
Abstract
We tested the potential of task-based functional neuroimaging as a biomarker of emerging prefrontal brain changes in progranulin (GRN) mutations carriers. Five GRN mutation carriers free of frontotemporal dementia (FTD) and 11 non-carriers from families with FTD-GRN underwent functional MRI while solving matrix-reasoning problems. Mutation carriers displayed slower responses for more difficult problems and lower lateral prefrontal activation across all problems. Overall task-evoked posterior ventrolateral prefrontal activation predicted mutation status with 100% sensitivity and 91% specificity. Volumetric differences did not account for activation differences. Prefrontal activation may have utility as a biomarker in GRN mutation.
Keywords
INTRODUCTION
Functional magnetic resonance imaging (fMRI) has been proposed as a biomarker for the earliest stages of neurodegenerative diseases [1]. Task-based paradigms may capture the functional impact of pathophysiological processes more directly than resting-state fMRI [2] because they allow tight control over the type of processing individuals engage in and provide concurrent behavioral evidence. We used task-based fMRI to investigate brain activation abnormalities due to mutations in the progranulin (GRN) gene prior to the onset of GRN-related frontotemporal dementia (FTD-GRN).
GRN mutations are a major cause of familial FTD. Progranulin is a secreted growth factor involved in numerous neuronal processes, including survival, cell cycle progression, proliferation, and migration as well as inflammation and wound-healing processes. All GRN mutations identified to date lead to a 50% loss in progranulin levels through lifelong haploinsufficiency [3]. While the average age of expected disease onset is 65 [4], this loss-of-function mechanism of GRN mutation indicates that the frontotemporal lobar degeneration associated with FTD-GRN likely unfolds over years [3]. Neuroimaging studies have reported early connectivity, metabolic, and structural brain abnormalities in GRN mutation carrier groups with mean age ranging from 40 to greater than 50 years. Together with parietal and temporal cortices, prefrontal cortex (PFC) is one of the most impacted brain regions. Reduced glucose metabolism has been found in ventrolateral and dorsolateral PFC regions [5, 6]. Altered functional connectivity has been observed within PFC and between PFC and posterior regions [7–9]. Gray matter volumetric changes have also been detected in PFC regions [10]. In the current task-based fMRI study, we employed a relational reasoning task [11] to test for lateral prefrontal functional abnormalities to advance the characterization of predementia FTD-GRN. As defined in our previous work, the term predementia describes mutation carriers that fall short of meeting FTD diagnostic criteria but may have some symptoms [6].
MATERIALS AND METHODS
Sixteen participants at risk for FTD-GRN due to an affected first degree relative were recruited through the University of British Columbia (UBC) Clinic for Alzheimer Disease and Related Disorders. Two additional subjects were excluded from analysis because of missing or erroneous data. The study was approved by the UBC Ethics Board. Written informed consent was obtained from each participant. All participants received cognitive, behavioral, and motor assessments at the time of imaging (see Table 1). A diagnostic classification of participants as unaffected (no symptoms identified) or clinically symptomatic no dementia (CSND) (symptoms suggestive of FTD identified but having no impact on everyday function) was assigned in a hierarchical manner [6]. A neurologist and neuropsychologist independently reviewed clinical assessments for each participant, and made a preliminary diagnosis based on their impressions. A consensus conference was then held to make a final diagnosis. This diagnosis consisted of the most clinically symptomatic classification from either neurologist or neuropsychologist. No participant met full criteria for FTD [12]. Genetic status was determined by GRN sequencing on blood samples at the Mayo Clinic, Jacksonville, Florida [13] and was unknown to the clinicians at the time of assessment and to the image analyst until the final stages of analyses.
Participant demographic and clinical data
*Age at scan – mean age at disease onset in family (negative numbers denote not having reached expected onset). **15-item version of Boston Naming Test. 3 MS, modified Mini-Mental State Examination; FAB, Frontal Assessment Battery; NPI, Neuropsychiatric Inventory; FRS, Functional Rating Scale; UPDRS, Unified Parkinson’s Disease Rating Scale. ***Demographic data for individual mutation carriers are not provided to maintain blinding.
Participants completed a relational reasoning task during fMRI scanning [11]. Problems were 3×3 matrices of low (0-relational; Fig. 1A), medium (1-relational; Fig. 1B), or high (2-relational; Fig. 1C) complexity. One image was missing and participants indicated which option best completed the matrix. After practice, participants completed four fMRI runs, each lasting 9.25 min and containing 18 slow event-related trials (Fig. 1D). Imaging on a 3T Philips scanner included functional T2-weighted gradient-echo EPI (TR 2000 ms, TE 30 ms, flip angle 90°, FOV 240×240 mm2, 80×80 matrix, 3×3 mm in-plane resolution, 36 slices, thickness 3 mm, 1-mm gap, whole-brain coverage), and a 3D T1-weighted anatomical scan (TR 7.6 ms, TE 3.6 ms, 256×200 matrix, 170 transverse slices, 1×1×1 mm resolution).
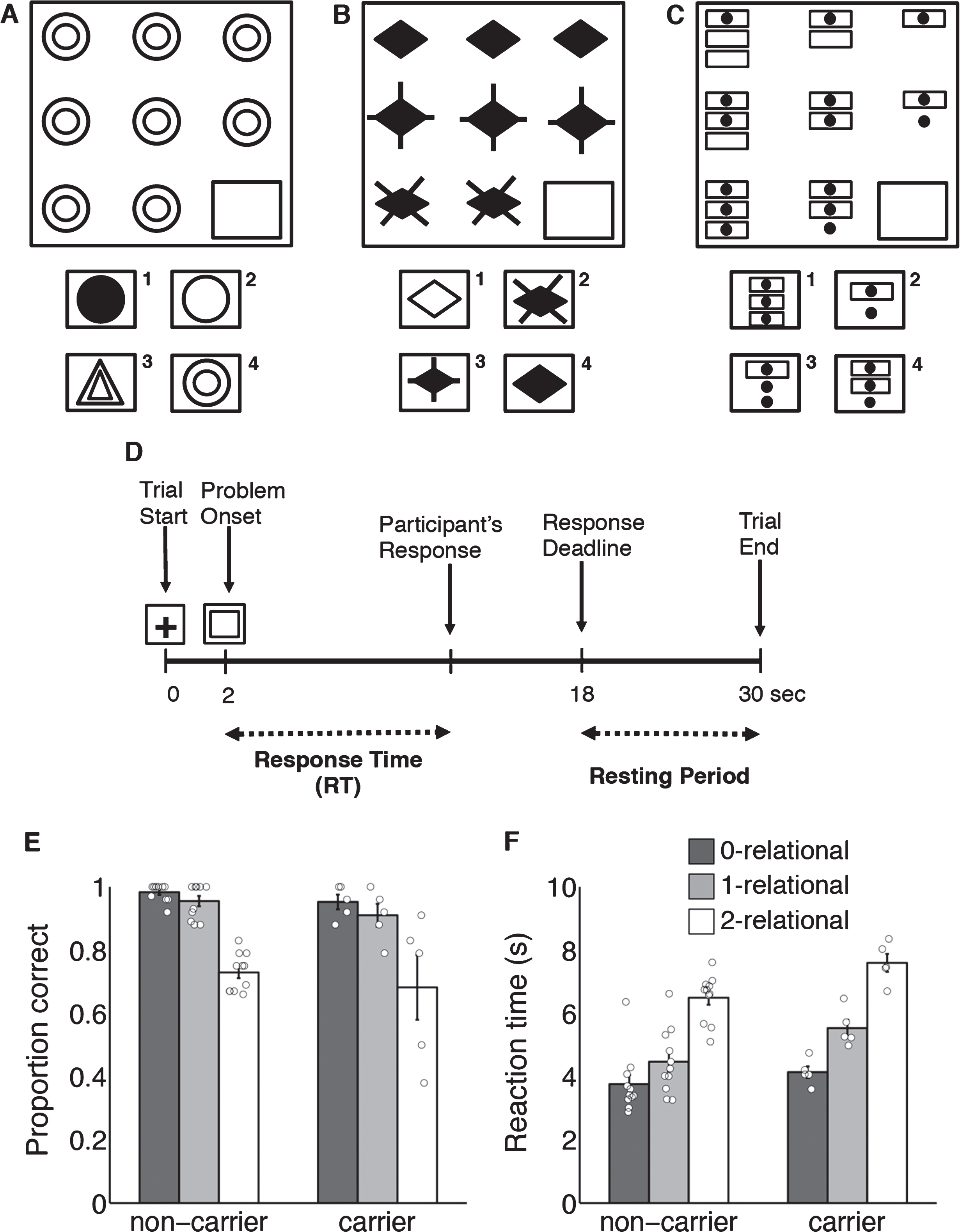
Behavioral task and behavioral results. A-C) Example of Raven’s Progressive Matrices, adopted from Christoff et al. [11]. Participants had to choose which object from the bottom completes the pattern. A) Zero-relational problem. B) One-relational problem. C) Two-relational problem. D) Timing of events within each trial. E) Proportion correct for each relational condition. F) Correct reaction time for each relational condition. Bar height represents group average, error bars represent standard error of the mean, open circles represent values for individual participants.
Image analysis was performed using SPM12 (Wellcome Department of Imaging Neuroscience, University College London). Preprocessing included realignment, slice timing correction, co-registration of the anatomical with the mean functional image, normalization to MNI space, spatial smoothing (8 mm), and high-pass filtering (128 s). Voxel-wise analysis using general linear model (GLM) was limited to correct trials. Trials were modeled as a box-car of reaction time (RT) length, convolved with canonical hemodynamic response function [9]. Three regressors of interest represented the correct 0-relational, 1-relational, and 2-relational trials. Incorrect trials were modeled separately and not considered further. Models included temporal derivatives.
To determine the effect of GRN-mutation, task-related activation was assessed in a set of regions that have been previously implicated in relational reasoning [11] and are typically affected in clinical FTD. Regions of interest (ROIs) were delineated using Harvard-Oxford atlas and included bilateral caudate, frontal pole, dorsolateral PFC (DLPFC), anterior ventrolateral PFC (aVLPFC), and posterior ventrolateral PFC (pVLPFC). GLM parameter estimates for each condition and participant were extracted from ROIs and submitted to repeated measures ANOVA (RM-ANOVA) with ROI and relational complexity level as within-subject factors, and carrier status as a between-subjects factor. Group comparisons within each ROI were performed using Wilcoxon rank sum test, appropriate for smaller sample size, with alpha = 0.01 to correct for multiple comparisons. For non-significant comparisons, p values exceeded by the statistical test/s were reported.
Reliable differences at the level of group means do not guarantee reliable diagnosticity for individual participants. To further probe the potential of task-based fMRI in detecting preclinical changes associated with a genetic mutation, we computed the probability of correct mutation status diagnosis based on task activation using a logistic regression classifier. A leave-one-out cross-validation procedure was used to estimate generalizability beyond the current sample, iteratively training a logistic regression classifier on 15 subjects and predicting the carrier status of the remaining subject. The prediction accuracy was tested against chance using a permutation test with 10000 Monte Carlo simulations.
To determine whether functional activation differences are driven by structural differences between groups, we performed a control volumetric analysis of the five ROIs selected for functional analysis including caudate, frontal pole, dorsolateral and ventrolateral PFC (anterior and posterior). Cortical reconstruction and volumetric segmentation was performed using Freesurfer (http://surfer.nmr.mgh.harvard.edu/). Three structural volumetric indices were extracted for each subject and ROI: raw volume in mm3, volume normalized by intracranial cavity volume to adjust for brain size, and cortical thickness (for cortical ROIs, excluding caudate ROI). The volumetric data were then compared between groups using the same procedures as for functional activation.
RESULTS
Genetic sequencing revealed that participants included 5 mutation carriers (4 females) and 11 non-carriers (6 females). Mutation carriers and non-carriers had comparable age and education (Table). No group differences were found for mean clinical scores (all p > 0.20). Among carriers, average years to estimated onset (subject age minus mean age at FTD onset in the family [14] was –5, that is, 5 years younger than mean onset in family, with a range of –16 to 5. Three mutation carriers were deemed CSND but only one had an abnormally elevated NPI score. All other scores for mutation carriers (cognitive, behavioral, and motor) were normal. Two non-carriers were also diagnosed as CSND. No participant met criteria for FTD. Carriers had similar matrix reasoning accuracy as non-carriers (Fig. 1E, all p > 0.25) but were slower on more complex reasoning problems (2-relational p = 0.03, 1-relational p = 0.07; 1F).
RM-ANOVA on parameter estimates (Fig. 2A) showed a main effect of region (p < 0.001), mutation status (smaller carrier than non-carrier activation: carriers mean = 0.42, SE = 0.128, non-carrier mean = 0.83, SE = 0.89, p = 0.02), and an ROI*mutation status interaction (p = 0.02). There was no main effect of relational complexity nor an interaction of relational complexity with carrier status (both F < 1.6, p > 0.2). Following up on the ROI*mutation status interaction, group comparisons within each ROI revealed that differences associated with mutation status were significant in aVLPFC (p = 0.006) and pVLPFC (p < 0.001). In pVLPFC, the region that showed the most reliable mutation status effect, we further tested the degree to which task-based activation (averaged across relational complexity level) may differentiate between mutation carriers and non-carriers on individual participant level. A logistic regression classifier separated perfectly between carriers and non-carriers based on overall task-related activation (Fig. 2B: all gray circles are above all white circles). Cross-validation showed an estimated out-of-sample generalization of classification accuracy of 94% (p = 0.0003 compared to chance; permutation test), with only one non-carrier misclassified, yielding 100% sensitivity and 91% specificity.
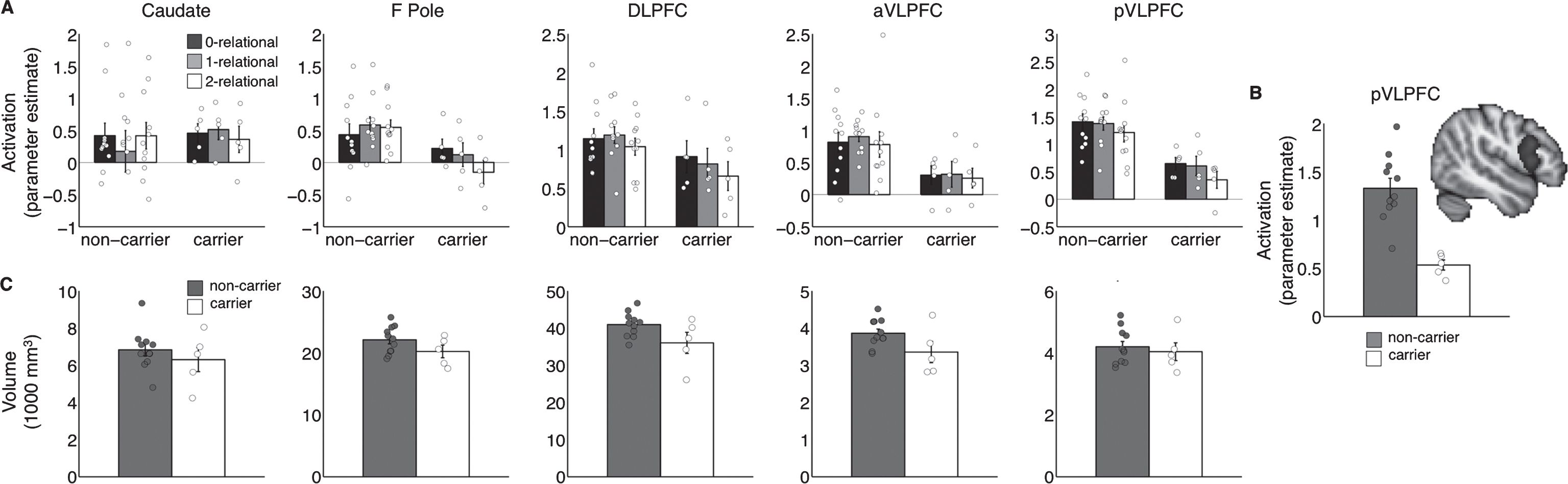
Group differences in ROI activation and volumes. Bar heights denote group means, circles denote individual participants. X-position of the circles is jittered to improve visualization of near-overlapping values. A) ROI activation for each relational difficulty. F Pole, frontal pole; DLPFC, dorsolateral prefrontal cortex; aVLPFC, anterior VLPFC; pVLPFC, posterior VLPFC. B) Task-related activation (averaged across conditions) in posterior VLPFC (depicted) for individual subjects who were mutation carriers (white circles) and non-carriers (gray circles). C) Raw volumes in cubic millimeters for each ROI from A.
Functional activation differences were more clear-cut and regionally distinct from anatomical differences (Fig. 2C). Regional normalized volume and cortical thickness did not differ between groups (RM-ANOVA carrier effect: both p > 0.8 not significant, n.s.; pairwise group comparisons within each of the five ROI: all p > 0.3) The RM-ANOVA on raw gray matter volume across the five ROIs showed a marginal main effect of carrier status (carrier mean = 15468 mm3, SE = 787, non-carrier mean = 17199, SE = 530, p = 0.090, n.s.) but group differences did not reach significance in any ROI (all uncorrected p > 0.08, n.s.). The only region in which group differences approached significance (p < 0.1) was aVLPFC (p = 0.089, n.s.). Logistic regression classifier based on aVLPFC raw volume yielded estimated out-of-sample classification accuracy of 69% (p > 0.3 compared to chance, n.s.; permutation test), at 82% specificity and 40% sensitivity. There were no raw volume differences in pVLPFC (p > 0.6, n.s.) even though that was the region with the strongest task-based activation differences.
DISCUSSION
We report the first task-based fMRI study of GRN mutation carriers prior to FTD onset, using a relational reasoning task known to engage lateral PFC and caudate [11]. Behaviorally, mutation carriers displayed slower response latencies for more complex problems. ROI analysis showed overall lowered functional activation in the mutation carriers in anterior and posterior ventrolateral PFC. Task-related activation in posterior VLPFC discriminated between mutation carriers and non-carriers with 94% accuracy, at 91% specificity, and 100% sensitivity. Volumetric analysis revealed that the task-related activation differences were not attributable to structural differences. Group differences in raw gray matter volume suggested a possible overall gray matter loss in mutation carriers. However, the volumetric differences did not reach statistical significance, did not reliably differentiate individual carriers from non-carriers, and appeared to be localized more anteriorly than activation differences.
Our findings of reduced VLPFC activation in mutation carriers align with the existing neuroimaging evidence highlighting early PFC functional changes in FTD-GRN that are not paralleled by structural changes, such as glucose metabolic reductions identified by FDG-PET [6], and prefrontal network disruptions identified by resting-state fMRI [9]. Consistent with these studies, we found reduced activation in lateral PFC regions in mutation carriers without clear-cut evidence of corresponding gray matter loss. Somewhat surprisingly, we found activation reductions in carriers compared to non-carriers at all levels of relational reasoning complexity, suggesting that task-based fMRI is highly sensitive to early GRN-related neurodegenerative changes even in conditions in which no behavioral differences were observed.
The caudate is another region reliably recruited in the matrix reasoning task used here [11] as well as a region previously reported in a FDG-PET study to have metabolic abnormalities associated with GRN. Interestingly, we did not find activation abnormalities in the caudate in the current study. Possibly, functional abnormalities in FTD-GRN originate early on in the PFC with secondary functional changes in the caudate. Such progression would be consistent with the pattern of progranulin expression that appears stronger in the cortex than striatum in mice [15].
The Genetic Frontotemporal dementia Initiative study, which includes the largest cohort of GRN mutation carriers evaluated to date, has estimated that the earliest volumetric brain changes occur 15 years before expected onset in this group [14]. Our mutation carriers were on average 5 years from expected onset but included a wide range. The smallness of our mutation carrier sample and our cross-sectional design limit the comparison of our results with this timeline. However, we note that decreased pVLPFC activation was present in all mutation carriers, regardless of years to expected onset and presence of CSND, with high estimated out-of-sample classification accuracy. This raises the possibility that reductions in PFC activation may be detectable earlier than structural changes, similarly to connectivity changes reported in mutation carriers in their fourth decade of life [8]. While we cannot address longitudinal functional changes associated with GRN mutations, and how they eventually lead to FTD, we hope that our findings will help inform the design of longitudinal and natural history approaches.
Our study was focused on functional activation in PFC regions but we recognize that temporal and parietal regions have also been implicated in early GRN disease. The GENFI data indicate gray matter volumetric loss approximately 10 years before estimated disease onset in these regions [14]. Longitudinal decreases in lateral temporal lobe have been reported even earlier, in mutation carriers with a mean age of 43 [5]. Other neuroimaging evidence has highlighted disruptions in fronto-temporal, fronto-parietal, and occipito-frontal networks [8, 16]. Thus, under appropriate task conditions, these regions may also reveal altered functional activation, possibly with sensitivity similar to the PFC.
In addition to these limitations, we acknowledge that RM ANOVA is not ideal for small, unbalanced designs. It should be noted that with the small sample size in each group and a strict significance threshold to correct for multiple comparisons, power for our statistical comparisons was low. While our approach ensured that all reported group differences are indeed reliable, additional differences may exist that we were not able to detect, and thus any null findings should be taken with caution. For this reason, we report all group differences in the figures as individual data points, so readers can evaluate for themselves all effects, whether supported by statistical significance or not.
Biomarkers for the preclinical and early stages of FTD-GRN are desperately needed to help map out the natural disease course and investigate the effects of potentially disease-modifying therapies along the predementia trajectory of FTD-GRN. Our results are preliminary but they highlight the potential of task-based PFC activation as a marker of early FTD-GRN that can be further evaluated in larger-scale longitudinal investigations and compared to other functional neuroimaging modalities.