
Abstract
Background:
Bone marrow-derived progenitor cells survey the vasculature and home to sites of tissue injury where they can promote repair and regeneration. It has been hypothesized that these cells may play a protective role neurodegenerative and vascular cognitive impairment.
Objective:
To evaluate progenitor cell levels in older adults with and without mild cognitive impairment (MCI), and to relate circulating levels to memory, brain volume, white matter lesion volume, and cerebral perfusion.
Method:
Thirty-two older adults, free of stroke and cardiovascular disease, were recruited from the community and evaluated for diagnosis of MCI versus cognitively normal (CN). Participants underwent brain MRI and blood samples were taken to quantify progenitor reserve using flow cytometry (CD34+, CD34+CD133+, and CD34+CD133+CD309+ cells).
Results:
Participants with MCI (n = 10) exhibited depletion of all CPC markers relative to those who were CN (n = 22), after controlling for age, sex, and education. Post-hoc age, sex, and education matched comparisons (n = 10 MCI, n = 10 CN) also revealed the same pattern of results. Depletion of CD34+ cells correlated with memory performance, left posterior cortical thickness, and bilateral hippocampal perfusion. Participants exhibited low levels of vascular risk and white matter lesion burden that did not correlate with progenitor levels.
Conclusions:
Circulating progenitor cells are associated with cognitive impairment, memory, cortical atrophy, and hippocampal perfusion. We hypothesize that progenitor depletion contributes to, or is triggered by, cognitive decline and cortical atrophy. Further study of progenitor cell depletion in older adults may benefit efforts to prevent or delay dementia.
INTRODUCTION
It is well established that vascular risk factors convey increased risk of cognitive decline in older adults [1]. Despite the role of vascular risk factors, very little is known about protective vascular factors in cognitive decline. One proposed mechanism of vascular protection involves circulating progenitor cells. Bone marrow-derived progenitor cells are frequently identified by expression of the hematopoietic stem cell marker, CD34, as well as the progenitor marker, CD133. Cells expressing both CD34 and CD133 are proangiogenic, and the subpopulation that also co-expresses CD309/VEGFR-2 (vascular endothelial growth factor receptor 2) represent endothelial progenitor cells, which have shown the ability topromote both angiogenesis and vasculogenesis [2, 3]. Collectively, these cells may promote brain health by homing to sites of cerebrovascular or neural injury and promoting angiogenesis and neurogenesis [4–11].
Despite the widely-hypothesized role of these cells in protection against cerebrovascular and neural injury, very few human studies have investigated their potential role in age-related cognitive decline, dementia, and aging brain health. One study using cell culture methods found that circulating angiogenic cells were depleted in Alzheimer’s disease (AD) patients, where their number correlated with global indices of cognitive impairment [12]. Other studies have further supported the notion that proangiogenic cells (CD34+CD133+) are reduced in AD [13]. There is also evidence that CD34+ cells in general may be reduced in AD, where they correlate with amyloid-β levels in the cerebrospinal fluid (CSF) [14]. More recently, multiple progenitor cell markers correlated with better cognitive performance in a large non-clinical sample of employees at a major university [15]. To date, no studies have investigated progenitor cells in relation to memory ability in older adults or whether these cells are depleted in mild cognitive impairment (MCI). There have also been no studies investigating whether these circulating progenitor cells correlate with MRI-based measures of aging brain health, such as cortical thickness and white matter lesion (WML) burden. Although studies have linked CD34+ cell levels to improved cerebral blood flow after stroke [16], no studies have examined their role in perfusion changes associated with normal and pathological aging independent of stroke.
The present study sought to fill these knowledge gaps by investigating circulating progenitor cell levels in relation to cognitive function in a sample of community dwelling older adults, and to further study how these cells relate to MRI-based indicators of brain volume, white matter disease and cerebralperfusion.
METHOD
Participants and setting
Thirty-four participants were recruited from the community through word-of-mouth, flyers, and community outreach events. All participants gave informed consent, and the study was approved by the University of Southern California (USC) Institutional Review Board (IRB). Study procedures were carried out in the Vascular Senescence and Cognition (VaSC) Laboratory, housed within the USC Department of Psychology. Participants were included if they were age 55 years or older and independently living with no history of clinical stroke, dementia, learning disability, traumatic brain injury, or other systemic or neurological illness or treatment that may impact central nervous system function. History of vascular risk factors was determined by clinical interview and objective measures, including blood pressure, body mass index (weight in kg/height in m2), dyslipidemia (history of elevated total cholesterol, low density lipoprotein cholesterol or triglycerides OR reduced high density cholesterol), diabetes (history of type I or II), history of cardiovascular disease (angina, intermittent claudication, myocardial infarction, stent placement, coronary artery bypass graft), or history of transient ischemic attack. Seated blood pressure was measured twice from each arm on two separate days (prior to neuropsychological testing for visit 1 and prior to MRI scan for visit 2), and systolic and diastolic pressures were averaged across these measures (pulse pressure was calculated as systolic minus diastolic pressure and mean arterial pressure was calculated as diastolic blood pressure plus one-third of the pulse pressure).
Two participants were excluded after clinical interview revealed additional factors that may impact progenitor cell function or neurocognition (one reported a history of human immunodeficiency virus and the other reported a family history of Huntington’s disease). After these participants were excluded, 32 participants remained in study.
Blood draw and flow cytometry
Venipuncture was performed after an overnight fast on the morning before neuropsychological testing. Blood was drawn into K3EDTA-coated tubes and 15 mL of anticoagulated whole blood was separated and diluted 1 : 1 with Dulbecco’s Phosphate Buffered Saline (DPBS) + 2% fetal bovine serum (FBS). Diluted blood was overlaid on Histopaque-1077 (Sigma Aldrich). Peripheral blood mononuclear cells (PBMCs) were isolated by density gradient centrifugation according to manufacturer instructions, and washed twice with DPBS + 2% FBS at 120×g for 10 min and 300×g for 8 min at room temperature. PBMCs were counted on a hemacytometer. 1 million cells were transferred to an unstained tube blank, and the remainder was transferred to the stained tube where 5 μL of Human BD Fc Block (BD Biosciences) was added and incubated for 10–15 min. An additional 1 μL of each of the following antibodies were then added: (1) CD34-PE-Vio770 (clone: AC136, Miltenyi Biotec), (2) CD133-VioBright FITC (clone: AC133, Miltenyi Biotec), (3) PerCP/Cy5.5-CD309 (clone: 7D4-6, BioLegend). Tubes were incubated in the dark for 30 min at 4°C. Samples were washed with 3 mL PBS, centrifuged at 300×g for 8 min, and fixed with 2% formaldehyde in PBS until analysis. Compensation controls were conducted using AbC Total Antibody Compensation Bead Kit (ThermoFisher). Samples were acquired on a BD LSR II flow cytometer and analyzed on FlowJo software. Within the lymphocyte gate, positive gates for each marker were established over their fluorescence-minus-one bounds. Progenitor cell populations of interest included: CD34+, CD34+CD133+, and CD34+CD133+CD309+. Progenitor cell concentrations were calculated as a percentage of total cells within the lymphocyte gate.
APOE genotyping
APOE genotyping was conducted on the blood cell pellet fraction obtained from plasma separation. DNA was isolated from the pellet fraction using the PureLink Genomic DNA Mini Kit (Thermo Fisher). Genotyping was conducted on isolated DNA using the TaqMan SNP Genotyping Assay (Thermo Fisher) on an Applied Biosystems 7300 Real Time PCR System. APOE gene SNPs were assessed for dbSNP IDs rs429358 and rs7412. Allelic discrimination was conducted using the included qPCR software. The APOE ɛ4 allele was designated as rs429358-C + rs7412-C.
Neuropsychological testing and MCI diagnosis
After the venipuncture, participants were fed a light snack before undergoing comprehensive neuropsychological testing, including assessment of global cognition, memory, attention/executive function, language, and visuospatial abilities. The Mattis Dementia Rating Scale was used to screen participants for dementia (raw score >126). Participants were classified as cognitively normal (CN) if they did not meet published neuropsychological criteria for MCI [17]. Specifically, MCI was diagnosed when participants scored >1 standard deviation below norm-referenced scores on >1 measure within a domain or >2 measures across domains. The neuropsychological battery used for diagnosis utilized 3 tests per domain of function (Supplementary Table 1). Numerous studies have previously established the validity of this neuropsychological approach to MCI and have shown improved stability [18], reduced false positive/negative rates [19, 20], and greater correlation with biomarkers of risk relative to other approaches [17].
Brain volumes and white matter lesions
A subset of participants (n = 30 out of 32; 2 excluded due to MRI contraindications) returned within two weeks for a second visit to undergo structural MRI of the brain on a 3T Siemens Prisma magnet. All participants who were scanned underwent a T1-weighted magnetization prepared rapid gradient-echo (MPRAGE) sequence (Scan parameters: TR = 2300 ms, TE = 2.98 ms, TI = 900 ms, slice thickness = 1.20 mm, flip angle = 9°, field of view = 256 mm) for evaluation of brain volumes, and a subset of participants (n = 25) also underwent T2-fluid attenuated inversion recovery (FLAIR) sequence (Scan parameters: TR = 10000 ms, TE = 91 ms, TI = 2500 ms, slice thickness = 5.0 mm, flip angle = 150°, field of view = 220 mm) for evaluation of WML volume.
For region-of-interest (ROI) analysis, post-processing of T1 scans was accomplished in FreeSurfer 5.3 using a semi-automated segmentation algorithm for quantification of bilateral hippocampal volumes (FreeSurfer 6.0), precuneus and posterior cingulate. Previous studies have detailed the specifics of FreeSurfer processing [21, 22]. After automated segmentation, each individual subject was checked for any inaccuracies or misclassifications; manual corrections were made as needed with FreeSurfer’s built-in editing tools, cases were then re-processed, and resulting volumes were used for analyses.
White matter lesions were segmented with the lesion growth algorithm [23] implemented in the LST toolbox version 2.0.15 (http://www.statistical-modelling.de/lst.html) for SPM12 (Wellcome De-partment of Cognitive Neurology, London, UK; http://www.fil.ion.ucl.ac.uk/spm/software/spm12). An optimal initial threshold was determined via visual inspection, and manual quality control check ensured no gross over- or under-estimation.
For voxel-level indexing of cortical volume, voxel-based morphometry (VBM) analyses were conducted using SPM12, as previously described [24]. Briefly, T1-weighted images were segmented into grey and white matter tissue classes using SPM12’s unified segmentation procedure, spatially normalized, and smoothed with an 8-mm full-width at half-maximum (FWHM) isotropic Gaussian kernel; the resulting grey matter images were checked for sample homogeneity to identify the presence of any potential outliers. Voxel-wise multiple regression was conducted with progenitor cell count as independent variable, grey matter map as dependent variable, and age, sex and total intracranial volume as covariates. As this was an exploratory analysis, we corrected for family-wise error in an initial analysis, but also conducted analyses with the cluster level significance threshold set at uncorrected p < 0.001.
Cerebral perfusion (arterial spin-labeling MRI)
Arterial spin-labeling (ASL)-MRI images were acquired on 28 participants using the Siemens PICORE-based pulsed ASL (PASL) sequence with the following parameters: TR = 3000 ms, TE = 12 ms, slice thickness = 4 mm, # of slices = 23, flip angle = 90°, field of view = 256 mm, TI1 = 700 ms, TI2 = 1900 ms, # of measurements = 105 (1 M0 image and 52 pairs of tag-control images), and total scan duration = 5 : 20. Images were processed with the ASLtbx Data Processing Toolbox (Ze Wang, University of Pennsylvania, https://cfn.upenn.edu/ zewang/ASLtbx.php) as implemented in SPM12 within MATLAB [25]. M0, ASL, and T1 images were first reoriented to ensure anterior commissure-posterior commissure alignment. ASL images were then motion corrected, coregistered to corresponding T1-weighted structural images, and spatially smoothed with a 6 mm FWHM isotropic Gaussian kernel. Pairwise tag-control subtractions resulted in 52 tag-control pairs, and these images were averaged to generate mean quantitative CBF maps, from which global mean perfusion (ml/100 g/min) for each subject was calculated.
Mean CBF images were normalized to MNI template space for ROI analyses of regions including bilateral hippocampus and posterior cingulate. ROIs were selected and exported from the Automated Anatomical Labeling (AAL) atlas using the WFU PickAtlas Toolbox (ANSIR Lab, Wake Forest University School of Medicine, http://fmri.wfubmc.edu/software/pickatlas) in MATLAB [26]. Once ROIs were defined, mean CBF values were extracted for individual ROIs using the REX toolbox (Gabrieli Lab, Massachusetts Institute of Technology, http://gablab.mit.edu/index.php/news/95-gablab-site/gablab/people/swg/83) in MATLAB.
Statistical analyses
Data were initially screened for outliers and departures from normality using indices of skewness and kurtosis. Log10-transformation was required to normalize the distribution of all progenitor cell counts and WML volumes.
Participant groups (CN versus MCI) were compared on demographics and traditional vascular risk factors using independent sample t-tests and chi-square tests. Age, sex, and vascular risk factors were also examined as potential contributors to progenitor cell counts. Progenitor cell counts were compared between groups using ANCOVA controlling for age, sex, and education level. Hierarchical multiple linear regression analyses examined relationships between progenitor cell counts and indicators of regional brain volume and perfusion in two regions previously implicated in cognitive decline: hippocampus and posterior cingulate. These limbic and cortical regions were selected due to their role in MCI and AD. Hierarchical regression models were also used to investigate progenitor cell counts in relation to WML volumes. For volumetric ROI and WML analyses, age, sex, and total intracranial volume were used as covariates.
RESULTS
Demographics and vascular risk factors
Participants who were diagnosed with MCI were significantly older (p = 0.02) than those identified as CN, but did not otherwise differ on any demographic or vascular risk factor (Table 1). All participants were free of cardiovascular disease and stroke, and exhibited relatively low levels of vascular risk. There were no significant differences in progenitor cell counts between those with or without any of the traditional vascular risk factors, between men and women, or between APOE ɛ4 carriers and non-carriers (all ps > 0.20; Supplementary Table 2). There was also no significant correlation between any of the progenitor cell counts and age, body mass index, or any blood pressure measure (Supplementary Table 3).
Demographics and clinical characteristics of the sample
Circulating progenitor cells in mild cognitive impairment
Relative to those identified as CN, participants meeting neuropsychological criteria for MCI exhibited attenuated circulating progenitor cells, including cells expressing CD34+, F (1,27) = 6.053, p = 0.02, η2 = 0.18, CD34+CD133+, F (1,27) = 7.007, p = 0.01, η2 = 0.21, and CD34+CD133+CD309+,F (1,27) = 6.992, p = 0.01, η2 = 0.21, with medium-to-large effect sizes after correcting for age, sex, and education (Fig. 1). The same pattern of findings was replicated in univariate post-hoc analyses using an age-, sex-, and education-matched subset of participants (n = 10 CN, n = 10 MCI), although CD34+CD133+CD309+ was no longer significant (p = 0.056) in this analysis (Supplementary Table 4).

Progenitor cell counts are attenuated in MCI relative to normal aging. Circulating progenitor cell levels were attenuated in older adults with MCI relative to those who were CN, *p < 0.05.
Circulating progenitor cells and memory
After correcting for age, sex, and education, analysis of progenitor cells levels in relation to memory function across the entire sample (CN and MCI combined) indicated that participants with greater CD34+ cell levels exhibited better immediate verbal memory (p = 0.04) and delayed verbal memory (p = 0.02), as well as immediate visual memory (p < 0.001) (Table 2A). Participants with greater circulating levels of CD34+CD133+ cells also exhibited better immediate visual memory, ΔR2 = 0.18, β= 0.43, p = 0.001. Finally, participants with higher levels of CD34+CD133+CD309+ cells showed better delayed verbal memory, ΔR2 = 0.14, β= 0.39, p = 0.009.
CD34+ cell count correlates with memory, brain volume, and perfusion
Circulating progenitor cells and MRI measures of cortical thickness and white matter lesions
In the subsample that underwent structural MRI (n = 30), ROI analyses indicated that higher circulating CD34+ cell counts were correlated with greater cortical thickness in the left posterior cingulate gyrus, after controlling for age, sex, and total intracranial volume (p = 0.006), but there was no significant association with right posterior cingulate volume or bilateral hippocampal volume (Table 2B). Parallel VBM analyses were not significant after family-wise error correction, but analyses using the uncorrected p < 0.001 cutoff confirmed that higher CD34+ cell counts correlated with greater cortical thickness within a significant cluster of left posterior brain regions, including the posterior cingulate, precuneus and lingual gyrus (Fig. 2; top row in Table 3; Supplementary Figure 1).
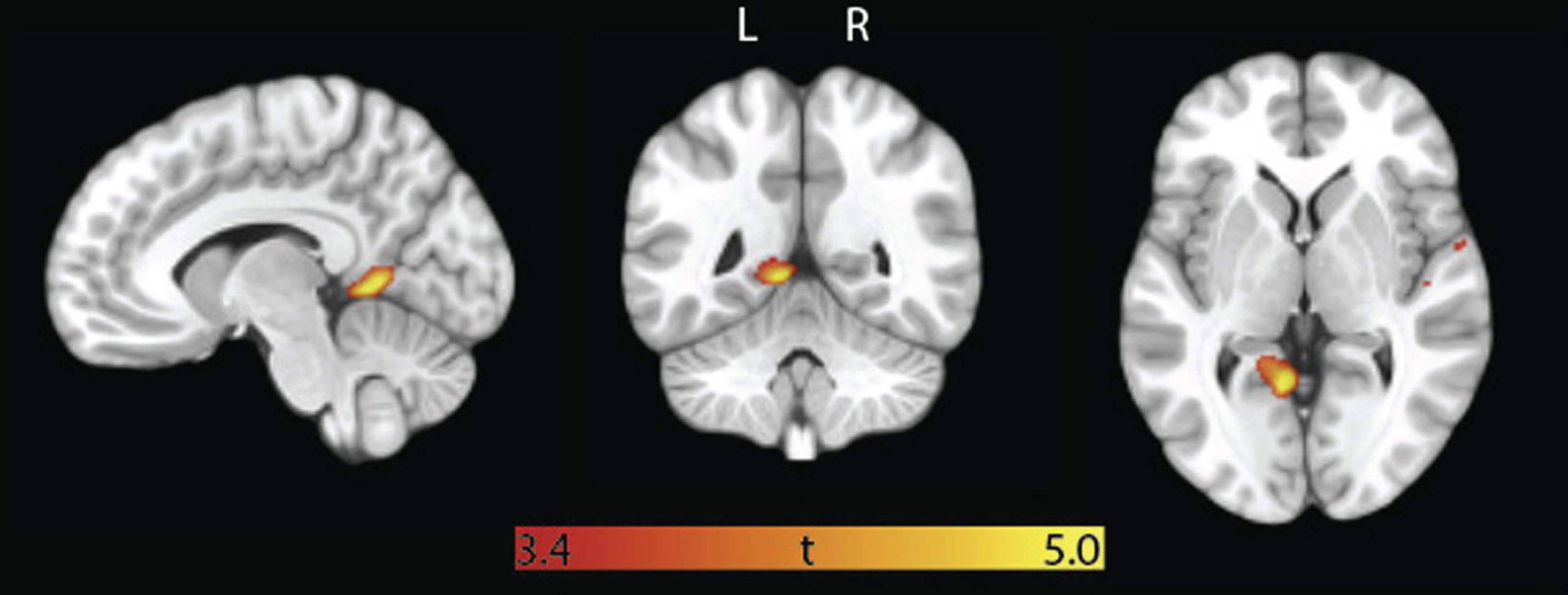
VBM analysis: CD34+ cell counts correlate with grey matter volume in posterior regions. Overlaid t-map is the result of voxel-wise multiple regression with CD34+ cell count as independent variable, grey matter volume as dependent variable, and age, sex, and total intracranial volume as covariates of no interest. Map is thresholded at cluster-level uncorrected p < 0.001. Minimum and maximum t-scores are displayed on the color bar.
CD34+ significant clusters in VBM analysis
Other progenitor cell counts (CD34+CD133+ and CD34+CD133+CD309+) did not show significant correlations with brain volumes in ROI analyses (all ps > 0.10). There was also no significant relationship between progenitor cell counts and WML volume (CD34+: ΔR2 = <0.001, β= 0.02, p = 0.90; CD34+CD133+: ΔR2 = 0.001, β= –0.03, p = 0.85; CD34+CD133+CD309+: ΔR2 = 0.01, β= –0.13, p = 0.45).
Circulating progenitor cells and cerebral perfusion
In the subsample that underwent ASL-MRI (n = 27), participants with lower CD34+ cell counts exhibited greater bilateral hippocampal perfusion, after controlling for age and sex, both ps < 0.05 (Table 2C). There was no significant relationship with posterior cingulate perfusion, and other progenitor cell counts (CD34+CD133+ and CD34+CD133+CD309+) did not show significant correlations with brain perfusion (all ps > 0.10).
DISCUSSION
The present study findings indicate that reduction of circulating progenitor cells in older adults is associated with worse memory function, diagnosis of MCI, thinning of posterior cortical regions and hyperperfusion of the bilateral hippocampus, all of which are indicators of pathological aging linked to increased risk of AD dementia. These findings are consistent with prior studies showing reduced CD34+ cells in dementia patients relative to normally aging controls [14], but further suggest that CD34+ cell loss may be related to early risk indicators rather than the mere product of advanced neurodegeneration. Although causality cannot be concluded from this observational study, the findings provide convergent support for an association between progenitor cell levels and multiple dementia risk factors. We hypothesize that progenitor cells may play a potentially protective role in aging brain health and could represent valuable therapeutic targets in prevention trials, but much more research is needed todetermine whether the observed associations in the present study are causal in nature. The current study evaluated only cross-sectional associations. Assessment of progenitor cell levels in relation to longitudinal decline in cognition and brain health will allow examination of the potential for progenitor cell levels as an early risk indicator. Future studies will employ logistic regression and other predictive modeling methods to determine whether progenitor cell counts may be of value in identifying those at risk for progressive cognitive decline and dementia.
Our MCI sample was significantly older than the CN group, but there was no significant relationship between age and progenitor cell levels, and post-hoc analysis of an age-matched sample revealed a similar pattern of findings, suggesting that demographic factors alone cannot account for the observed attenuation of progenitor cells in MCI. Prior studies have shown that CD34+ cells correlate with performance on global cognitive screening measures among dementia patients [13, 27]. The present study extends these findings to non-demented older adults with low vascular risk, where progenitor cell levels correlated with neuropsychological tests of memory across the entire sample. Insidious memory decline is characteristic of the earliest stages of AD, and may be detectable even in older adults with normal range cognitive function [28]. The observed relationship between progenitor cell levels and memory function raises the possibility that progenitor depletion may contribute to the earliest stages of cognitive decline in patients at risk for AD dementia, although the relevance of progenitor cells to AD cannot be inferred from the present study of non-demented older adults.
Prior studies have been mixed with regard to the role of circulating progenitor cells in AD versus vascular cognitive impairment [13, 27]. The present study did not include assessment of AD biomarkers, limiting conclusions regarding etiological factors, but the study sample was deliberately selected to be low in vascular risk burden. No participants reported a known history of cardiovascular disease or stroke and the level of WML burden was correspondingly low. We also noted that circulating progenitor cell levels were not associated with vascular risk factors or WML volume, and vascular risk burden did notdiffer between CN and MCI participants. However, we note that the limited sample size does not allow us to rule out significant associations between vascular risk and progenitor cell levels in the general population. Worse memory performance, diagnosis of MCI, and thinning of posterior cingulate cortex are all characteristic of the early stages of AD [29–32]. This is consistent with prior studies indicating that CD34+ cells are depleted in dementia due to AD, where they correlate with global impairment and CSF amyloid-β levels [13, 14].
Animal studies have demonstrated that circulating progenitor cells may be derived from the bone marrow [33], but can also mobilize to the brain and differentiate into neural tissues [11, 34] or promote neurogenesis indirectly through paracrine growth factor signaling [10, 35]. Alternatively, these cells may help clear excess amyloid beta by differentiating into macrophages, microglia, or astrocytes [11, 36–38]. The subset of CD34+ cells with proangiogenic capability, including CD133+ and CD309+ cells, may also promote neurovascular unit function [39], which is compromised early in the AD process [40, 41]. The present study found that although CD34+CD133+ and CD34+CD133+CD309+ cells were attenuated in MCI and showed some correlation with memory function, they did not relate to brain volume, perfusion or WML burden. This could be due to the nature of our low vascular risk sample, as these markers may have greater relevance to vascular disease.
Prior studies have indicated that circulating CD34+ cell levels are associated with greater cerebral perfusion in stroke patients [16], suggesting that improved cerebral perfusion may be one mechanism by which CD34+ cells could benefit cognitive function. The present study found that older adults with lower CD34+ cells actually exhibited greater hippocampal perfusion. This finding is consistent with several prior studies demonstrating hippocampal hyperperfusion in CN older adults at genetic risk for AD [42], those with subtle memory decline [43] and in MCI [44]. Based on these studies, a biphasic model has been hypothesized in which early compensatory changes in neurovascular function lead to an initial hyperperfusion of the hippocampus followed by an eventual decline in perfusion after the onset of AD dementia [45]. Thus, the inverse correlation between CD34+ cell levels and hippocampal perfusion is consistent with the notion that lower CD34+ cell counts are associated with AD risk factors. Postmortem studies in human bone marrow transplant patients demonstrate that progenitor cells migrate to the hippocampus where they undergo transdifferentiation into neurons, astrocytes and microglia, specifically within hippocampal regions implicated in adult neurogenesis [11], although this mechanism is highly controversial. The hippocampus and posteriorcingulate are highly interconnected structures that play critical roles in memory function and are implicated in AD dementia [46]. Thus, the observed correlations suggest a relationship between CD34+ cell levels and both structural and functional markers in these regions, as well as memory performance. We hypothesize that CD34+ cells may be playing a supportive role in memory system function in older adults at risk of AD dementia.
It remains unclear whether progenitor cell depletion is a contributing cause or downstream consequence of neuropathology. In the context of chronic tissue inflammation, circulating progenitor cell levels may attenuate as these cells migrate to and are sequestered within sites of injury, where they may undergo terminal differentiation and lose their regenerative capacity [47]. However, progenitor cells may also be mobilized by tissue injury and other pathogenic factors [48]. For example, in a study relating CD34+ cell counts to CSF amyloid-β levels, AD patients had reduced CD34+ cells relative to controls, yet there was an inverse relationship between CD34+ cell counts and CSF amyloid-β [14], suggesting that higher CD34+ counts were related to greater cerebral retention of amyloid-β [49]. These complexities may partly explain the mixed findings regarding whether CD34+ cell counts are increased or decreased in more advanced AD dementia patients versus controls[13, 50–52].
The present study addresses some of these questions from the prior literature by investigating progenitor cell counts during the earlier, MCI stage of cognitive decline in a sample with low levels of vascular risk. The use of a community-based sample with actuarial criteria for MCI is another study strength relative to prior findings comparing memory clinic-based samples to normal volunteers. The present study is also the first to investigate progenitor cell correlations with neuropsychological measures of memory and MRI-based indicators of brain volume and perfusion. Findings are consistent with the progenitor reserve hypothesis, which states that circulating progenitor cells may help buffer cognitive decline in the context of neuropathology [53, 54]. Thus, it is possible that having lower levels of progenitor reserve increases the risk of dementia, but it is also possible that progenitor cells are depleted from circulation during the early stages of disease due to migration and sequestration at sites of neuronal and cerebrovascular injury. Further studies utilizing longitudinal design and in vivo markers of AD pathophysiology are needed to better establish how progenitor cells may relate to disease progression and clinical manifestation. Future studies in animal models may also examine how tissue migration and progenitor cell fate interact with disease processes to shed further light on potential mechanisms that could mediate a hypothesized protective benefit of progenitor cell reserve.
Prior animal studies have found that granulocyte macrophage colony stimulating factor (GM-CSF) and other factors that stimulate mobilization of progenitor cells are critically involved in learning, memory, and synapse formation and maintenance[4, 56]. Thus, pharmacological manipulation of circulating progenitor cells through GM-CSF analogues may have therapeutic potential. Direct progenitor cell transplantation has also been successfully applied to coronary artery disease in clinical trials [57], suggesting potential for multiple therapeutic modalities. Clinical trials in AD dementia patient samples using GM-CSF analogues are underway, but these trials may fail due to treatment initiation during a late stage of neurodegeneration after the emergence of frank dementia. The current study suggests that the focus of these trials shift to the MCI phase of disease when secondary prevention options may provide a greater benefit with regard to risk of progression to dementia.
The present study findings are observational in nature, but they support a relationship between greater progenitor cell reserve and decreased risk factors for dementia. It has been hypothesized that early intervention to increase circulating progenitor cell levels could reduce dementia risk by supporting cerebrovascular and neuronal function. Future studies further examining the potential protective impact of circulating progenitor cells are warranted. Additionally, further studies are needed to explore other specific progenitor subpopulations in an effort to identify specific cell-types and regulatory factors that may be relevant to cognitive decline.
The present study is not without limitations in that it was small and cross-sectional. We did not employ multiple comparison corrections due to the exploratory nature of the study, and we did not have sufficient power for our voxel-level findings to survive family-wise error correction. Age and APOE ɛ4 differences between cognitively normaland MCI groups may have confounded study findings, although matched comparisons showed a similar pattern of results. Despite these caveats, the study findings contribute substantially to our currently very limited understanding of the relationship between progenitor cells and both aging brain health and dementia risk factors.