
Abstract
Peripheral biomarkers for dementia are few and far between. Despite research into blood plasma/serum biomarkers for dementia diagnostics, there is a lack of information on erythrocytes and their vast proteomes as potential biomarkers. This review identifies a number of relevant and potentially promising erythrocyte biomarkers for various subtypes of dementia. These include erythrocyte morphology, oxidative stress, and erythrocyte membrane proteins such as the glucose transporter (GLUT-1), amyloid-β, IgG, Hsp90, calpain-1, and band 3 protein. Of those proteins identified Hsp90, amyloid-β, calpain-1 and band 3 show the most promise as pre-clinical biomarkers. However, the most intriguing aspect of erythrocytes is their changed morphology in dementia. The altered morphology not only could be used as a diagnostic biomarker but may be crucial in early pathogenesis of the disease. Further work must be done to establish the pathological connection between the periphery and central disease processes.
INTRODUCTION
The search for reliable blood biomarkers to diagnose dementia has largely concentrated on the plasma or serum. A number of promising peripheral biomarkers have been identified in plasma and serum, such as the inflammatory (IL6 and IL10) and trophic markers (proGNF and TNF-alpha), matrix metalloproteinases (MMP3 and MMP9) [1], amyloid-β peptides [2], α-1-antitrypsin, α-2-macroglobulin, apolipoprotein E,and complement C3 [3], but none of them have been taken forward for use in routine clinical settings. This is primarily due to the undefined origin of these markers and their inconsistent reproducibility, the unreducible relationship with cognitive and non-cognitive dementia symptoms, as well as their unknown relationship to other age-associated diseases and associated pharmacological treatments. In contrast, identifying dementia biomarkers in blood cells somewhat lags behind, largely due to the laborious methodology to isolate certain blood cells and concentrating cellular proteins to measurable levels. The latter have been largely seen as potentially unpractical for development of user-friendly portable devices.
Among blood cells, platelets have been extensively investigated [4 –6], and a number of characteristic Alzheimer’s disease (AD) related proteins have been found to correlate with both cognitive [4, 7]and non-cognitive, behavioral [8] changes in AD. However, more work is now needed to determine whether the platelet proteins may successfully differentiate between distinct dementia subtypes and how these changes respond to current anti-dementia therapies. The potential of other peripheral blood cellular components has not yet been fully investigated. Thus, the type and number of circulating peripheral blood mononuclear cells, including lymphocytes, macrophages, monocytes and leukocytes, in particular, may reflect both the age- and dementia-associated upregulation of certain types of immune/inflammatory responses [9] and altered calcium signal transduction pathway [10]. Although some of these preliminary findings seem promising, further evaluation needs to follow.
Since all blood cells are generated from the same multipotential hematopoietic stem cell pool (hematocytoblasts), and erythrocytes and platelets derive from common myeloid progenitor cells, red blood cells are potential candidates for dementia biomarkers and may be as important as other blood cells that have been investigated so far. Furthermore, erythrocytes are the largest population of blood cells, accounting for 40–45% of all sanguine cellular components, and thus provide viable material to be investigated further for dementia on par with the plasma. In this review, we address the evidence for erythrocytes as biomarkers for dementia, and discuss their utility in routine clinical setting. In particular, we focus on the morphology changes in erythrocytes and discuss several dementia-related proteins identified in erythrocytes, i.e., GLUT-1, amyloid-β (Aβ), IgG, Hsp90, calpain-1, and band 3 protein.
SEARCH METHODS
This review is based on a comprehensive search that included Medline, Embase, GoogleScholar, and PubMed databases and studies published until 31 January 2017. The key words used in the current search were: dementia, AD, vascular dementia, Parkinson’s disease, dementia with Lewy body, erythrocyte, biomarker, protein, enzyme, proteomics, and genomics. The identified erythrocytic protein/enzyme components were further utilized to search for additional references in relation to dementia, and they included: morphology, inflammation, oxidative stress, amyloid-β protein precursor, iron, ferritin, GLUT-1, Aβ, IgG, Hsp90, calpain-1, and band 3 protein. Alpha-synuclein is not included in the current review in the light of recent publications concentrating on its function and role in erythrocytes [11]. All abstracts were critically reviewed by at least two co-authors (AS and EBM-L, or DL and EBM-L, DL, PK and EBM-L, or PK and EBM-L) and the full text of relevant papers was obtained, and critically evaluated in terms of their clinical relevance.
ERYTHROCYTES IN AGING AND DEMENTIA
Erythrocytes in healthy aging
In healthy aging, erythrocytes naturally live up to 120 days in circulation. The aging of the erythrocytes constitutes in increased cell density and reduced cell volume. This structural alteration has been linked to oxidative damage. The removal of senescent erythrocytes is mediated by conformational changes in band 3 which then allows the binding of autologous IgG [12]. Both erythrocyte count and hemoglobin (Hb) levels decrease in the elderly [13]. This suggests that oxygen delivery is reduced with age. Potentially, these factors contribute to the pathology of age-related neurodegeneration.
Erythrocytes preparation for biomarker analysis
As most erythrocyte biomarker studies to date are purely exploratory, there is little consensus on any given marker. Therefore, it is important that the methods used are robust and comparable. The aim is to isolate non-nucleated biconcave red cells in the absence of plasma and platetlets. Current studies have used various methods to extract and purify these cells (see Tables 1 and 2). These differences are due to the particular biomarker focus of the studies, specifically in relation to whether the proteins are intracellular, extracellular, or within the membrane. Studies exploring associated and bound proteins simply centrifuge and wash the erythrocytes in PBS in order to remove nucleated cells or cell fragments such as platelets [14, 15].
Erythrocyte proteome in dementia. Summary of the literature showing alterations of the erythrocytic proteome in neurodegenerative disease
3MSE, Modified Mini-Mental State Examination (scale 1 to 100); AChE, acetylcholinesterase; AD, Alzheimer’s disease; ADAS, Alzheimer’s Disease Assessment Scale; AMC, age-matched controls; BuChE, butyrylcholinesterase; C, controls; CAT, catalase activity; CDR, Clinical Dementia Rating; DSM III, Diagnostic and Statistical Manual of Mental Disorders III; ERC, erythrocyte; GDS, Global Deterioration Scale; GSH, reduced glutathione; GSSG, oxidized glutathione; Hb, hemoglobin; MCI, mild cognitive impairment; MIX, mixed dementia; MMSE, Mini-Mental State Examination; MRI, magnetic resonance imaging; NOx, sum of nitrates and nitrites as measured via the main metabolic end-products of nitric oxide and peroxinitrite anion; NPB, neuropsychological battery; NINCDS-Alzheimer’s ADRDA, National Institute of Neurological and Communicative Disorders and Stroke and the Alzheimer’s Disease and Related Disorders Association; PC, protein carbonyl; PDD, Parkinson’s disease dementia; PIT, peroxidation induction test; RBC, red blood cells; VaD, vascular dementia; YC, young controls.
Erythrocyte proteome in comorbidities of dementia. Summary of literature showing alterations of the erythroctic proteome in diseases which are known dementia risk factors including aging, obesity, diabetes mellitus, and Down syndrome
AD, Alzheimer’s disease; Aβ, amyloid-β; BMI, body mass index; C, controls; DS, Down syndrome; ERC, erythrocyte; GSH, reduced glutathione; NINCDS-Alzheimer’s ADRDA, National Institute of Neurological and Communicative Disorders and Stroke and the Alzheimer’s Disease and Related Disorders Association.
Some have studied erythrocyte ghost membranes; these methods significantly reduce any interference from the large amounts of Hb found in erythrocytes [16]. This is a well-established method that has been recently confirmed to be comparable to biologically intact erythrocyte membranes [17].
Others have lysed the cells to study the isolated intracellular and membrane proteins in neutral pH buffer such as pH 7.4 Tris [15] or phosphate buffer [18]. Hemolysates have potentially the mostbeneficial use in a clinical setting if a multiplex assay was developed to analyze multiple protein levels in erythrocytes.
Erythrocyte morphology and Alzheimer’s disease
Proteomic studies have identified 751 proteins in the human erythrocyte [19]. Of these, 340 are membrane proteins and suspected to have a role in erythrocyte membrane stability and morphology [20]. Recent studies on erythrocyte cell membrane proteins reported decreased deformability in dementia subjects and normal controls’ erythrocyte morphology [21]. It has been reported that 15% of erythrocytes in AD subjects are elongated and have altered membrane architecture [18].
In AD subjects, the erythrocyte morphological changes appear closely linked to higher erythrocyte levels of un-liganded iron and ferritin [22], as well as direct [23] or indirect (via fibrin [24] or copper [25]) erythrocyte interactions with plasma Aβ protein. Also, when erythrocytes are exposed to Aβ1 - 40 or Aβ1 - 42 fibrils, they show 8.6% or 11.1% elongated cells, respectively, in contrast to only 2.9% or 1.3% of erythrocytes being elongated when blood is treated with buffer or mock fibrils generated from Aβ42 - 1 [21]. Furthermore, 98% of peripheral AD erythrocytes are binding with amyloid, in comparison to only 38% in healthy control older people [23]. The interaction between the Aβ peptide and erythrocytes also results in larger cell size [26], increased mean corpuscular volume [27, 28], and more low density erythrocytes that contain elevated levels of Aβx - 40 peptide [28].
Interestingly, some of the anti-dementia drugs, i.e., tacrine, appear to strengthen cytoskeletal protein-protein interactions within the erythrocyte membranes [29]. This suggests that the currently available anti-dementia drugs, except for their properties of increasing acetylcholine in the AD brain tissue, may also improve the erythrocytes elasticity, flexibility, and shape by correcting the erythrocyte membrane deformability, and thus help to improve blood flow and microcirculation [30, 31].
The altered erythrocytes morphology in dementia has a direct impact on increased erythrocyte aggregation and sedimentation. Flow abnormalities, frequently seen in blood vessels of people with dementia, are also present in bloods of people with other degenerative diseases (i.e., atherosclerosis, diabetes, and inflammation), all of which are known to be associated with AD and vascular dementia[32, 33].
Erythrocyte membrane components
Several membrane and cytoskeletal proteins are linked to alterations of erythrocyte morphology in AD subjects [21 , 34]. Normal erythrocyte deformability depends on the integrity of certain membrane proteins. The cytoskeleton is predominantly composed of spectrin tetramers (α and β subunits) associated with other proteins, such as erythrocyte membrane protein band 4.1, tropomyosin, glycoproteins, and actin filaments at the junctional complex site [18]. Other peripheral proteins, such as demantin and adducin, also participate in the junctional complex and play a role in maintaining erythrocyte shape and membrane stability [35] (Fig. 1). Alterations of these proteins could have debilitating effects on erythrocyte flexibility and stability, hence, may contribute to the decreased deformability and increased elongation of the erythrocytes as found in AD subjects. Interestingly, the levels of demantin, tropomodulin, tropomyosin, glycophorin, and F-actin capping proteins were increased in AD subjects compared to control counterparts [18]. In addition, the complete elimination and/or reduction of some of these proteins, such as demantin and β-adducin, caused erythrocyte fragility and anemia [35]. These findings therefore suggest that erythrocytes in AD appear to be less elastic and more prone to damage as a result of specific erythrocyte membrane protein changes. It is both difficult and compelling to speculate on the nature or these changes, and why they occur. Are they the initial site of a dementia disease process, or a consequence of a dementia process seen as a systematic illness, that does not affect only the brain tissue, but also has an impact to vital organs at the periphery? Tables 1 and 2 summarize the changes of erythrocyte membrane proteins discussed in the literature.
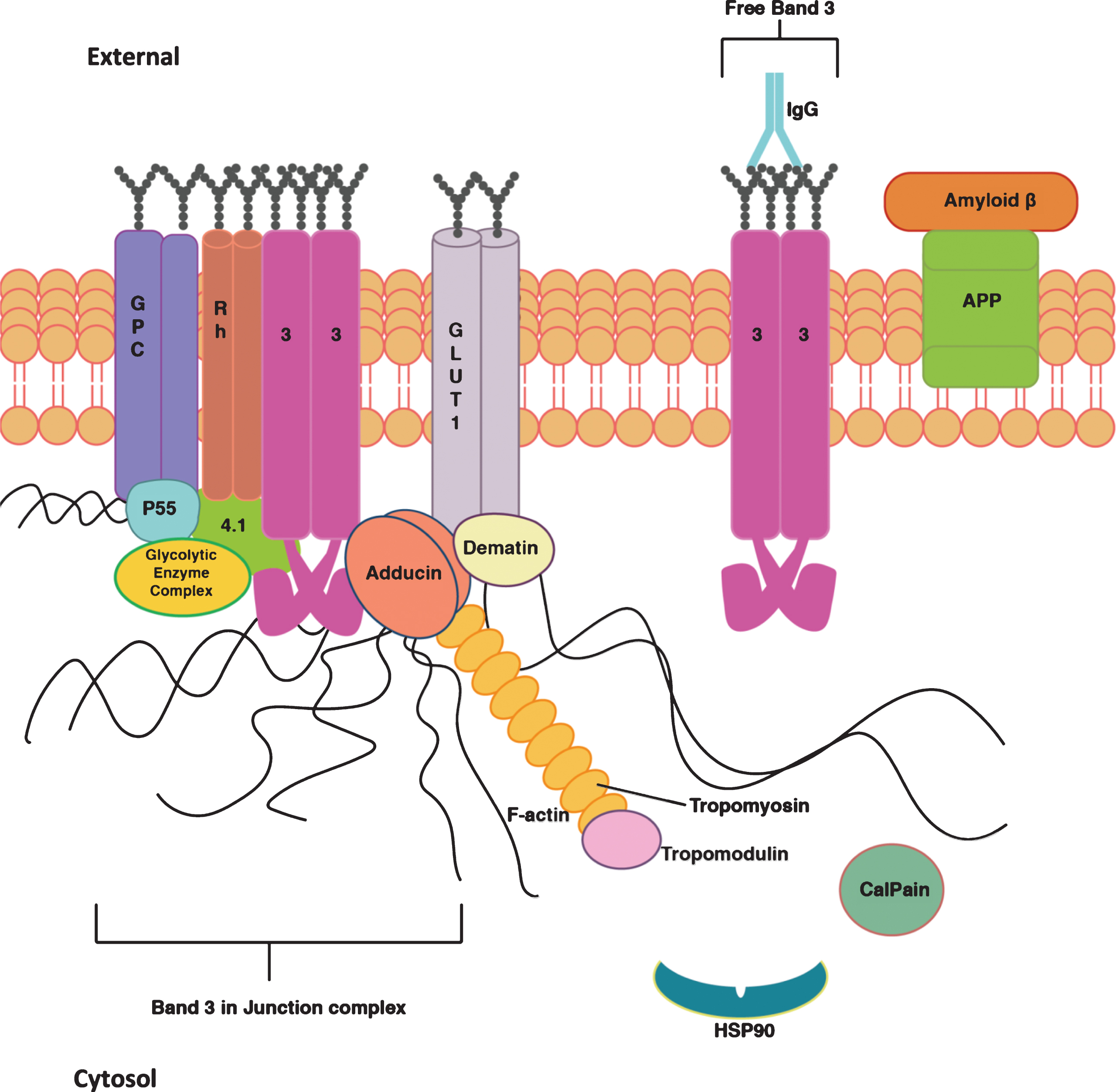
Erythrocyte membrane proteins. The interactions and functions of the proteins in the erythrocyte (ERC) membrane. The figure shows the junctional complex made up of glycoporin C (GPC), Rh protein (Rh), band 3 anion exchanger (3), Glucose transporter 1 (GLUT-1), demantin, adducing, P55, protein 4.1, glycolytic enzyme complex, F-actin, tropomyosin and tropomodulin. This complex anchors spectrin in the cytoskeleton. band 3 anion exchanger also exists separately in the membrane. There are chaperone and enzyme proteins present in the cytosol such as calpain and HSP90. IgG and amyloid beta are peripheral proteins which associate with the surface of the membrane. Amyloid-β is associating with amyloid-β protein precursor (APP), it is unclear whether it may also bind directly to ERC surface membrane.
There are several methods employed which define erythrocytes as appropriate biomarkers for dementia. Some studies extract the red blood cells and analyze them as complete cells analyzing any associated or bound proteins, others lyse and measure levels of intracellular proteins and then there are studies which extract the erythrocyte membrane ghosts and analyze these.
RELEVANT MOLECULAR ERYTHROCYTE CHANGES IN DEMENTIA
Alterations of erythrocytes during oxidative stress
Oxidative stress is a predominantly vascular event, and erythrocytes would, therefore be prone to similar alterations. Erythrocytes are highly susceptible to oxidative damage due to the presence of Hb, which transports molecules of oxygen. As a by-product of Fe(II) to Fe(III) functional oxidation, oxygen-free radicals are produced, causing structural and morphological changes in the erythrocyte membrane and cytoplasmic proteins and lipids.
Consequences of oxidative stress are remarkable in AD postmortem brains particularly in areas of increased degradation [36]. However, these changes do not appear to be characteristic for the preclinical or mild stages of dementia. Thus, measures of oxidative stress markers in erythrocytes, i.e., antioxidant enzymes (superoxide dismutase (SOD), catalase, glutathione peroxidase, and total glutathione level) are similar between cognitively intact subjects and those with subclinical or mild AD [37] (Fig. 2). However, in more severe cases of AD, the oxidative parameters are intensified. In particular, erythrocyte expression of TBARS, SOD, and Na, K-ATPase are increased in comparison to cognitively age-matched control subjects, as well as young control subjects [38]. This suggests that the oxidative stress measures in dementia appear to be associated with clinically overt dementia stages. Thus, oxidative stress may not be the initial precursor of the dementia pathology, but rather an outcome of either disease progression or a by-product of a yet unidentified pathological dementia mechanism.

The effects of aging on the interactions between oxidative stress and erythrocyte membrane proteins. Pathways which elucidate the effects of aging on oxidative stress and erythrocytes’ proteomic alterations. The blue arrows illustrate the consensus on the general changes in levels of these proteins identified in erythrocyte membranes when affected by aging. Age increases levels of oxidative stress which in turn effect GSH, SOD, GPx, GSSG, and PC in erythrocytes. Age also induces conformational changes in band 3 which activate Calpain1 and lead to the expression of SCA on the outside of the membrane. IgG levels increase as they bind to SCA. General levels of Hb fall as people age. Aβ binds to the outside of the erythrocytes as levels increase in circulation when aging. GSH, reduced glutathione; SOD, superoxide dismutase; GPx, glutathione peroxidase; GSSG, oxidized glutathione; PC, protein carbonyl; SCA, senescent cell antigen; Hb, hemoglobin.
High oxidative stress in erythrocytes induces the production of reactive oxygen species (ROS). ROS highly affects the structure of erythrocytes as they have the ability to alter Hb and oxygen structure to produce methemoglobin. This decreases the affinity of oxygen binding, hence, decreases oxygen transportation to the brain [39]. Also, oxidation of Hb can lead to an altered Hb structure resulting in the production of Heinz bodies. These Heinz bodies are removed by reticulo-endothelial macrophages, producing further oxidative stress which eventually results in erythrocyte hemolysis [40]. Furthermore, ROS can affect cytoskeleton membrane composition by oxidizing amino acid residues, resulting in further protein fragmentation and degradation. This, therefore, leads to further impaired deformability of erythrocytes and markedly contributes to erythrocyte elongation [41, 42] and consequent cellular death. There are many parallels that could be drawn between membrane proteins that are affected by oxidative stress in erythrocytes and similar changes linked to CNS pathology in AD. Further insight into this interplay between oxidative stress in the brain and the periphery would clarify this. The proteins involved in oxidative stress include: GLUT-1, Aβ, IgG and Hsp90.
GLUT-1
Closely related to oxidative stress is altered glucose metabolism. In AD, there is a slowing of glucose metabolism causing glucose starvation possibly secondary to hypoxia. The GLUT-1 transporter, which primarily transports glucose and docosahexaenoic acid, is highly abundant in erythrocyte membranes. In addition to erythrocytes, it is highly expressed in the blood-brain barrier epithelium and astrocytes.
In contrast to brain studies, a significant increase in GLUT-1 transporters and insulin receptors on erythrocytes in AD subjects compared to healthy controls has been described [43]. This is similar to a previous study that found an increase in hyperglycemic bloods associated with diabetes [44], a condition that increases the dementia risk, including AD and vascular dementia. As these changes are characteristic for diabetic patients, this could confound GLUT-1 as a biomarker that is specific to dementia patients. However, to date there are no specific studies addressing GLUT-1 changes either in the cerebrospinal fluid or bloods/blood compartments to support the latter.
Aβ and ubiquitin interactions
The presence of Aβ plaques in the postmortem AD brain tissue is the defining pathology of AD. It is also among the two confirmed cerebrospinal fluid biomarkers used to diagnose AD [45]. Aβ impairs mitochondrial function, synaptic plasticity, and long-term potentiation, and ultimately causes apoptosis in neuronal cells [23]. Since erythrocytes lack both mitochondria and nuclei, Aβ causes erythrocyte damage indirectly, via erythrocyte membrane binding and oxidative stress [46]. In particular, it is the hyperphosphorylated Aβ that binds to the erythrocyte membrane proteins [15] and causes phospholipid peroxidation and decrease in erythrocyte endogenous carotenoids, important for erythrocyte oxidative function [15, 47]. In addition, adherence of Aβ to erythrocyte’s integral proteins causes oxidative impairments in erythrocyte function by decreasing its deformability, leading to decreased blood flow to the brain, and thus contributing to brain hypoxia leading to further neuronal loss in AD [21, 26].
Other studies suggest that the modification of erythrocyte’s shape and morphology is affected by Aβ-induced oxidative damage [48]. Oxidative damage in erythrocytes affects the cytoplasmic membrane resulting in lipid peroxidation. When exposed to by-products of lipid peroxidation, such as 4-hydroxynonenal, Aβ can act as an inhibitor of ubiquitin-dependent protein degradation in vitro by affecting the proteolytic core, 20S proteasome, through the inhibition of its chymotrypsin-like activity [49]. This interaction may explain the pathological manifestation of the increased Aβ levels that are not found only in the brain of subjects with AD, but also their erythrocytes [50].
Failure to activate ubiquitination of spectrin sub-units, alongside the increase in oxidative damage of actin in the cytoskeleton can modify the erythrocyte cytoskeletal morphology [20]. This is evident in sickle cell disease patients, an irreversible disease causing the aggregation of erythrocytes due to its modification to sickle shape, making erythrocytes increasingly more adhesive. In turn erythrocyte ubiquitin levels may influence their morphology and shape.
Band 3 protein
The erythrocyte membrane protein, band 3, is considered to be involved in a number of functions such as the maintenance of cell stability [51]. Erythrocytes undergo cellular removal, as a result of healthy aging, through the binding of serum IgG to senescent cell antigen (SCA) produced from band 3 anion exchanger [52, 53]. Band 3 proteins in erythrocytes have electrochemical and physical roles as they act as a bicarbonate/chloride exchanger [54] and membrane stabilizer [55, 56], respectively. In terms of dementia, the latter role seems significant as erythrocytesdisplay various morphological membranous alterations [17], which could be attributed to the loss of the membrane stabilizer, band 3. Additionally, the increased degradation of band 3 in AD erythrocytes would increase its by-product SCA which binds to IgG [57]. This in turn possibly increases the membrane-bound IgG in AD erythrocytes. As IgG initiates cellular removal, its presence could signify cellular aging. Based on this, AD may be associated with rapid cellular erythrocyte aging. This needs to be explored in further studies.
There is an accelerated breakdown of band 3 protein in AD subjects, as demonstrated by measuring the anion exchange rate and levels of glucose transport into blood cells [14]. Thus, increase in IgG levels correlate with the decrease of band 3 proteins in AD subjects [14]. This was backed up and explored further through a novel technique measuring anion exchange [58] that also confirmed decreased levels of faulty band 3 proteins, as a result of enhanced degradation, in AD patients.
Immunoglobulin G
During physiological aging, erythrocytes invariably accumulate products of oxidative damage. As a result, erythrocytes produce iron chelators that promote increased binding of autologous IgG [59]. Although IgG binds to erythrocyte membranes, they also appear to have an affinity toward the erythrocyte cytoskeletal proteins, with the cytoskeleton being a primary target in the opsonization of hemolyzed erythrocyte membranes by naturally occurring plasma autoantibodies [60]. However, the plasma immunoglobulin opsonization of the cytoskeleton may occur well prior to the decomposition of hemolyzed erythrocytes [60], suggesting that cytoskeletal opsonization represents the initial step in the removal of hemolyzed erythrocytes from the circulation. Aged erythrocytes are opsonized using IgG by recognizing SCA [61].
There is an increase in IgG levels on erythrocyte surface membranes in subjects with neurodegenerative-related diseases such as Parkinson’s disease and AD [62]. This, alongside the findings that an increase in erythrocyte-bound IgG enhances the degradation of band 3 protein in patients with Down syndrome [63], suggests that this process may indicate abnormal aging, and thus even be associated with AD pathology. However, in a subsequent study they found that increased IgG levels were not linked directly to AD progression [64]. There arecorrelative clinico-neuropathological studies that have found a strong correlation between Aβ-IgG levels with the Clinical Dementia Rating scale commonly used to assess cognitive function in AD. This argues that blood-based IgG measure may be used as a by-proxy measure of cerebral Aβ and has a potential to serve as a diagnostic tool to assess the progression of AD [65].
Calpain-1
It has been previously reported that continued activation of calpain-1 results in increased AD pathology [66]. Calpain-1, a Ca2+-activated proteolytic enzyme which is inhibited by Calpastatin, may also play a role in modifying cytoskeletal membrane proteins in AD erythrocytes. Thus, Caplain-1 mediates proteolysis of important membrane proteins such as band 3, adducin, demantin, and protein 4.1R in AD erythrocytes [67]. As reviewed above, degraded cytoskeletal proteins contribute to erythrocytes deformability in AD patients [66, 68]. Further investigations with calpain-1-knockout mice showed a decrease in Ca2 + activity, resulting in an increase of erythrocytes deformability [67, 68] which indicates that calpain-1 negatively regulates erythrocyte membrane stability and morphology. Interestingly, one of the calpain-1 inhibitors, BDA-410, appears to have a direct influence on erythrocyte morphology and stability via preventing hypoxia-induced erythrocyte dehydration [69]. It is thus not surprising that this inhibitor has been taken into clinical trials, demonstrating further its clinical potential in terms of both restoring normal synaptic function and improving spatial-working memory in an APP/PS1 mouse animal model of AD [70]. Moreover, since calpain-1 mediates degradation of Hb, we cannot exclude the possibility that calpain-1 altered metabolism may also lead to the formation of Heinz bodies [71]. However, their role in dementia is not yet fully understood.
Heat shock protein 90 kDa
One of the erythrocyte proteins identified by proteomic analysis that is closely associated with dementia is Hsp90. It is a cytoplasmic protein that has a number of physiological roles, mainly to respond to oxidative stress accumulated through aging and the progression of disease. It has a diverse function in cellular-signaling pathways and cellular development, currently extensively researched in preventing cancer proliferation [72].
Hsp90 is part of a family of chaperone proteins, which promote the clearance of misfolded proteins and the prevention of aggregate formation via promotion of proteasomal and autophagic lysosomal pathways [73]. There is growing evidence that Hsp90 inhibitors could be a viable treatment for AD [74], and thus having a peripheral blood biomarker that will address the expression of Hsp90 at the periphery will be clinically relevant. A number of studies have repeatedly confirmed an increase in erythrocytes Hsp90 levels in AD subjects compared to controls [18 , 76]. These findings suggest that erythrocyte Hsp90 levels have a potential to be used as a diagnostic biomarker in AD. However, further studies are needed to fully understand the clinical relevance of this potential dementia biomarker, and what role it plays in the pathophysiological processes involved in dementia.
SUMMARY AND CONCLUSIONS
The clinical symptomatology of dementia is closely related to the extent and topographical location of the pathological hallmarks including plaques, tangles, synaptic loss, vascular changes etc. Although advances have been made to visualize the central pathological features using functional neuroradiological tools, having a peripheral biomarker, closely related and reflecting the central dementia pathological process still remains a goal for the future. Erythrocytes appear to reflect the intrinsic pathophysiological characteristics of a number of dementia syndrome, including AD and vascular dementia. Most of the altered erythrocyte proteome is related to cellular deformability, aging and oxidative stress, elements which are all closely interlinked to aging and neurodegenerative disorders (Tables 1 and 2). However, oxidative stress has been demonstrated to be a consequence of AD pathology and altered protein levels rather than a cause of AD. This questions the usefulness as an erythrocyte preclinical biomarker for dementia. A far more promising biomarker of dementia is the erythrocyte morphology and, specifically, the decreased elasticity which makes them more prone to damage. Altered erythrocyte morphology in AD is the common result of almost all altered erythrocyte membrane proteins discussed in this review. So far, the most promising of these erythrocyte protein biomarkers are Hsp90, band 3, and IgG. The difficulty with the other explored proteins is their lack of specificity to dementia pathology and non-consistent findings. The issue of common erythrocyte protein changes between progression of neurodegenerative diseases and the effects of drugs on erythrocyte protein levels must be carefully considered and explored further prior to accepting erythrocyte protein biochemistry in the dementia biomarker field. The data largely come from studies based on small clinical samples, and predominantly including subjects with AD. Especially, there is an absence of erythrocyte blood biomarker research into other forms of dementias, such as vascular dementia, frontotemporal lobe dementia, Parkinson’s disease dementia, and dementia with lewy bodies. This review suggests that erythrocytes appear just as relevant as other blood compartments in this area of biomarker research. Importantly, this review has highlighted that further research must be done into exploring band 3, IgG, Aβ, calpain-1, and Hsp90 erythrocyte protein levels in mild cognitive impairment to assess their usefulness as preclinical blood biomarkers to dementia. Larger studies are needed to confirm the clinical utility of all these erythrocyte biomarkers in relation to dementia subtypes, correlation with clinical symptoms of both cognitive and behavioral changes in dementia, as well as how these biomarkers are affected with age, gender, comorbidity, and pharmacological treatments.
Footnotes
ACKNOWLEDGMENTS
This study is part of a MSci thesis (AS). DL and PK helped with the review of literature whilst doing their summer undergraduate attachment. We thank Mr. Kevin O’Malley (Department of Engineering, University of Glasgow) for graphical design of ![]() . We thank the Alzheimer’s Society, London (UK) (EBM-L and RK) for supporting this research.
. We thank the Alzheimer’s Society, London (UK) (EBM-L and RK) for supporting this research.