
Abstract
Background:
The α7 subtype of the nicotinic acetylcholine receptor (nAChR) is considered important in higher cognitive functions, and cholinergic loss underpins the pathophysiology of Alzheimer’s disease (AD). However, the relationships between α7 nAChR function and clinical functions or amyloid-β (Aβ) deposition remain to be explored in the living AD brain.
Objective:
We aimed to elucidate the relationship between α7 nAChR availability in the specific cholinergic region and cognitive decline in the Aβ-confirmed AD brain.
Methods:
Twenty AD patients and ten age-matched healthy subjects were examined. The α7-nAChR availability and Aβ deposition were evaluated using positron emission tomography with an α7 nAChR radiotracer 11C-(R)-MeQAA and 11C-Pittsburg compound B (11C-PiB), respectively. Semi-quantified values of tracer binding were estimated with a simplified reference tissue method for BPND of 11C-(R)-MeQAA and a tissue ratio method for SUVR of 11C-PiB. These parameters and clinical scores were compared voxel-wise using a statistical parametric mapping method.
Results:
The levels of 11C-(R)-MeQAA BPND in the temporal and prefrontal cholinergic projection regions were significantly lower in AD, and negative correlations were found between 11C-PiB SUVR and 11C-(R)-MeQAA BPND in the region of the nucleus basalis magnocellularis and medial prefrontal cortex. Levels of 11C-(R)-MeQAA BPND were significantly correlated with memory and frontal function scores in AD.
Conclusion:
The association between Aβ burden and α7-nAChR reduction in the basal forebrain cholinergic system was highlighted in relation to cognitive decline in AD. This suggests that Aβ-linked α7-nAChR reduction is clinico-pathophyisologically important for considering a good therapeutic target in AD.
Keywords
INTRODUCTION
The nicotinic acetylcholine receptor (nAChR) system has a crucial role in modulating attention and enhancing cognitive performance [1, 2], and the α7 nAChR subtype is involved in many cognitive-behavioral actions [3]. This is supported by human studies in which α7 nAChR-mediated agonists act on working memory and attentional performance [4, 5], and a marked reduction in α7 nAChR availability is present in the postmortem Alzheimer’s disease (AD) brain [6]. Thus, measuring α7 nAChR availability in vivo may enable prediction of the level of cognitive decline at the molecular level. Dysfunction of the cholinergic system contributes to cognitive impairments in AD, in which amyloid-β protein (Aβ) is a pathological hallmark. Our recent positron emission tomography (PET) study [7] has revealed that in AD patients, 18F-2FA-85380 binding, a measure of the α4β2 subtype of nAChR, in the medial prefrontal cortex (mPFC) and nucleus basalis magnocellularis (NBM) correlates positively with frontal functions, whereas 11C-Pittsburgh compound B (11C-PiB) does not. This finding suggested that the dysfunction of the α4β2 nAChR system, rather than the deposition of Aβ in these regions, played an important role in cognitive dysfunction in AD, which cast one question about another important cholinergic component, α7-nAChR, for its contribution to the AD pathophysiology.
In the ex vivo experimental setting, the results of previous postmortem studies evaluating α7-nAChR in the AD brain are controversial: there are reports of increased, decreased, and unchanged levels in AD compared with controls [8–18]. However, the discrepancy appears to be a result of the regions of interest (ROI) and techniques used. For instance, autoradiography studies using 125I-α-bungarotoxin have reported unchanged binding in the cerebral cortex [7–10], and decreased [8] or stable [9] binding in the hippocampus. In contrast, the expression levels of α7 mRNA have shown to be stable in the temporal cortex [7] and increased in the hippocampus [8] and basal forebrain [12]. The protein levels of α7 are unchanged [13, 14] or decreased in the cerebral cortex [15, 16] and decreased in the hippocampus [14]. Immunohistochemical studies have consistently reported decreases in α7-immunopositive neurons and increases in α7-immunopositive astrocytes in the temporal cortex and hippocampus [9, 18]. Collectively, ex vivo findings suggest that the decreased number of neurons expressing α7-nAChR with a mixture of the increased glial expression of the receptor might result in decreased or unchanged number of binding sites in the cerebral cortex and that α7 mRNA is probably expressed highly in compensation in the basal forebrain.
We have recently developed a novel radiotracer for α7-nAChR, (R)-2-[11C]methylaminobenzoic acid 1-aza-bicycle[2.2.2]oct-3-yl ester (11C-(R)-MeQAA) [18]. The binding affinity, Ki, of 11C-(R)-MeQAA for α7-nAChR is 41 nM, and the specific binding can be displaced by methyllycaconitine, an antagonist of α7-nAChR, in the mouse brain, or by SSR190711, a partial agonist of the receptor, in the monkey brain [19]. Although a recently reported α7-nAChR tracer (18F-labeled ASEM) might have a better affinity than our 11C-(R)-MeQAA’s [20], the 11C-labeled tracer has a merit for combination of PET scans with different tracers within a day. The development of this radiotracer led us to investigate the relationship between α7-nAChR availability and Aβ deposition as well as cognitive dysfunction in patients with early-stage AD by measuring 11C-(R)-MeQAA binding and 11C-PiB uptake using PET simultaneously. Our initial hypotheses were as follows: 1) the availability of α7-nAChR is decreased in the brains of AD patients; 2) the decrease in α7-nAChR availability is correlated with Aβ deposition; and 3) the decreased availability of α7-nAChR in the cholinergic projection system, such as the NBM and mPFC, is associated with frontal cognitive dysfunction.
MATERIALS AND METHODS
Participants
We tested twenty drug-naïve never-smoking AD patients and ten age-matched non-smoking cognitively normal healthy subjects out of 20 healthy people (consisting of young to old non-smokers). We did not include any persons with smoking habit (in the past or present) or occasional smokers. The data of the young group were used preliminarily for an aging effect of this tracer (Supplementary Figure 1). The demographic features of the current 11C-(R)-MeQAA study are shown in Table 1. The diagnosis of AD was based on the criteria of the National Institute of Neurological and Communicative Disorders and Stroke-Alzheimer’s Disease and Related Disorders Association (NINCDS/ADRDA). All neuropsychological tests as shown in Table 1 were performed within a month of PET measurement. The healthy subjects had no family histories of psychiatric or neurological diseases and no abnormal findings by magnetic resonance imaging (MRI). No participants took any nootropic drugs, including acetylcholinesterase inhibitors. The Ethics Committee of Hamamatsu University approved the study, and written informed consent was obtained from all participants prior to enrollment.
Demographic features of participants
Data are expressed as mean (SD). *p < 0.05, unpaired t-test vs. elderly control for cross-sectional comparison. aPearson’s Chi-square test. bStudent t-test. cPoints of connections: Eight points represents a perfect score. CDR, Clinical Dementia Rating; FAB, Frontal Assessment Battery; MMSE, Mini-Mental State Examination; WMS-R LM, Logical Memory Subset of the Wechsler Memory Scale Revised; SDS, Zung Self-rating Depression Scale; n/a, not applicable.
MRI data acquisition
Prior to PET measurements, MRI was performed to determine the brain regions of concern for setting ROIs by using a 1.5 T MR scanner (Signa HDxt, GE, USA) with the following acquisition parameters: 3D mode sampling, TR/TE (25/Minimum), 30° flip angle, 1.5 mm slice thickness with no gap, and 256×256 matrices.
PET data acquisition
We used a new high-resolution PET camera (HITS-655K; Hamamatsu Photonics KK, Hamamatsu, Japan) [21] with LYSO scintillator arrays (1.2 mm) and 8×8 array MPPCs, yielding 83 slices simultaneously. The axial FOV is 202 mm, and its spatial resolution is 1.4 mm, with a sensitivity of 4% over. By use of a head cap with light emitting diode markers and location detectors, head motion was monitored during scans in a similar manner to using a facemask [22, 23]. The synthesis of 11C-(R)-MeQAA was based on the previous literature [19]. After intravenous injection of a dose of 5 MBq/kg 11C-(R)-MeQAA, dynamic scans were performed for 60 min (time frames:12×10 s, 18×60 s, and 20×300 s) because the tracer comes to equilibrium around 40 min postinjection. The amounts of 11C-(R)-MeQAA given, injected doses and specific radioactivity were 4.21±1.00 MBq/kg, 0.98±0.32 μg and 106.08±23.83 GBq/μmol in the AD group and 3.75±0.99, 1.12±0.39, 113.53±42.71 in the normal group, respectively.
For the amyloid measurement by PET, 70 min-duration scans (time frames: 12×10, 18×60, and 10×300 s) initiated immediately after an injection of 5 MBq/kg 11C-PiB without blood sampling were performed within a week before or after the 11C-(R)-MeQAA. The amounts of 11C-PiB given were 5.23±0.34 MBq/kg in the AD group and 5.13±0.19 in the normal group.
Evaluation of 11C-(R)-MeQAA binding potential and amyloid-β accumulation
The non-displaceable binding potential (BPND) of 11C-(R)-MeQAA was calculated on the basis of a simplified reference tissue model, in which the corpus callosum was selected as the reference region conceivably devoid of nAChR [20, 24]. However, the report that nAChR in this region would be affected by smoking [25] suggests that the current BPND of this tracer might be underestimated. The irregular ROIs were determined bilaterally in the following eleven brain regions: cerebellum, amygdala, NBM, caudate, putamen, thalamus, mPFC, frontal, temporal, parietal, and medial parietal cortices in MR images by using an MR image atlas [26]. This procedure was able to minimize the partial volume effect. The bilateral values were averaged for comparison because of no laterality in tracer binding (Supplementary Figure 2). In this procedure, the ROI of the NBM region was specifically determined in the area beneath the anterior commissure at the nucleus accumbens according to the recent report showing the human NBM on MRIs [27]. The ROIs placed in MRIs were automatically transferred to the corresponding parametric BPND images by using image-processing software (Dr View, Asahi Kasei Co., Tokyo, Japan) on a SUN workstation (UltraSPARC, SUN Microsystems, San Diego, CA, USA), as previously described [28, 29]. The parametric BPND images were analyzed by both ROI-based analyses and voxel-based analyses by using statistical parametric mapping (SPM8, Wellcome Department of Cognitive Neurology, London, UK).
Parametric images of 11C-PiB uptake in the brain were generated from PET data accumulated from 50 to 70 min postinjection with a unit of standardized uptake value relative to the cerebellar count (SUVR) [30, 31]. The 11C-PiB SUVR parametric images were used for further voxel-based analyses.
Statistical analysis
In the ROI-based analyses, comparisons between dependent variables were performed using Spearman’s rank correlation, and significance was defined as p < 0.0015 because of multiple comparisons (11 ROIs×3 variables). These analyses were conducted using SAS (version 9.0, SAS Institute Inc.). To examine the direct relationship between 11C-(R)-MeQAA binding and Aβ burden in the same regions, simple regression analyses were performed to compare 11C-(R)-MeQAA BPND with 11C-PiB SUVR in the same ROI within the AD group. Significance was defined as p < 0.05. In addition, to examine the effect of Aβ burden in the cholinergic origin (NBM) on the cholinergic projection regions, multiple regression analyses were conducted with a significance threshold of p < 0.0045, accounting for multiple comparisons.
In the voxel-based analyses, individual MRI data were first normalized to a standard anatomical orientation (MNI space) by using an MRI template in SPM8. Then, using generated normalization parameters, the 11C-(R)-MeQAA BPND parametric images of each individual were normalized to the MNI space. The normalized images were smoothed with an isotropic Gaussian kernel of 6 mm full width half maximum. Voxel-wise multiple regression analyses were performed to compare the 11C-(R)-MeQAA BPND levels with the clinical variables and the 11C-PiB SUVR. Using the factorial SPM model, one-way analysis-of-variance was performed to determine differences in 11C-(R)-MeQAA BPND levels between groups, with gender, age, and education treated as confounding covariates. The level of significance was set at p < 0.001, uncorrected for peak height, with a cluster size larger than 50, because the regions of focus were known a priori as areas with cholinergic innervation [32].
RESULTS
Decrease in 11C-(R)-MeQAA binding in AD
Compared with the normal group (Fig. 1A), area under the curves (AUCs) of tissue time activity curves in both the thalamus (F-test, AUC-AD versus AUC-normal: f = 17.6, p = 0.0014) and frontal cortex (f = 20.3, p < 0.0001) were reduced significantly in the AD group (Fig. 1B), whereas the curves of the corpus callosum were similar (f = 2.02, p = 0.162). In addition, in this corpus callosum, there was no difference in AUC-cc between young and elderly control groups (f = 4.34, p = 0.0516). The latter finding indicates that the corpus callosum might be adequate as the reference region for the α7-nAChR tracer, similarly to the α4β2 nAChR tracer, as previously reported [7, 33], while this theory may not be available among studies with some smokers or nicotine-treated subjects in view of previous literature [25]. ROI- and voxel-based between-group analyses showed a widespread decrease in the 11C-(R)-MeQAA BPND level over the NBM and NBM-origin cholinergic projection regions (prefrontal cortex, medial and lateral temporal cortices, parietal cortex) (Fig. 1C, D and Table 2). In the preliminary study, use of SUVR-cc might be better to differentiate AD from healthy elderly than use of SUVR-pons (Supplementary Figure 3). Still, the statistical threshold used in the current result with BPND was more robust than the threshold of these indices.

Decrease in 11C-(R)-MeQAA binding in AD patients. Decreased 11C-(R)-MeQAA uptake was observed in the time activity curves of the receptor-rich (thalamus) and receptor-sparse (frontal) regions despite a lack of differences in the reference region (corpus callosum) (A: normal, B: AD). T-test showed an extensive decrease in 11C-(R)-MeQAA binding in almost all brain regions except for the cerebellum (C, *p < 0.05). SPM analysis confirmed these regions of interest-based findings (D). BPND of 11C-(R)-MeQAA was determined using a pseudo-reference region.
Statistical parametric mapping results for 11C-(R)-MeQAA binding reduced in AD patients compared with healthy controls
Statistical significance was assumed at an individual voxel level of p < 0.001, uncorrected, k > 100 voxels. †Talairach brain atlas (x, y, z). BA, Brodmann’s area; L/R, left or right side of the brain; NBM, nucleus basalis magnocellularis.
Regional 11C-(R)-MeQAA binding and cognitive functions in AD
In the AD group, the Spearman rank correlation test showed a significant correlation between the FAB score and 11C-(R)-MeQAA BPND levels in the NBM (ρ= 0.655, p = 0.0017) and a weak correlation between FAB score and 11C-(R)-MeQAA BPND levels in the mPFC region (ρ= 0.375, p = 0.130), but no significance regarding the thalamus (ρ= 0.204, p = 0.341) (Fig. 2A). SPM correlation analysis supported this finding (Fig. 2B, Table 3). No other clinical variables showed a significant correlation with regional 11C-(R)-MeQAA BPND in any regions.

Correlations between 11C-(R)-MeQAA binding and clinical parameters in AD patients. Spearman’s rank correlation analysis showed a significant positive correlation between FAB scores and 11C-(R)-MeQAA BPND values in the NBM (*p < 0.05 corrected for multiple comparisons) (A). SPM correlation analysis results were consistent with this finding (B). BPND of 11C-(R)-MeQAA was determined using a pseudo-reference region.
Statistical parametric mapping results on correlations between 11C-(R)-MeQAA binding and FAB scores in AD patients
Statistical significance was assumed at an individual voxel level of p < 0.001, uncorrected, k > 100 voxels. †Talairach brain atlas (x, y, z). BA: Brodmann’s area; L/R: left or right side of the brain; FAB: frontal assessment battery; NBM: nucleus basalis magnocellularis.
Aβ deposition and decreased 11C-(R)-MeQAA binding in the AD brain
Consistent with a previous report [34], as seen in Fig. 3A, SPM analysis showed significant elevation of 11C-PiB SUVR (red) globally in the AD brain with prefrontal and precuneus dominance and partly overlapping regions (purple) with reduction in 11C-(R)-MeQAA binding (blue). ROI-based analyses showed a significant negative correlation between 11C-PiB SUVR and 11C-(R)-MeQAA BPND in the NBM (r = 0.618, p = 0.00214) and a tendency toward negative correlations in the mPFC, medial parietal and lateral parietal cortices (Fig. 3B).
Further regression analyses on the effect of the Aβ burden in the NBM (Fig. 4A) on the cholinergic terminal regions showed that the NBM 11C-PiB SUVR was negatively correlated with the 11C-(R)-MeQAA BPND level in the mPFC (r = 0.676, p = 0.0011) and the medial parietal cortex (r = 0.691, p = 0.0007), whereas no significant correlation was found in the thalamus (Fig. 4B). The SPM finding was compatible with these ROI-based results (Fig. 4C).
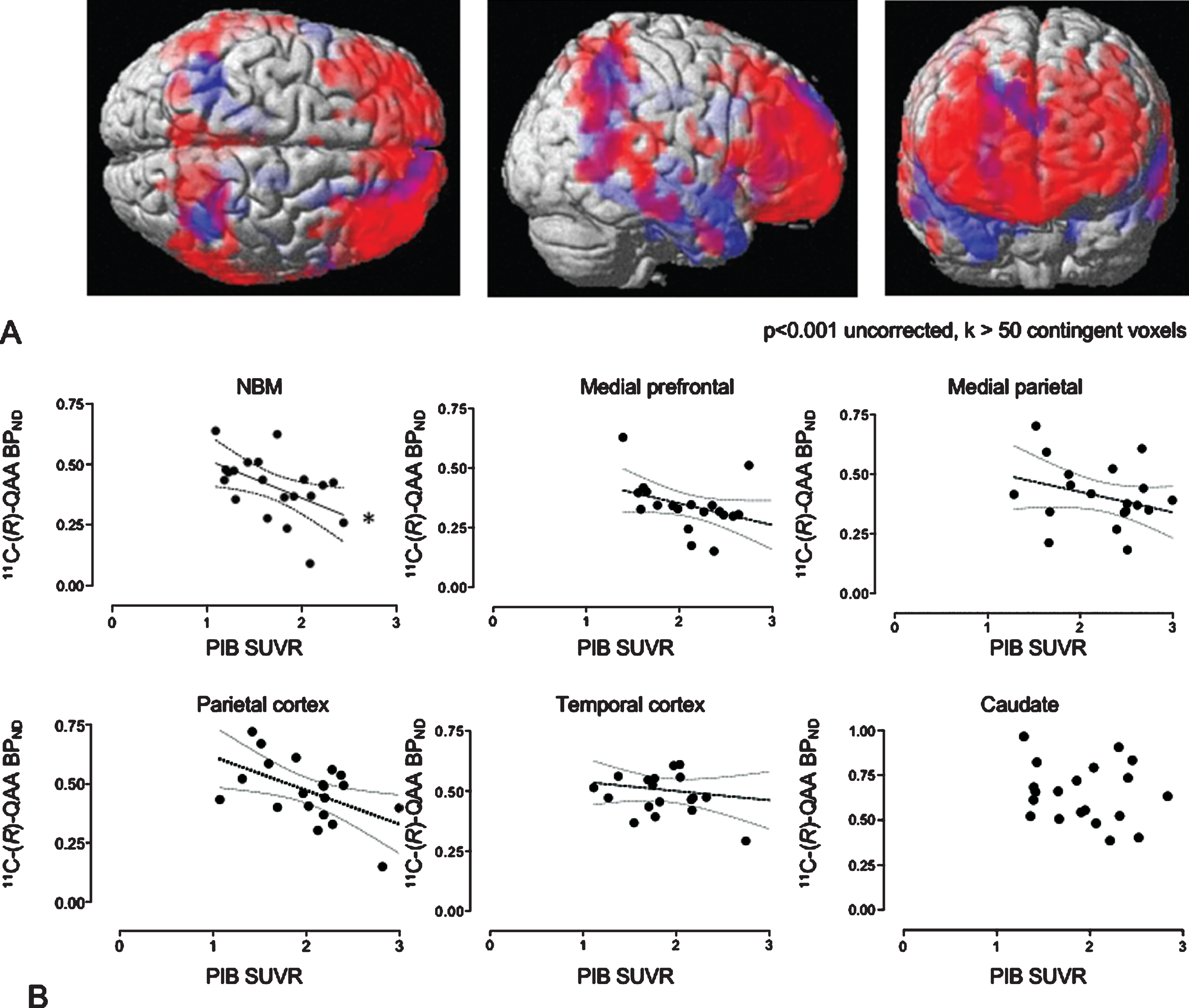
Correlations between 11C-(R)-MeQAA binding and 11C-PiB uptake within the same regions. (A) SPM rendering images show a significant increase in 11C-PiB uptake (red) and significant decrease in 11C-(R)-MeQAA binding (blue). The purple is for overlap. There is a significant correlation between 11C-(R)-MeQAA binding and 11C-PiB uptake in the NBM (*p < 0.05, corrected for multiple comparisons). Dotted lines represent 95% confidence intervals of the fitted lines in the scattergram (B). BPND of 11C-(R)-MeQAA was determined using a pseudo-reference region.

Correlations between regional 11C-(R)-MeQAA binding and NBM 11C-PiB accumulation. MRI/PET fusion images indicate a significant 11C-PiB increase (red) and 11C-(R)-MeQAA decrease (green) (A). Correlations of 11C-PiB uptake in the NBM with 11C-(R)-MeQAA binding in other brain regions show significant negative reductions in the MPF and medial parietal cortices (*p < 0.05 corrected). Dotted lines represent 95% confidence intervals of the fitted lines in the scattergrams (B). SPM correlation analysis was consistent with this finding (C). BPND of 11C-(R)-MeQAA was determined using a pseudo-reference region.
DISCUSSION
The current study showed that the levels of 11C-(R)-MeQAA BPND were significantly lower in the cortical and subcortical brain regions in AD patients compared to healthy controls. Decreased 11C-(R)-MeQAA binding in the NBM, but not in the mPFC, significantly correlated with cognitive decline as assessed by FAB in AD patients. In AD, increased levels of 11C-PiB SUVR in the NBM were associated with decreases in 11C-(R)-MeQAA binding in the NBM and mPFC.
To the best of our knowledge, this report is the first to illustrate the availability of α7-nAChR in the living brains of Aβ-confirmed AD patients. The availability of α7-nAChR decreased with age (Supplementary Figure 1), similarly to the results from a different α7-nAChR tracer 18F-ASEM [35] and the postmortem data from the healthy cortical samples [36]. As described earlier, the findings from postmortem studies suggest that the decreased number of neurons expressing α7-nAChR and the increased glial expression of the receptor result in a stable or decreased total number of binding sites and protein levels of α7-nAChR in the cerebral cortex. The decreased levels of 11C-(R)-MeQAA BPND in the cholinergic projection regions in the living AD brain found in the current study where age was used as a confounding covariate support the postmortem findings.
Although there was not a significant correlation between the levels of 11C-(R)-MeQAA BPND and 11C-PiB SUVR in the same ROI, except for the NBM, the 11C-(R)-MeQAA BPND levels in the mPFC and medial parietal regions were negatively correlated with the levels of 11C-PiB SUVR in the NBM in the AD group. This result indicates that the load of Aβ outside the NBM might not be directly related to altering the availability of α7-nAChR in the same loci of the brain. In contrast, the Aβ burden in the NBM might be more associated with the decrease in α7-nAChR availability in regions receiving cholinergic projections from the NBM. The positive correlation of the FAB score with the α7-nAChR availability but not with the 11C-PiB accumulation in these cholinergic projection regions further supports the idea that cognitive decline is attributable to a decrease in α7-nAChR availability in the NBM in AD. The precuneus is the most vulnerable area for 11C-PiB deposition, but the amount does not parallel cognitive deterioration in AD [37], thus indicating that the pathophysiological implication in cognitive decline is not the amount of Aβ accumulation but the area where Aβ accumulates. Indeed, in line with previous suggestions, even a small amount of Aβ deposition would be detrimental in the cholinergic origin NBM [37], as shown in this study. This hypothesis would further suggest that the cholinergic α7-nAChR system may be a key factor in determining future symptoms in cognitively normal elderly people with a larger amount of 11C-PiB in their brains [38, 39].
It has been reported that Aβ has a high binding affinity to α7-nAChR. Whereas Aβ at micromolar concentrations does not bind to 5-HT3, NMDA, or muscarinic receptors, Aβ binds to α7-nAChR with exceptionally high affinity, approximately 5,000-fold higher than with α4β2-nAChR [40]. These results indicate that an exceptionally high-affinity interaction between Aβ and α7-nAChR may serve as a precipitating factor in the formation of amyloid plaques and thereby contribute to the selective degeneration of the cholinergic neurons that originate in the NBM. A previous histochemical study has shown that neurons containing a high amount of intracellular Aβ express relatively high levels of α7-nAChR in the AD brain and that the rate of Aβ internalization is selectively blocked by α-bungarotoxin [41]. These lines of evidence support our results, indicating that Aβ deposition in the NBM cholinergic system might be more important than amyloid pathology in other regions to the deterioration of cognitive function in AD.
Consistently with previous results [7], the 11C-(R)-MeQAA binding (α7-nAChR availability) was associated with only the FAB score among neuropsychological tests in AD patients. These findings suggest that the FAB is sensitive to the assessment of attention-required cognitive function, as is believed worldwide [42], because the common component for cognitive valence between the α4β2- and α7-nAChRs is considered to be attention rather than memory and learning ability. However, this finding cannot exclude a clinical phenotype contribution of α7-nAChR to the regulation of memory and learning because a new α7-nAChR agonist agent, TC-5619, shows potential benefits of improved memory and executive functions in patients with schizophrenia [43]. Indeed, the long-term administration of galantamine, an acetylcholinesterase inhibitor that also acts as an allosteric modulator at the α4β2- and the α7-nAChRs, has been found to improve task speed and verbal memory in schizophrenic patients [44]. Thus, a more sensitive psychological test specifically for memory function such as subdomains of WMS-R would show any significant correlation with α7-nAChR binding in this study.
Limitations
The estimation of binding potential of 11C-(R)-MeQAA might be better improved with a gold-standard arterial sampling protocol, but it is actually difficult to apply it to vulnerable participants such as dementia patients. Still, the current estimation of BPND was shown to be a better index compared to more clinically-friendly SUVR method as shown in Supplementary Figure 3. Because animal studies showed a possibility of this tracer 11C-(R)-MeQAA binding to the 5-HT3 receptor in monkeys and a reportedly low specificity in mice, a more specific tracer might be preferable in humans albeit unknown species difference. However, the evidence that a considerable amount of α7-nAChR was blocked by its antagonist and that few reports about the 5-HT3 receptor exist in the AD research allows at this moment to proceed this type of in vivo pathophysiological study. Although the current study reported the correlations between Aβ deposition and α7 nAChR dysfunction, this finding could not disclose the cause-effect problem. Further studies with medical intervention or preclinical evaluation are needed to answer for the core mechanism for dementia progression in AD.
Conclusion
The current results confirm that an alteration in α7-nAChR is important in AD pathophysiology, in which Aβ deposition-linked α7-nAChR dysfunction specifically in the NBM may be one cause for cognitive decline in AD.
Footnotes
ACKNOWLEDGMENTS
The authors would like to thank Dr. M. Ogawa (Hamamatsu University School of Medicine), Messrs. T. Yamashita, M. Watanabe, H. Okada, I. Ando, M. Futatsubashi, T. Kosugi, K. Sato (Hamamatsu Photonics KK), and Mmes. Iori Suzuki and T. Shinke (Hamamatsu Photonics KK), and T. Kanno (Hamamatsu Photonics Medical Foundation) for their dedicated support. This work was supported by the grants from the New Energy and Industrial Technology Development Organization (NEDO), the Japanese Ministry of Education, Culture, Sports, Science and Technology, the Takeda Science Foundation, and Smoking Research Foundation.