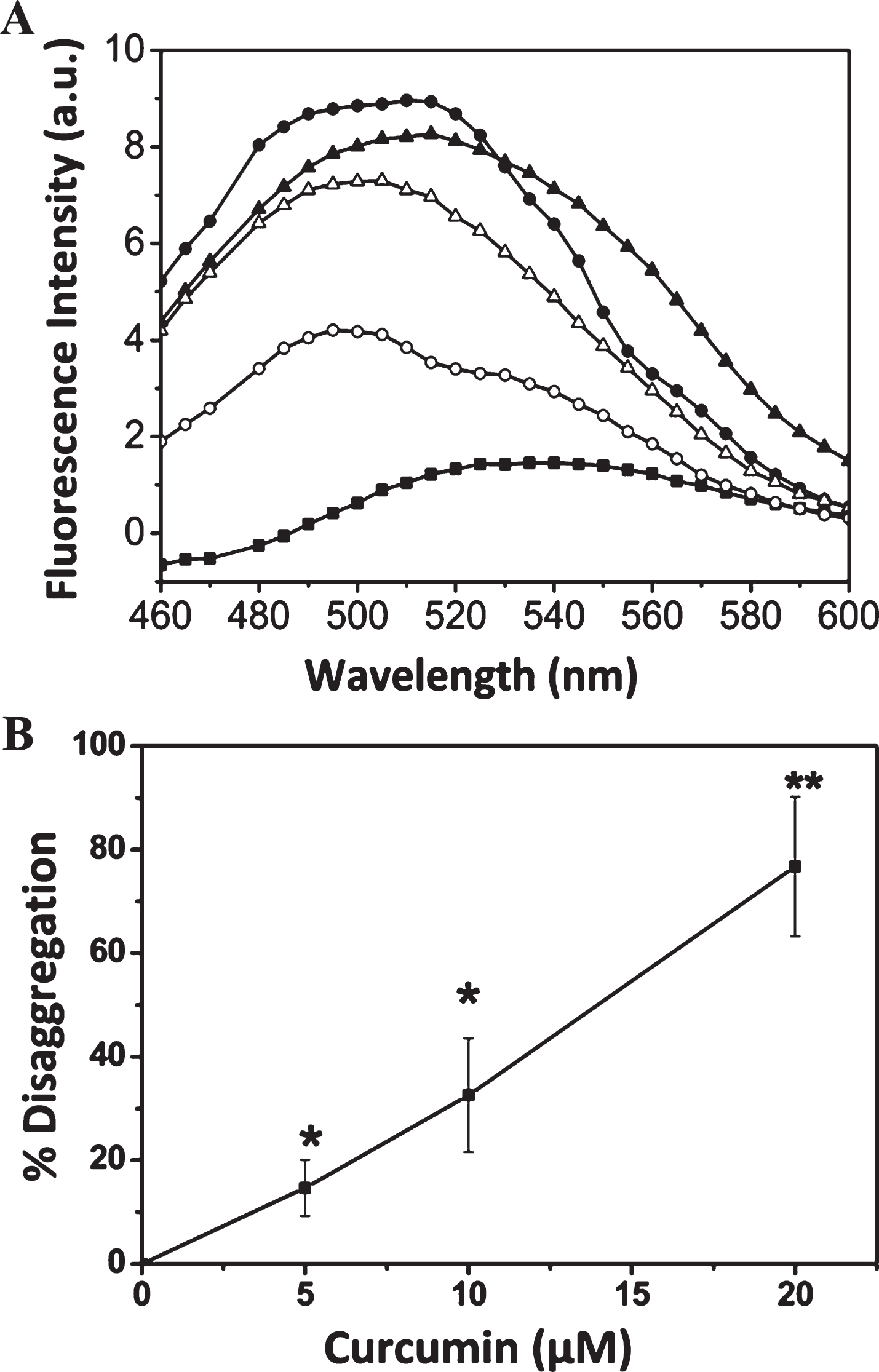
Curcumin induced the tau disaggregation. Tau (5 μM) was incubated with AA (75 μM) at 37°C for 24 h. Then, the preformed tau aggregates were incubated with and without curcumin at 37°C for 3 h. A) Thioflavin S spectra of tau aggregates without (●) and with 5 (▴); 10 (Δ) and 20 (○) μM curcumin. Thioflavin S spectra of soluble tau denoted as (■). B) The disaggregation of tau at different concentrations of curcumin is plotted. Results are mean±SEM (n = 3) (* p < 0.05 and ** p < 0.01).
Curcumin inhibits the formation of β sheet of tau
The effect of curcumin on the secondary structures of tau was analyzed by monitoring far-UV CDspectra. Consistent with the previous reports [44, 45], tau was found to primarily consist of random coil structures. The soluble tau exhibits a peak at 200 nm, which is an indication of random coil structures. The shift of tau peak towards higher wavelength and the broadening of the peak near 215 nm indicated the aggregation of tau (Fig. 6). An analysis of thesecondary structures indicated that there is an increase in β sheet content of tau in the presence of AA (Fig. 6). Curcumin reduced the β sheet formation in tau (Table 1). Therefore, curcumin may inhibit the aggregation of tau by inhibiting the β sheet formation in tau.
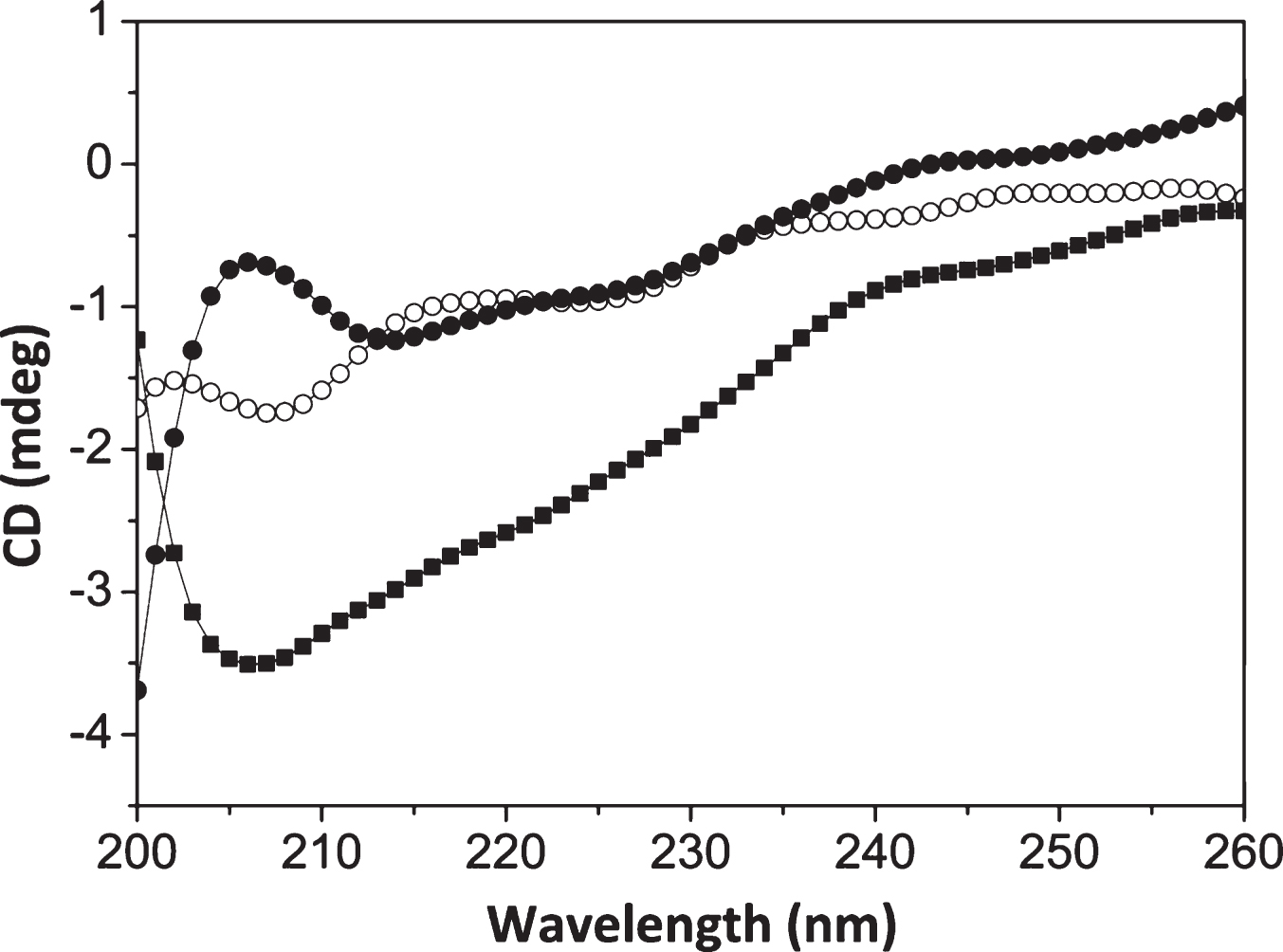
Curcumin inhibits β sheet formation of tau. Tau (10 μM) was incubated with AA (75 μM) in the absence and in the presence of 20 μM curcumin at 37°C for 24 h. The structural change was monitored by CD. The spectra of only tau (○); tau with AA (■); tau with AA and curcumin (●) are shown. (n = 5).
Secondary structures of tau were estimated from the far-UV CD spectra
Results were mean±SEM. (n = 5). The p-value was calculated by comparing the data of tau in the presence of AA and tau in the presence of AA and curcumin.
Curcumin inhibits the filamentation of tau
The effect of curcumin on tau filamentation was examined using electron microscopy and atomic force microscopy (Fig. 7). Tau formed filaments and small aggregates in the absence of curcumin while filaments were not observed in the presence of curcumin (Fig. 7A, B). Here the observed filaments were not the same as the pathological forms found in Alzheimer’s disease brain. The tau filaments were artificially prepared using AA. In the absence of AA, tau did not form filaments (Fig. 7C). Also, no tau filaments were observed in the presence of curcumin without AA (Fig. 7D). The electron microscopy results were corroborated by atomic force microscopic analysis (Fig. 7E, F). In the absence of curcumin, tau fibrilization took place along with the formation of tau aggregates (Fig. 7E). The mean height of tau filaments was found to be 3.2±0.1 nm (Fig. 7E). Curcumin strongly inhibited tau fibrilization (Fig. 7F). Atomic force microscopic images showed that tau did not form filaments in the absence of AA (Fig. 7G). Also, no tau filament was observed in the presence of curcumin without AA (Fig. 7H). The results together suggested that curcumin inhibited tau filamentation, which is considered to be an important step in the PHFs formation in the Alzheimer’s disease.

Curcumin inhibits the filamentation of tau. Tau (20 μM) was incubated with AA (75 μM) in the absence and in the presence of 20 μM curcumin at 37°C for 16 days. A-D) Electron microscopy: tau in presence of AA (A); tau in presence of AA and curcumin (B); only tau (C); tau in presence of curcumin (D); scale bar = 200 nm. E-F) Atomic force microscopy: tau in presence of AA (E); tau in presence of AA and curcumin (F); only tau (G); tau in presence of curcumin (H), scale bar = 1 μm.
Curcumin disaggregates tau fibrils
Tau was incubated with AA for 16 days to form filaments. The preformed tau filaments were incubated without or with curcumin for 24 h. The effect of curcumin on the preformed tau filaments were examined by electron microscopy, atomic force microscopy, and light scattering (Fig. 8). The electron microscopic images showed that curcumin disintegrated the tau filaments (Fig. 8A-C). The AFM images demonstrated that the most of the preformed tau filaments were disassembled upon curcumin treatment (Fig. 8D-F). Further, a light scattering experiment indicated that curcumin could disassemble tau filaments (Fig. 8G).
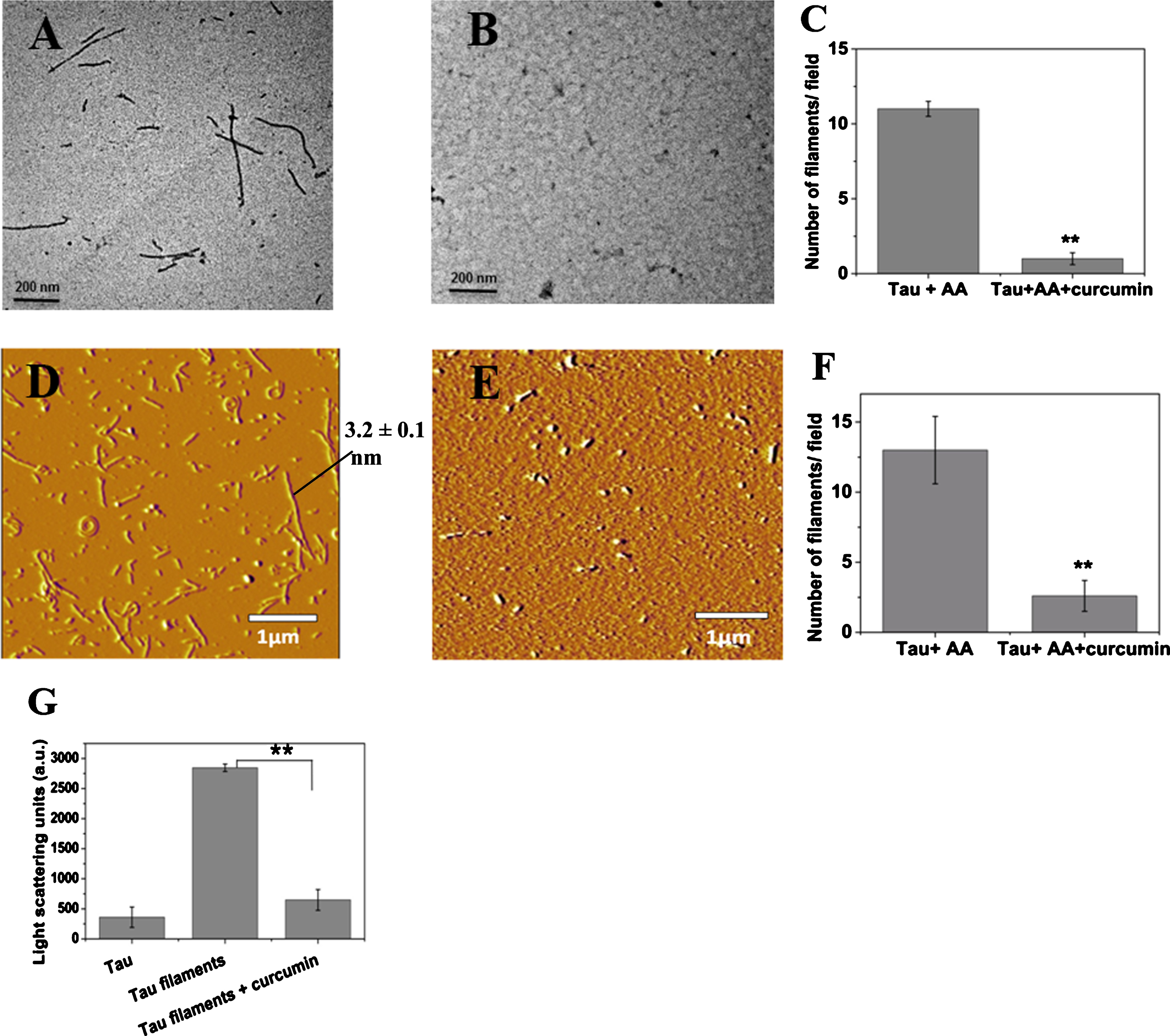
Curcumin disaggregates preformed tau filaments. Tau (20 μM) was incubated with AA (75 μM) at 37°C for 16 days. The preformed tau filaments were incubated in the absence and in the presence of 20 μM of curcumin at 37°C for 24 h. A-C) Electron microscopy: tau aggregates (A); tau aggregate in the presence of curcumin (B); scale bar = 200 nm. Curcumin reduced the number of tau filaments per field of view (C). D-F) Atomic force microscopy: tau aggregates (D); tau aggregate in the presence of curcumin (E); scale bar = 1 μm. Curcumin reduced the number of tau filaments per field (F). Results are mean±SEM of 10 individual images (** p < 0.01). G) The preformed tau filaments were incubated with 20 μM curcumin for 24 h and the defibrillation was monitored by 90° Light scattering (** p < 0.01) at 400 nm.
DISCUSSION
The formation of neurofibrillary tangles due to pathological aggregation of tau is a hallmark of tauopathies [46, 47]. The inhibition of tau aggregation is thought to be a good therapeutic strategy for treating tauopathies [48 –50]. Small molecules inhibitors have been screened out, including the identification of compounds from a combinatorial library screen [18 , 51–54]. In this study, we found that a natural product curcumin potently inhibited the aggregation of purified tau, a key process in the Alzheimer’s disease. Further, it increased the disaggregation rate of tau indicating a possibility that even after the formation of the neurofibrillary tangles; curcumin could reverse it by dissolving the polymers.
Curcumin binds to 4R0N tau with much stronger affinity than 3R0N tau, lacking R2 part of MTBR domain, indicating that R2 is involved in the binding of curcumin. A docking analysis reveals that curcumin forms strong hydrophilic interaction with D225 (R2 MTBR) residue of tau. Further, mutations (V255A, V292A) were incorporated in MTBR region of tau using computational approach. The docking of curcumin on the in silico mutated tau constructs indicated that curcumin weakly binds the mutated tau constructs as compared to the native tau (Supplementary Table 1). The electrostatic interaction between curcumin and MTBR region might hinder the tau-tau interaction, which was hypothesized to be the most important step in the aggregation process of tau. Therefore, the biochemical binding data and docking analysis together indicated that curcumin binds to the MTBR regions of tau, which plays an important role in the aggregation of tau [41 , 56].
Curcumin inhibited tau aggregation in a dose-dependent manner. The IC50 value of curcumin (10.6±1 μM) was in the range of some potent tau aggregation inhibitors like hemin chloride (8.2 μM) and thionin (12 μM) [54]. However, curcumin was found to display weaker activity than several inhibitors of tau aggregation, including methylene blue (IC50, 1.9 μM), azure A (IC50, 2.6 μM), azure B (IC50, 1.9 μM), N744 (IC50, 280 nM), myricetin (IC50, 1.2 μM), and exifone (IC50, 3.3 μM) [54, 57]. Importantly, curcumin could suppress the rate of tau aggregation. The aggregation of tau is a multistep process; the initial step of tau aggregation is the formation of β sheet of tau where the MTBR region of tau stacked upon each other [45]. Further, it forms dimer and trimer, and then small soluble oligomers [58]. These small soluble oligomers forms twisted tau filaments called PHFs and subsequently, PHFs form neurofibrillary tangles [58]. The aggregation of tau was found to be associated with the formation of β sheets, and curcumin inhibited theβ sheet formation in tau indicating that curcumin suppresses the tau aggregation at its initial stage. The DLS and AFM experiments revealed that curcumin inhibited tau oligomerization (Fig. 4). However, we could not determine the effect of curcumin on tauoligomerization using SDS-PAGE [59, 60]. Under the conditions used, the mobility of tau oligomers in the SDS-PAGE was not observed possibly due to the formation of high molecular weights tau oligomers. Further, the disaggregation study in vitro suggested that curcumin treatment can dissolve preformed tau aggregates and filaments, indicating that even after the formation of the tangles, the tau-mediated symptoms can be corrected.
In summary, curcumin bound to tau, potently inhibited the aggregation of tau and dissolved the tau aggregates in vitro. We also provide a possible mechanism of the inhibition of tau aggregation by curcumin. In addition to its ability to inhibit tau aggregation, curcumin has several other neuroprotective benefits because of its anti-inflammatory and antioxidant activities [20, 21]. It also inhibits the aggregation of amyloid proteins like Aβ and α-synuclein [22, 23]. Therefore, it might have potential as a lead compound for the development of small molecule inhibitors that reduce amyloidogenesis in neurodegenerativediseases.
Footnotes
ACKNOWLEDGMENTS
The work is supported by a grant from Department of Biotechnology, Government of India. DP and JSR are supported by TATA Innovation fellowship from Department of Biotechnology, and INSPIRE fellowship from Department Science & Technology Government of India, respectively. We sincerely acknowledge Prof. S. Feinstein, University of California Santa Barbara for the tau clone. We are also thankful to central instrumentation facility, Indian Institute of Technology Bombay for atomic force microscopy and electron microscopy.