
Abstract
Psychosocial stress and physical, cognitive, and social activity predict the risk of cognitive decline and dementia. The aim of this study was to elucidate brain-derived neurotrophic factor (BDNF), irisin, and the kynurenine pathway (KP) as potential underlying biological correlates. We evaluated associations of irisin and the KP with BDNF in serum and with cognition, stress, and activities. Furthermore, changes in serum concentrations of BDNF, irisin, and KP metabolites were investigated after physical or cognitive training. Forty-seven older adults at risk of dementia were assigned to 10 weeks of physical training, cognitive training, or a wait-list control condition. Previous physical, cognitive, and social activities and stressful life events were recorded; global cognition, episodic memory, and executive functions were assessed. Serum levels of L-kynurenine, kynurenic acid, 3-hydroxykynurenine (3-HK), and quinolinic acid (QUIN) were determined by validated assays based on liquid chromatography coupled to tandem mass spectrometry. BDNF and irisin serum levels were determined with enzyme-linked immunosorbent assays. BDNF and irisin correlated positively with global cognition and episodic memory, while the neurotoxic metabolite QUIN correlated negatively with executive functions. Stressful life events were associated with reduced BDNF and increased 3-HK. 3-HK decreased after cognitive training, while BDNF tended to increase after physical training. This suggests that psychosocial stress as well as cognitive and physical training may impact BDNF serum levels and the KP. Irisin and QUIN may constitute novel serum biomarkers of cognitive impairment, in addition to BDNF. Larger scale trials are needed to replicate and extend these novel findings.
Keywords
INTRODUCTION
A number of lifestyle factors influence the risk of cognitive decline and dementia in old age. While a high amount of physical, cognitive, and social activities lowers the risk of cognitive decline and dementia [1], psychosocial stress can increase the risk [2, 3]. In recent years, an increasing number of studies focused on unravelling the underlying biological mechanisms of lifestyle effects, with the aim of identifying novel treatment strategies for an increasing number of older adults with neurodegenerative diseases.
The neurotrophin brain-derived neurotrophic factor (BDNF) is a probable mediator of lifestyle effects on cognition. BDNF plays a major role in neuroplasticity [4] and is linked to learning and memory processes [5, 6]. Reduced peripheral BDNF concentrations have been reported in patients with Alzheimer’s disease (AD) [7, 8] and in individuals with mild cognitive impairment (MCI) [9, 10], but results are mixed [11]. Experimental studies demonstrated an involvement of BDNF in the positive effects of physical exercise on cognition [12]. Some studies also reported BDNF enhancements in serum after cognitive interventions [13–15]. In contrast, serum BDNF seems to be reduced in individuals with high stress loads at work [16] or a higher number of stressful life events [17]. Recently, exercise-induced BDNF alterations have been linked to the myokine irisin and the kynurenine pathway (KP) in animal studies.
The discovery of the “exercise hormone” irisin attracted a great deal of attention [18]. Irisin was identified as a communicator between the skeletal muscle and adipocytes, and thus a potential bearer of positive effects of physical exercise on other target organs outside the muscle [19]. Boström et al. [18] demonstrated that irisin is cleaved from the transmembrane receptor fibronectin type III domain containing 5 (FNDC5) in skeletal muscle and secreted into the periphery as a myokine, from where it acts on adipocytes. Ten weeks of physical training resulted in increasedlevels of irisin in plasma of humans [18]. Subsequentstudies confirmed altered irisin levels after acute bouts of exercise [20, 21]. Results were, however, more equivocal with respect to physical training interventions over several weeks [22–25].
The observation that irisin administration increased the proliferation of hippocampal cells in vitro [26] and FNDC5 expression resulted in elevated irisin concentrations and BDNF gene expression in cell-culture [27] suggested that irisin might also constitute a potential therapeutic target in neurodegenerative disorders [28–30]. Despite that, associations between cognition and irisin in older adults or patients with neurocognitive disorders have not yet been investigated. Two studies reported correlations between cognition and irisin in serum in younger healthy adults, with one finding positive associations of irisin with the Mini-Mental State Examination (MMSE) [31], the other finding negative associations with measures of executive functions [32].
Another link between lifestyle factors, cognition and neuroplasticity constitutes the KP, the major route of tryptophan (TRP) metabolism. The essential amino acid TRP is metabolized into L-kynurenine (KYN). The KP itself consists of two branches with neuroactive metabolites as key products: One branch is initiated by kynurenine 3-monooxygenase (KMO)— an enzyme that catalyzes compounds with rather neurotoxic characteristics, including 3-hydroxykynurenine (3-HK) [33] and quinolinic acid (QUIN) [34]. The other branch of the KP leads to the formation of kynurenic acid (KYNA) through kynurenine aminotransferases (KAT I-IV). Depending on its concentration, KYNA can have neuroprotective effects and is able to counteract QUIN-induced neurotoxicity in the brain [35]. Furthermore, KYNA is not able to pass the blood-brain barrier, in contrast to KYN [36]. Thus, the catabolism of KYN to KYNA in the periphery may also reduce the amount of (deleterious) KYN passing tothe brain.
Altered levels in kynurenine metabolites have been detected in neurodegenerative disorders [37, 38]. In AD, an increased activation of the KP and a shift toward the neurotoxic metabolites with higher levels of 3-HK and QUIN was found in the hippocampus [39, 40] as well as in peripheral tissue [41–43]. Cognitive performance was negatively associated with QUIN and positively associated with KYNA concentrations in plasma [41]. Agudelo and colleagues [36] recently demonstrated in an animal study that stress increased the activity of KMO in plasma and reduced BDNF in the hippocampus. Both actions could be prevented by overexpression of the peroxisome proliferator-activated receptor gamma coactivator 1 alpha (PGC-1α) in the muscle, as a model for physical exercise. PGC-1α overexpression as well as physical exercise increased metabolism along the KYNA branch, demonstrated by an increased gene expression of KATs in the muscle and corresponding increases of KYNA in plasma. Similarly, 3 weeks of physical training in humans were also associated with increases inKATs [36].
In sum, BDNF has been related to effects of stress, physical and cognitive training, and an activity-enriched environment on cognition, although evidence in humans is mixed. Irisin and the KP may be further underlying biological mechanisms of lifestyle effects, including physical and cognitive activity, on cognition and may be linked to BDNF increases in the brain.
This study investigated older adults at risk of AD, who completed an elaborate assessment of lifestyle parameters as well as neuropsychological tests at baseline, and received a 10-week period of either physical training (PT), cognitive training (CT), or a wait-list control (WLC) condition. We aimed at evaluating BDNF, irisin, and kynurenine metabolites as potential underlying biological mechanisms of associations between lifestyle risk and protective factors of dementia and cognition and of training effects on cognition. We therefore tested the hypotheses that 1) Irisin and the KP metabolites are associated with BDNF in serum; 2) BDNF, irisin, and the neuroprotective KP metabolite KYNA are associated with better cognitive performance, while the neurotoxic KP metabolites (3-HK and QUIN) are associated with poorer cognitive performance at baseline; 3) BDNF is negatively and the neurotoxic KYN metabolites are positively associated with stress; and 4) BDNF, irisin and KYNA are positively associated, while the neurotoxic KP metabolites are negatively associated with the amount of physical, cognitive, and social activities. We further evaluated 5) whether a short physical or cognitive training program altered serum levels of irisin, the KP metabolites and BDNF as early biomarkers.
MATERIALS AND METHODS
Study design
The results reported here are part of a clinical trial (ClinicalTrials.gov Identifier NCT01061489, registered February 2, 2010), of which we previously reported cognitive outcomes [44] and diffusion tensor imaging data [45]. Here, we report associations and training-induced alterations of blood-basedbiomarkers.
Participants
The study population has previously been described in detail [44]. In brief, subjects were recruited in the Memory Clinic of the University Hospital Ulm, Germany and the Center for Psychiatry Reichenau, Germany or via public advertisements. Inclusion criteria were age of 55 years or older, subjective memory complaints and either objective (German version of the California Verbal Learning Test [46]: average of learning and long-delayed free recall trials below –1 SD of the age norm) or clinically apparent memory impairment (e.g., increased difficulty in relocating objects, keeping appointments, remembering conversations or events), and fluency in the German language. Exclusion criteria were any psychiatric or neurologic disorders, severe hearing or visual impairment, physical impairment which would have prevented participation in the PT program, changes in antidementive or antidepressive medication before study initiation, and moderate or severe dementia (MMSE <20; see Fig. 1).
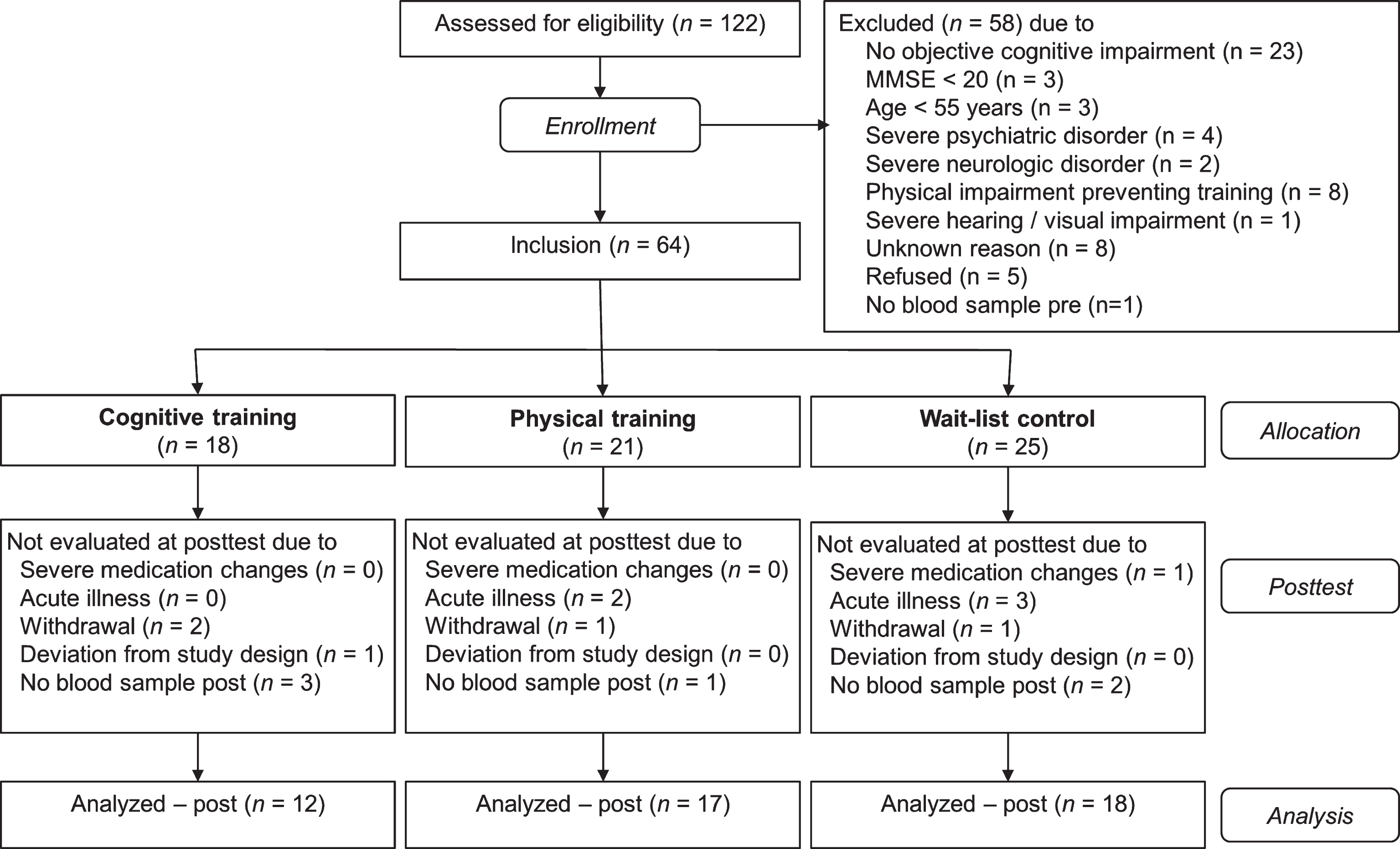
Flow of participants within the cognitive training, physical training, and wait-list control group.
Procedure
The study was approved by the Ethics Committees of the Universities of Konstanz and Ulm, Germany. Written informed consent was obtained from participants prior to study participation. Cognitive tests, a diagnostic interview with self-report questionnaires, and the collection of blood were usually performed on two appointments for each, the pre- and the post-test. One to 4 weeks after the pre-test, the 10-week intervention period started, followed by the post-test up to 4 weeks after the last trainingsession.
Groups were matched with respect to age, education, gender, and cognitive status (MMSE) using a minimization approach, in order to avoid a selection bias. The investigators who conducted the neuropsychological assessments at post-test and who performed the BDNF and irisin measurements were blinded to the subjects’ group assignments. Investigators conducting the KP measurements were not blinded to group allocation in order to evenly distribute the samples of the three groups on the measurement plates to avoid a measurement bias.
Assessment of biological parameters in serum
Blood collection, sample pre-processing and storage
Venipuncture was performed between 8 : 30 and 11 : 00 a.m., prior to which participants were asked to refrain from physically demanding activities and were seated for a resting period of at least 5 min before venipuncture. Fasting was not mandatory. Blood collection time at pre- and post-test was kept constant for each participant. Blood was collected in 7.5 ml serum tubes (Sarstedt, Nümbrecht, Germany) containing beads coated with a clotting activator (silcate). Blood samples were centrifuged 2.5 h (±30 min) post collection for 4 min at 2700 g and 4°C. Serum samples were frozen at –80° C until analysis.
BDNF
Serum levels of total BDNF were measured using the Enzyme-linked Immunosorbent Assay (ELISA) kit BDNF Emax ImmunoAssay Systems (Cat #: G7610, Promega Corporation, Madison, WI, USA) according to the manufacturer’s instructions. All samples of each participant and each time-point (pre, post) were assayed in duplicate on each plate, in order to test the intra-assay variation. Serum samples were diluted 1 : 100 in blocking buffer and acidified before used in the system. A BDNF standard was used on each plate to generate a linear standard curve ranging from 7.8–500 pg/ml. The absorbance at 450 nm was recorded within each well using an automated microplate reader (Biotek). The samples were processed on 4 plates in total. Samples, which were to be compared, i.e., pre- and posttest samples of each participant, were processed on the same plate. Four samples, in which the absorbance of the duplicates differed by 100 pg/ml or more, were excluded from the analyses.
Irisin
Irisin concentrations in serum were determined using a competitive, commercially available ELISA kit (Phoenix, EK-067-52) according to the manufacturer’s instructions. Samples were measured in duplicate; given values are averages. To determine irisin concentrations, absorbance at 450 nm wavelength was measured using a spectrophotometer (Thermo Scientific, Multiskan FC).
Kynurenine metabolites
Measurements were conducted at Boehringer Ingelheim Pharma GmbH & Co. KG, DMPK Germany, Biberach an der Riss, Germany. Quantification of serum levels of TRP and its catabolites KYN, KYNA, 3-HK, and QUIN was performed by validated assays based on liquid chromatography tandem mass spectrometry. TRP, KYNA, 3-HK, and QUIN concentration levels in serum samples were quantified together in one assay, while KYN serum levels were quantified separately. The assays comprised sample clean-up by protein precipitation followed by reversed-phase chromatography and mass spectrometric detection in the positive ion multiple reaction monitoring mode using the deuterated analogues of the analytes, namely [D5] tryptophan, [D4] kynurenine, [D5] kynurenic acid, [D3] 3-hydroxykynurenine, and [D3] quinolinic acid as internal standards. The lower limits of quantification in serum were 2000 nM for TRP, 770 nM for KYN, 5 nM for KYNA, 20 nM for 3-HK, and 50 nM for QUIN. Assay accuracy (in terms of relative deviation, dev., from nominal concentrations) and precision (in terms of coefficient of variability, CV, of multiple measurements) were determined for each analyte by the fourfold analysis of quality control samples at four concentration levels. Assay accuracy and precision were <6.1% (dev.) and <8.7% (CV) for TRP;<6.7% (dev.) and <14.1% (CV) for 3-HK;<23.2% (dev.) and 5.3% (CV) for KYN; <11.3% (dev.) and 20.1% (CV) for KYNA; and <14.1% (dev.) and <8.6% (CV) for QUIN.
Cognitive assessment
Global cognition, memory functions, and attentional and executive functions were assessed with an extensive neuropsychological test battery. Principal component analysis served to construct these three component scores (see [44]). All variables were z-standardized by using the baseline data. For each participant the two component scores (memory functions and attention/executive functions) were built as the weighted average of the standardized variables with loadings of at least aij = 0.40 on the respective component. The global cognition score represents the average of the two component scores.
The neuropsychological assessment included German versions of the MMSE [47], subtests of the test battery of the Consortium to Establish a Registry for Alzheimer’s Disease [48], the Alzheimer’s Disease Assessment Scale – Cognitive Subscale [49], and an adapted German version of the California Verbal Learning Test [46] as well as the digit-span and digit-symbol-coding test of the Wechsler Adult Intelligence Scale [50] and the working memory subtest of the Everyday Cognition Battery [51].
Assessment of lifestyle protective and risk factors
Physical, cognitive, and social activities
The Community Healthy Activities Model Program for Seniors Physical Activity Questionnaire for Older Adults [52] was applied to assess regular physical, cognitive, and social activities of the participants. The questionnaire assesses the frequency and duration of 40 activities in a typical week within the previous 4 weeks. The activities were categorized into physical, cognitive, and social activity domains, as reported previously [44]. A score for each activity domain was built, reflecting the percentage of performed activities in relation to the possible number of activities in this domain. Then, an overall activity score was built by averaging the three domain scores.
Stressful life events
The Clinician-Administered PTSD Scale Life Events Checklist [53] was applied to assess the number of potentially traumatic life events. An event (personally experienced or witnessed) is considered traumatic if it poses a potential threat to life or physical integrity (criterion A1 for posttraumatic stress disorder) and is accompanied by the experience of intensive anxiety, helplessness, or horror (criterion A2). Regarding 19 event types, participants were asked whether they had experienced or witnessed the event at least once in their life and to shortly describe it including the experienced emotions. Two sum-scores were built, one comprising all events fulfilling criterion A1 (critical life events) and one including only those events which also fulfilled criterion A2 (traumatic life events).
Depressive symptoms
The Geriatric Depression Scale-15 [54] was used to determine depressive symptoms. Scores over 5 are indicative of mild to moderate depression; scores over 10 indicate severe depression.
Other potential influencing factors
We assessed age, education in years, the number of cigarettes smoked per day, and the number of glasses of alcohol consumed per day in a standardized interview and measured height and weight to calculate the body mass index.
Training interventions
Cognitive training
The CT group was asked to complete 1-h computer-based training sessions five times per week for a total of 10 weeks (i.e., 50 sessions in total), which were carried out individually at the participants’ homes. The training was an adapted and translated German version of a program developed by the Posit Science Corporation, San Francisco, CA and focuses on the training of auditory discrimination and working memory (for detailed descriptions see [44, 55]).
Physical training
The PT group was asked to attend 1-h sessions twice a week in groups of five to ten participants and three 20-min sessions per week at home for 10 weeks (i.e., 20 group- and 30 home-based sessions in total). The multimodal PT program included endurance, coordination, balance, flexibility, and strengthening components, embedded into an imaginary journey. A similar program with the same structure yielded beneficial effect on cognition in frail nursing-home residents [56].
Wait-list control group
Participants of the WLC group were asked to continue their daily routine as usual and were offered to take part in one of the training programs after their study participation.
Statistical analyses
Statistical analyses were carried out with R version 3.1.2 [57]. Baseline group differences in continuous variables were evaluated with one-way analyses of variance. Baseline differences in categorical variables were analyzed with χ²-tests.
Associations of irisin and the KP measures (KYN, KYNA, 3-HK, QUIN) with cognition and with risk and protective lifestyle factors were calculated as Pearson’s product-moment correlations. As BDNF values were not normally distributed, all associations with BDNF were calculated with Spearman rank correlations (see also [58]). Significant associations with measures of cognition are also reported after statistically accounting for influencing covariates, by including the covariates into a multiple regression analysis (for irisin and KP measures) or calculating partial correlations after partialling out the covariates (for BDNF).
To evaluate effects of training on irisin and KP measures, linear mixed effects models were conducted with Group (PT, CT, WLC) and Time (pre, post) as fixed effects and Subject as a random intercept, using the nlme package 3.1–119 in R. Significant Group×Time interactions indicated effects of training on the biological measures. Changes from pre- to post-test within each group were analyzed with paired t-tests. Effects of training on BDNF were analyzed with Wilcoxon tests within each group.
RESULTS
Subject characteristics
The study population consisted of 20 male and 27 female participants with a mean age of 71.2 years (SD = 6.0, range 60–88 years), a mean education time of 14.1 years (SD = 3.4), and a mean MMSE score of 28.0 (SD = 1.9). The sample included mostly individuals with MCI (n = 32), 11 participants with no objective memory impairment, and four participants with probable beginning dementia. Three participants (n = 1 in each group) had Geriatric Depression Scale scores indicative of mild to moderate depression (6–8 points). Self-reported leisure activity was high (6–20 regular activities, M = 13.77, SD = 3.70) considering the age of the sample. The participants reported 0 to 11 critical life events (M = 3.74, SD = 2.49), of which 0 to 5 were traumatic (M = 1.23, SD = 1.35). The three groups (CT, PT, and WLC) did not differ in any demographic characteristics, cognitive status, lifestyle, or biomarker concentrations (see Table 1). Thirty-four participants reported current medical conditions, most of them with one (n = 10) or two (n = 14) diagnoses, the most frequent being hypertension (n = 14) and arthrosis (n = 9). Thirty-six subjects reported current medication intake, most frequently antihypertensives (n = 15) or thyroidhormones (n = 9). Six participants took antidementive, two participants antidepressive medication.
Baseline characteristics for each of the three intervention groups
KYN, KYNA, 3-HK, and QUIN are measured in nM, irisin is measured in pg/ml, and BDNF is measured in ng/ml. 3-HK, 3-hydroxykynurenine; BDNF, brain-derived neurotrophic factor; CAPS, Clinician-Administered PTSD Scale; CT, cognitive training group; GDS-15, Geriatric Depression Scale-15; KYN, L-kynurenine; KYNA, kynurenic acid; MMSE, Mini-Mental State Examination; PT, physical training group; QUIN, quinolinic acid; WLC, wait-list control group.
Irisin measurements were missing for five participants, KP metabolite measurements were missing for six participants, as there were not enough serum samples available. BDNF measurements were missing for five subjects at baseline. For pre-post comparisons, the BDNF data of a further 6 subjects were excluded, as post-data were missing. Test-retest reliability between pre- and post-measurements was reasonable for all biomarkers (Irisin: r = 0.67; BDNF: ρ= 0.70; KYN: r = 0.68; KYNA: r = 0.55; 3-HK: r = 0.72; QUIN: r = 0.83).
Irisin but not the kynurenine metabolites are associated with BDNF
To evaluate proposed links of irisin and the KP with BDNF, baseline associations between the biomarkers were calculated. Irisin levels correlated positively with BDNF levels (ρ= 0.32, p = 0.05), while the kynurenine metabolites were not associated with BDNF (ps≥0.45).
Irisin, QUIN, and BDNF levels correlate with measures of cognition
To test the hypothesis whether irisin, KP metabolites, and BDNF are associated with cognitive performance, correlations with global cognition as well as with the composite scores were calculated at baseline (see Table 2). Global cognition was significantly associated with irisin serum concentrations (r = 0.37, p = 0.02) as well as a with BDNF serum concentrations (ρ= 0.33, p = 0.04), but not with any of the KP metabolites (ps≥0.14). Within the two cognitive component scores, memory correlated significantly with irisin (r = 0.45, p = 0.003) and BDNF (ρ= 0.36, p = 0.02), while the component of attention/executive functions was inversely correlated with QUIN (r = –0.31, p = 0.05; see Fig. 2).
Baseline associations of biological parameters with measures of cognition
Associations with BDNF are Spearman rank correlations, all other associations are Pearson product-moment correlations. 3-HK, 3-hydroxykynurenine; Attention/EF, attention/executive functions; BDNF, brain-derived neurotrophic factor; KYN, L-kynurenine; KYNA, kynurenic acid; QUIN, quinolinic acid. **p < 0.01, *p < 0.05.

Baseline associations of biomarkers with composite measures of cognition. Higher brain-derived neurotrophic factor (BDNF) and irisin serum levels were significantly associated with better memory performance (ρ= 0.36, p = 0.02; and r = 0.45, p = 0.003, respectively). Higher levels of quinolinic acid (QUIN) in serum were associated with poorer performance in a composite score of attention and executive functions (r = –0.31, p = 0.05).
After statistically accounting for age and education, the associations of global cognition with BDNF (p < 0.01) as well as of memory with irisin (p = 0.046) and BDNF (p < 0.01) remained significant, while the associations of global cognition with irisin was no longer significant (p = 0.21). The association between attention/executive functions and QUIN remained marginally significant after accounting for either age and education (p = 0.07) or alcohol consumption (p = 0.10).
Lifetime stress is associated with lower BDNF and higher 3-HK levels
To assess associations of the biomarkers with lifetime stress as a risk factor of AD we used the Clinician-Administered PTSD Scale Life Events Checklist. The number of experienced critical life events, which pose a threat to life or physical integrity, was negatively associated with BDNF (ρ= –0.40, p = 0.009) and tended to positively correlate with 3-HK levels (r = 0.29, p = 0.07, see Fig. 3 and Table 3). Associations with the number of traumatic life events which were also connected with the experience of extreme fear or helplessness, were not significant.

Baseline associations of critical life events with 3-HK and BDNF serum levels. The number of critical life events was assessed with the Life Events Checklist of the Clinician-Administered PTSD Scale (CAPS), as a measure of lifetime psychosocial stress. There was a trend for a positive correlation of critical life events with 3-hydroxykynurenine (3-HK) serum levels (r = 0.29, p = 0.07) and a significant negative correlation with brain-derived neurotrophic factor (BDNF) serum levels (ρ= –0.40, p = 0.009).
Baseline associations of biological parameters with lifestyle risk and protective factors and covariates
Associations with BDNF are Spearman rank correlations, all other associations are Pearson product-moment correlations. 3-HK, 3-hydroxykynurenine; BDNF, brain-derived neurotrophic factor; CAPS, Clinician-Administered PTSD Scale; GDS-15, Geriatric Depression Scale-15; KYN, L-kynurenine; KYNA, kynurenic acid; QUIN, quinolinic acid. **p < 0.01, *p < 0.05, #p < 0.10.
No association of self-reported activity with the investigated biomarkers
The amount of regular physical, cognitive, and social activities before study participation was assessed with a questionnaire. The amount of activity was not associated with serum levels of irisin, KP metabolites, or BDNF at baseline (see Table 3).
Training-induced alterations of BDNF and 3-HK levels
There was a trend for an increase in BDNF levels from pre to post in the PT group (Wilcoxon V = 25, p = 0.09). BDNF also increased in the CT group, but the difference did not reach significance (V = 11, p = 0.11; see Fig. 4 and Table 4). When taking the two training groups (CT and PT) together, the BDNF serum levels significantly increased from pre- to post-test (V = 66, p = 0.02), while there was no alteration in the WLC group.
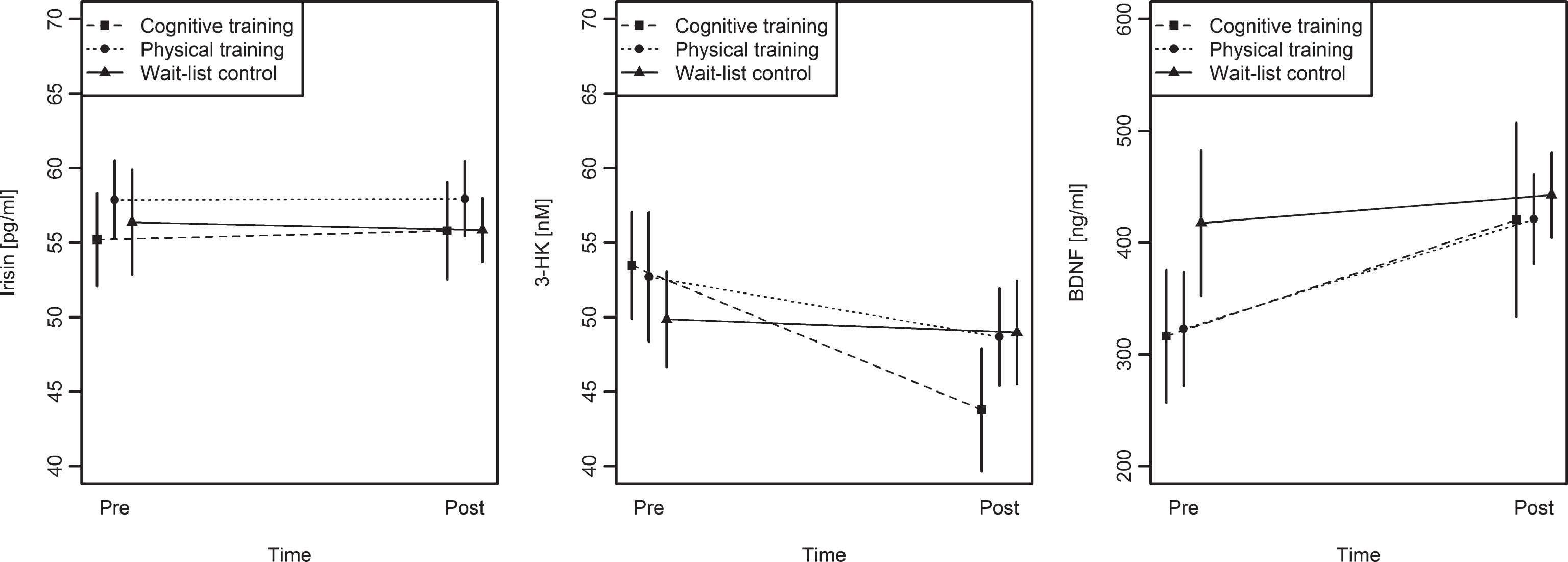
Pre to post changes in irisin, 3-HK and BDNF serum levels for the three groups. Irisin levels remained unchanged in all three groups. 3-HK levels decreased significantly from pre to post within the cognitive training group in comparison to the wait-list control group. BDNF levels tended to increase within the physical training group, while levels remained unchanged in the wait-list control group. Error bars depict standard errors of the mean.
Comparisons of biological parameters in the three intervention groups between pre- and post-test
KYN, KYNA, 3-HK, and QUIN are measured in nM, irisin is measured in pg/ml, and BDNF is measured in ng/ml. 3-HK, 3-hydroxykynurenine; BDNF, brain-derived neurotrophic factor; CT, cognitive training group; KYN, L-kynurenine; KYNA, kynurenic acid; QUIN, quinolinic acid; PT, physical training group; WLC, wait-list control group. adescriptive statistics for pre- and post-test expressed as median and interquartile range.
We found a significant Group×Time interaction on 3-HK, F(2,37) = 3.25, p = 0.05 (see Fig. 4). 3-HK decreased significantly in the CT group, t(9) = 3.17, p = 0.01, and tended to decrease in the PT group, t(13) = 1.76, p = 0.10, while it remained unchanged in the WLC group, t(15) = –0.11, p = 0.91. KYNA decreased significantly within the CT group, t(9) = 2.23, p = 0.05, but a Group×Time interaction was not significant, F(2,38) = 0.36, p = 0.70.
There were no other significant Group×Time interactions for modelling kynurenine metabolites (ps > 0.18). There was also no significant Group × Time interaction on irisin levels, F(2,39) = 0.05, p = 0.95.
DISCUSSION
We aimed to evaluate potential neurobiological correlates of lifestyle- and training-related associations with cognition in a sample of older adults at risk of dementia. We found significant associations of BDNF, irisin, and the neuroactive kynurenine metabolite QUIN in serum with measures of cognition. Lifetime psychosocial stress, as a risk factor of cognitive decline and dementia, correlated with BDNF and 3-HK. In the interventional part of the study, we found significant decreases of the neurotoxic 3-HK after 10 weeks of cognitive training in comparison to a wait-list control group and a trend for an increase of BDNF after physical training.
Involvement of Irisin and the kynurenine pathway in neuroplasticity and cognition
Corroborating our hypotheses, we found that irisin levels were positively associated with BDNF in serum, a marker of neuroplasticity, and with cognition, especially with hippocampus-related memory functions. This adds to evidence of animal and in vitro studies, which suggested connections of irisin and its progenitor FNDC5 with BDNF and neuroplasticity in the central nervous system, as part of a pathway between physical exercise and cognition [26, 59]. In accordance with our results, a positive association of serum irisin levels with a screening measure of cognition as well as with BDNF had been demonstrated in young adults [31]. The herein observed associations between irisin, BDNF, and cognition might indicate BDNF as a mediator between irisin and cognition, as suggested by previous research in animals [27]. However, other mechanisms by which irisin is connected to cognition are also plausible, for instance by an influence of mitochondrial processes. In adipocytes, FNDC5 enhanced mitochondrial density [18]; and in rat cardiomyoblasts, irisin upregulated mitochondrial metabolism [60].
In our study, higher QUIN levels were associated with poorer performance in attentional and executive functions. Our results are in line with studies demonstrating an increased QUIN concentration in brain [40] and serum [41] of AD patients, with a negative association between QUIN serum levels and the clock-drawing test, a cognitive screening tool for dementia.
Associations of the kynurenine pathway and BDNF with lifetime psychosocial stress
In line with the hypothesis, we found reduced BDNF serum levels and higher 3-HK serum levels in individuals who reported stressful life events. This adds evidence to results of animal studies, which demonstrated stress-related changes in tryptophan metabolism, with a shift toward the KP and an increase of neuroactive KP metabolites in plasma and brain [36, 62], including increases of 3-HK [61]. The stress-related increases of 3-HK and reductions of BDNF observed here might contribute to the increased risk of cognitive deficits and dementia in stressed individuals [63, 64]. The number of experienced stressful life events was low, as expected within a non-psychiatric sample of this age [65]. Future studies should examine the associations with 3-HK and BDNF in samples with higher exposure to stress.
Biological alterations with physical or cognitive training
We observed increases in BDNF levels and reductions of 3-HK after 10 weeks of physical and cognitive training, respectively. Physical exercise-induced changes in BDNF concentrations have been reported in numerous animal studies, mainly within the hippocampus [66, 67]. Effects of physical trainingon peripheral BDNF in humans, including older adults [68, 69] and patients with MCI [70] were less consistent [71]. Changes in BDNF concentrations after 10 weeks of cognitive training did not reach significance in this study. Cognitive training with the BrainFitness program (as applied in this study) yielded BDNF enhancements in other patient populations [14, 15], but have not been reported in the context of neurodegenerative disease. Further research with larger sample sizes is needed to elucidate the underlying biological mechanisms of cognitive training, particularly in individuals with neurocognitive disorders.
We observed a decrease of the neurotoxic metabolite 3-HK in the cognitive training group and a tendency for a decrease in the physical training group, in comparison to the control group. Little literature exists with respect to training-induced alterations in kynurenine metabolism. Changes in kynurenine metabolism have been demonstrated after a longer period of physical training in mice and humans, namely an increase of KATs in skeletal muscle which convert KYN to KYNA [36]. However, other studies [72]— like ours— failed to find effects of physical activity on kynurenine metabolites in serum of humans. As KP alterations after cognitive training have not been reported before, the 3-HK reductions observed here were not expected and need further confirmation.
Irisin levels remained unchanged after physical training. This is in accordance with previous reports [25], although other studies reported effects of physical training regimens with similar duration as ours [18, 23]. It has been hypothesized that irisin levels only rise after acute bouts of physical exercise but are not chronically elevated in response to physical training programs [73]. This notion is supported by reports of only transient irisin enhancements after a bout of exercise with a subsequent return to initial levels within 24 h [20, 74]. Another exciting hypothesis is that irisin is only increased in states of energy need [20, 29], that is when exercise constitutes a challenge to present energy resources, for example, in untrained, sedentary individuals. The participants in our study were quite active before taking part in the physical training. Thus, energy demands of the training program may have been too low in these participants to yield elevations of irisin.
Notably, we did not find any significant training-related effects on cognition in this sample [44]. This seems contrary to the alterations in BDNF and 3-HK on the molecular level reported here. In contrast to alterations on the biomarker level, training-induced changes in cognition might have been masked by substantial retest-effects in all groups. Furthermore, alterations on the biomarker level might have been too small to transfer to improvements in cognition. Cognitive improvements during the same time-period were, however, correlated to the self-reported activity level of the participants’ lifestyles. Considering both the training-related alterations on the biomarker level and the lifestyle-related changes in cognition, another possibility is that changes at the molecular level are more likely to be influenced by short-term measures, such as the 10-week training programs, while changes at the cognitive level rather occur after longer term activity, as with an active lifestyle.
Limitations and outlook
By nature of the human study population, biomarker measurements were obtained in the periphery. Thus, we cannot infer that the observed alterations in biomarker concentrations in serum reflect those in the brain. However, regarding BDNF, close relationships between brain and serum concentrations of BDNF have been demonstrated in different species [75–77]. Furthermore, BDNF is able to pass the blood-brain barrier in both directions [78] and an interesting study recently demonstrated that the brain is the main source of exercise-induced BDNF elevations in the blood [79].
The validity of irisin ELISA measurements, in particular the identification of irisin in serum as a cleaved product of the transmembrane receptor FNDC5, has been doubted [80]. However, the immunoblot identification of irisin has recently been validated by mass spectrometry, which implicates ELISA validity [81]. BDNF measurements faced challenges due to a large variance. For this reason, statistical analyses including the BDNF measures were performed with non-parametrical, rank-based tests, as recommended by Ziegenhorn and colleagues [58]. This may have constrained the power to detect changes, e.g., after cognitive training. Unfortunately, no international recommendations or standard operating procedures with respect to BDNF measurements in serum exist and measurements may be influenced by the applied commercial assay [82]. Standardized procedures and validation for the assays for the determination of BDNF in human material are needed to improve the comparability of study results. The catabolite 3-HK turned out to be quite instable during measurements, which might imply influences of clotting time on the concentration. However, venipuncture and centrifugation procedures were kept constant and did not differ between groups. Furthermore, a high correlation between pre- and post-measurements indicated reasonably reliable measures. Clotting time was fairly long (2-3 h, but compare [82]), but was kept constant for all probes. Pre-analytic handling of the probes may influence the measurements of the biomarkers [82], which is why a standardized procedure is of utmost importance. The validity of the results in this study might be improved by a different pre-analytic procedure.
Other demographic or lifestyle factors, such as age, education, or the body mass index, may influence biomarker levels or their associations with cognition. We found associations of age with irisin and of alcohol consumption with some of the kynurenine metabolites. After statistically accounting for these variables most of the reported associations between biomarker concentrations and cognitive performance remained (at least marginally) significant. However, the association between irisin and global cognition lost significance after accounting for age, indicating that this association may be influenced by associations with age. Apart from that, serum concentrations of BDNF, irisin, and the kynurenine metabolites are probably not specifically associated to cognition, psychosocial stress, or training, as described here, but also to other diseases (e.g., adiposity [74] or depression [83]) or treatments (e.g., antidepressant medication [83]).
The limited sample size likely impeded the detection of small effects. Further research is needed to confirm our results and investigate the potential role of BDNF, irisin, and the KP in lifestyle- and training-effects on cognition in the context of dementia and other neurodegenerative disorders. In addition, other potential mediators of effects of physical and cognitive training on neuroplasticity and cognition should be examined. Understanding the biological mechanisms which connect an “active lifestyle” to enhanced brain health will strengthen the support for lifestyle changes and also open possibilities for the development of further treatment strategies, including pharmacological ones.
Conclusion
Associations of irisin and metabolites of the KP with BDNF and cognition on the one hand, and with psychosocial stress as well as cognitive or physical training on the other hand, indicate that thesebiological measures may constitute candidate mediators of lifestyle influences on cognition and dementia in old age. Further research in larger scale studies is necessary to confirm the results and elucidate the potential role of irisin and kynurenine metabolites in prevention strategies of neurodegenerative diseases.
Footnotes
ACKNOWLEDGMENTS
This study was supported by the Heidelberg Academy of Sciences and Humanities, Germany.Kynurenine metabolites were measured at Boehringer Ingelheim Pharma GmbH & Co. KG,DMPK Germany, Biberach an der Riss, Germany. We thank Rosine Gröschel, Nelli Hirschauer, Jens Kalchthaler, Anne Korzowski, Claudia Massau, Dörte Polivka, Karl Pröbster, Heike Riedke, Christina Schaldecker, and Christiane Wolf for support in blood collection and processing, subject recruitment, data acquisition, and training implementation.