
Abstract
Accumulating evidence suggests a diet high in protein and fiber may confer some protection against Alzheimer’s disease (AD). However, no human studies to-date have assessed the relationship between protein and fiber intake, and plasma and brain amyloid-β (Aβ). Consequently, this cross-sectional study, investigated the association of self-reported dietary intakes of protein and fiber, with plasma and brain Aβ burden (n = 541, and n = 162 respectively), in a well-characterized cohort of cognitively normal older adults, drawn from the larger Australian Imaging, Biomarkers and Lifestyle (AIBL) study of aging. We observed 12.59 and 8.43 higher odds of ‘high’ brain Aβ burden (PiB PET SUVR≥1.5) if protein intake fell in the lowest and middle tertile, respectively, compared to the highest tertile (p = 0.008; p = 0.013). Thus, in this cohort, the more protein consumed, the less likelihood of ‘high’ Aβ burden in the brain. No other significant associations were observed. The results of this study highlight the potentially protective impact of high dietary protein intake on brain Aβ burden in older adults, before objective memory decline is apparent. While longitudinal validation is required, these findings may assist in the development of dietary approaches aimed at preventing or delaying AD onset.
INTRODUCTION
Deposition of insoluble amyloid-β (Aβ) aggregates into extracellular neuritic plaques represents a well-described pathognomonic hallmark characteristic of Alzheimer’s disease (AD). Changes in plasma Aβ peptide levels have been proposed to reflect Aβ deposition occurring within the brain [1, 2], and to be associated with development of mild cognitive impairment (MCI) and AD [2, 3]. Positron emission tomography (PET) imaging studies have demonstrated that cerebral Aβ deposition commences well before detectable cognitive decline, and ∼15–20 years before clinical diagnosis of AD [4]; thereby presenting a potential window of opportunity to change disease trajectory.
Currently, no cure for AD exists, and it is therefore essential that effective and broadly applicable approaches to prevent or delay disease onset are developed. Research investigating the regulatory effects of dietary constituents on the onset and progression of AD neuropathology is gaining momentum (reviewed in [5]), and accumulating evidence suggests that dietary intakes of fiber and protein may be important contributors to the development of such preventative strategies [6, 7].
Low dietary intake of cereal fiber has been associated with increased risk of type-2 diabetes [8, 9], while a high fiber diet has been proposed to protect against obesity by lowering insulin levels [10]; importantly, type-2 diabetes and obesity are risk factors for AD [11]. Furthermore, dietary fiber containing phytosterols, and short chain fatty acids which are produced when dietary fiber is fermented in the colon, demonstrate cholesterol lowering, appetite regulation, and enhanced peripheral Aβ clearance effects; factors which are proposed to synergistically act to lower Aβ deposition in the brain [6].
Recent observational studies and clinical trials have suggested that increased protein intake, particularly from plant sources, may reduce blood pressure and attenuate increase in blood pressure over time, thereby decreasing risk of cardiovascular disease (CVD) [12–21]. These findings are of particular interest considering the wealth of data consistently proposing a strong association between CVD, its risk factors, and AD. [22–29]. Moreover, risk of MCI or dementia has been shown to be reduced among individuals adhering to a diet with high percentage caloric intake from protein sources [30].
Collectively these findings suggest that a diet high in protein and fiber may confer a degree of protection against AD risk. Yet, there have been no human studies directly assessing the relationship between protein and fiber intake, and plasma and brain Aβ. Consequently, the aim of this cross-sectional study was to determine the relationship between dietary protein and fiber intake, and plasma and brain Aβ, in a well-characterized cohort of cognitively normal (CN) older adults, drawn from the larger Australian Imaging, Biomarkers and Lifestyle (AIBL) study of aging [31]: we focused on CN older adults in order to avoid potential differential misclassification bias of dietary intake due to cognitive impairment.
MATERIALS AND METHODS
Study participants
This cross-sectional report utilizes baseline data collected from cognitively normal participants enrolled in the AIBL study [31]. At baseline, a total of 1,112 participants were recruited into the AIBL study cohort, including individuals classified as cognitively normal (n = 768), or meeting clinical criteria for mild cognitive impairment (MCI; n = 133) or AD (n = 211) [31]. Of the cognitively normal older adults, 723 completed the Cancer Council of Victoria Food Frequency Questionnaire (CCVFFQ) [32], data from which was used to determine intake of protein and fiber. Of these 723 participants, plasma Aβ data was available for 541, a subset of whom (n = 162) also underwent neuroimaging to determine brain Aβ burden.
Participants were classified as cognitively normal if their neuropsychological testing results were in normative ranges for their age and education [31]. All volunteers were aged 60 years and above at enrollment, and were excluded from study participation if they had a history of non-AD dementia, schizophrenia, bipolar disorder, significant current depression, Parkinson’s disease, cancer (other than basal cell skin carcinoma) within the last two years, symptomatic stroke, insulin-dependent diabetes, uncontrolled diabetes mellitus, or current regular alcohol use exceeding two standard drinks per day for women or four per day for men. Further details regarding AIBL study recruitment, assessment, inclusion and exclusion criteria are described in Ellis et al. [31]. The AIBL study is approved by the institutional ethics committees of Austin Health, St Vincent’s Health, Hollywood Private Hospital and Edith Cowan University [31].
Diet
The CCVFFQ is a 74-item self-administered semi-quantitative FFQ which assesses usual daily food intake over the preceding 12 months. The CCVFFQ has previously been validated relative to 7-day weighed food records [32]. The FFQ responses were analyzed by the Cancer Council in Carlton, Victoria. The food composition data used to calculate daily nutrient intake originates from the Nutrient Tables for use in Australia 1995 (NUTTAB95). Grams per day intake of protein and fiber, and kcal per day energy intake captured by the FFQ were utilized in the current analysis. A protein to fiber intake ratio was also calculated for each individual (using the grams per day intake of protein and fiber) and used in the current analysis.
Genotyping
Fasting blood samples were obtained using standard venepuncture of the antecubital vein and collected into EDTA tubes containing Prostaglandin E1 (PGE; Sapphire Bioscience, NSW, Australia, 33.3 ng/ml) to prevent platelet activation. Extraction of DNA from 5 ml of whole blood was undertaken using QIAamp DNA Blood Maxi Kits (Qiagen, Hilden, Germany) as per the manufacturer’s instructions. Specific TaqMan® (Thermo Fisher Scientific, Waltham, MA, USA) genotyping assays were used for ascertaining APOE genotype (rs7412, assay ID: C____904973_10; rs429358, assay ID: C___3084793_20), which were performed on a QuantStudio 12K Flex™ Real-Time-PCR system (Thermo Fisher Scientific, Waltham, MA, USA) using the TaqMan® GTXpress™ Master Mix (Thermo Fisher Scientific, Waltham, MA, USA).
Plasma amyloid-β measurement
Plasma Aβ1 - 40 and Aβ1 - 42 were measured using INNO-BIA plasma Aβ forms assays (Innogenetics, Inc., Gent, Belgium), a multiplex microsphere-based Luminex xMAP approach. The multiplex assays were run according to the manufacturer’s instructions, and as previously described in detail [2]. All plasma samples, Controls and Standards were run in duplicate. Assays were read using a Luminex xMAP reader system (Bio-Plex 200 System, Bio- Rad). A minimum of 100 beads per region were read for each sample. Data was analyzed using a 5-PL logarithmic equation in-built into the software of the instrument and automatically calculated for dilution factors. The plasma ratio of Aβ1 - 42/1 -40 was subsequently calculated using the Aβ1 - 42 and Aβ1 - 40 results.
Brain amyloid-β measurement
A subset of 162 participants underwent a [11C] Pittsburgh compound-B positron emission tomography (PiB PET) scan at the Austin Health Centre for PET, Melbourne, the Western Australia PET and cyclotron service, Sir Charles Gairdner Hospital or Oceanic Medical Imaging, Perth, as previously described [4, 33]. Briefly, each participant received an intravenous injection of ∼370 MBq [11C] PiB over 1 min. PET images were acquired 40–70 min post-PiB injection. A transmission scan was performed for attenuation correction. Acquired [11C] PiB PET images were processed using a semi-automatic sampling method. Standardised uptake values (SUV) for PiB were calculated for all brain regions examined and SUV ratios (SUVR) were generated by dividing all regional SUV by the cerebellar cortex SUV. Neocortical Aβ burden was expressed as the average SUVR of the area-weighted mean for the following brain regions: frontal (consisting of dorsolateral prefrontal, ventrolateral prefrontal and orbitofrontal regions), superior parietal, lateral temporal, lateral occipital, and anterior and posterior cingulate.
Statistical analyses
Statistical analyses were performed using IBM SPSS Statistics 22 for Windows Vista (SPSS Inc., Chicago, IL, USA), with a p-value of 0.05 or smaller determining a significant result. Means, standard deviations, and percentages are provided for the entire cognitively normal cohort, and subgroups of the cohort based on stratification by dietary protein and fiber intake (in tertiles). The three independent variables (fiber intake, protein intake, and the protein to fiber intake ratio) were divided into tertiles, with lowest intake denoted as tertile 1. Initially, general linear models were used to assess associations between the independent variable tertiles and the continuous dependent variables (plasma Aβ1 - 40, plasma Aβ1 - 42, and plasma Aβ1 - 42/1 -40 ratio). The general linear models included age, gender, years of education (≤12 and >12 years), APOE ɛ4 allele carriage (absence of ɛ4 allele versus presence of either one or two ɛ4 alleles), country of birth (Australian or other), body mass index (BMI; calculated as weight in kilograms divided by height in meters squared), and energy intake (kcal) as covariates. Using a pre-determined and well-published cut-off for ‘low’ and ‘high’ brain Aβ burden in the AIBL cohort [4], PiB PET SUVR was converted to a binary score with ‘0’ indicating ‘low Aβ burden’ in the brain (SUVR <1.5), and ‘1’ indicating ‘high’ brain Aβ burden (SUVR≥1.5). This binary score (‘low Aβ burden’ as the reference) was subsequently used in multinomial logistic regression models to calculate odds ratios for the dietary tertiles with the same confounding variables controlled for as in the general linear models. False discovery rate (FDR; R environment, version 3.3.2) was used for corrections for multiple comparisons [34].
RESULTS
This cohort comprised 541 cognitively normal participants (41% male) with average age of 69.84 years. Over 70% of participants were born in Australia, more than 45% had 12 or less years of education and 26% were carriers of the APOE ɛ4 allele. The cohort had an average body mass index of 26.57 kg/m2, with an average daily energy intake of 1693.06 kcal, mean daily protein and fiber intakes of 84.44 and 22.96 grams respectively, and an average protein to fiber intake ratio of 3.82 (Table 1). With respect to plasma measures of Aβ, the cohort had an average Aβ1 - 40 level of 151.43 pg ml-1, an average Aβ1 - 42 level of 33.81 pg ml-1, and an average Aβ1 - 42/1 -40 ratio of 0.23 (Table 1). Of the subset of participants who underwent PiB PET (n = 162), the average PiB SUVR was 1.41 (Table 1). Following stratification of the cohort by dietary protein and fiber intake tertiles, both increasing protein and fiber intakes were inversely associated with the percentage of individuals reporting 12 or less years of education (χ2; p = 0.043 and p = 0.002, respectively). Further, as expected, more males than females reported the highest intakes (tertile 3) of dietary protein and fiber (χ2; in both instances, p < 0.001), and increasing intakes of dietary protein and fiber were associated with increasing energy intake (F; in both instances, p < 0.001; Table 1).
Descriptive statistics for the entire cognitively normal cohort who completed the Cancer Council of Victoria food frequency questionnaire, and subgroups of the cohort based on stratification by dietary protein and fiber intake tertiles
Unless otherwise described, data are presented as mean±standard deviation of the mean. Bold indicates statistical significance (p < 0.05); characteristics compared using Analysis of variance (ANOVA) for continuous variables and χ2 for categorical variables. Tertile 1 = lowest dietary intake, Tertile 3 = highest dietary intake. *Body Mass Index calculated as weight in kilograms divided by height in meters squared. †PiB PET imaging was undertaken on a subset of 162 participants. Aβ, amyloid-β; APOE, Apolipoprotein E; g/day, grams per day; kcal, kilocalories; kg, kilograms; m2, meters squared; PiB PET, Pittsburgh compound B positron emission tomography; pg ml-1, picograms per milliliter; SUVR, standardized uptake value ratio; y, years.
In the general linear models assessing the association between intake of protein, fiber, or the protein to fiber intake ratio, and measures of blood Aβ, no significant associations were observed (n = 541; Table 2).
Relationship between dietary intake of protein and fiber and amyloid beta levels measured in the plasma of the cognitively normal cohort (n = 541)
General linear models; F- and p-values shown. Model adjusted for age, gender, years of education, APOE ɛ4 allele carrier status, country of birth (Australia versus Other), body mass index, and energy intake. Aβ, amyloid-β; APOE, Apolipoprotein E.
In the multinomial logistic regression models assessing the association between intake of protein, fiber or the protein to fiber intake ratio, and ‘low’ or ‘high’ brain Aβ (i.e., ‘low Aβ’, SUVR <1.5, or ‘high Aβ’, SUVR≥1.5), there were 12.59 higher odds of demonstrating ‘high’ brain Aβ burden if protein intake was in the lowest intake tertile compared to the highest intake tertile (Confidence Interval, CI; 1.71–93.02: p = 0.008). Additionally, there were 8.43 higher odds of demonstrating ‘high’ brain Aβ burden if protein intake was in the middle intake tertile compared to the highest intake tertile (CI; 1.74–40.84: p = 0.013; Table 3 and Fig. 1). Thus, in this neuroimaging subset of the cohort, the more protein consumed, the less likelihood of having ‘high’ brain Aβ burden. No other significant associations were observed (n = 162; Table 3).
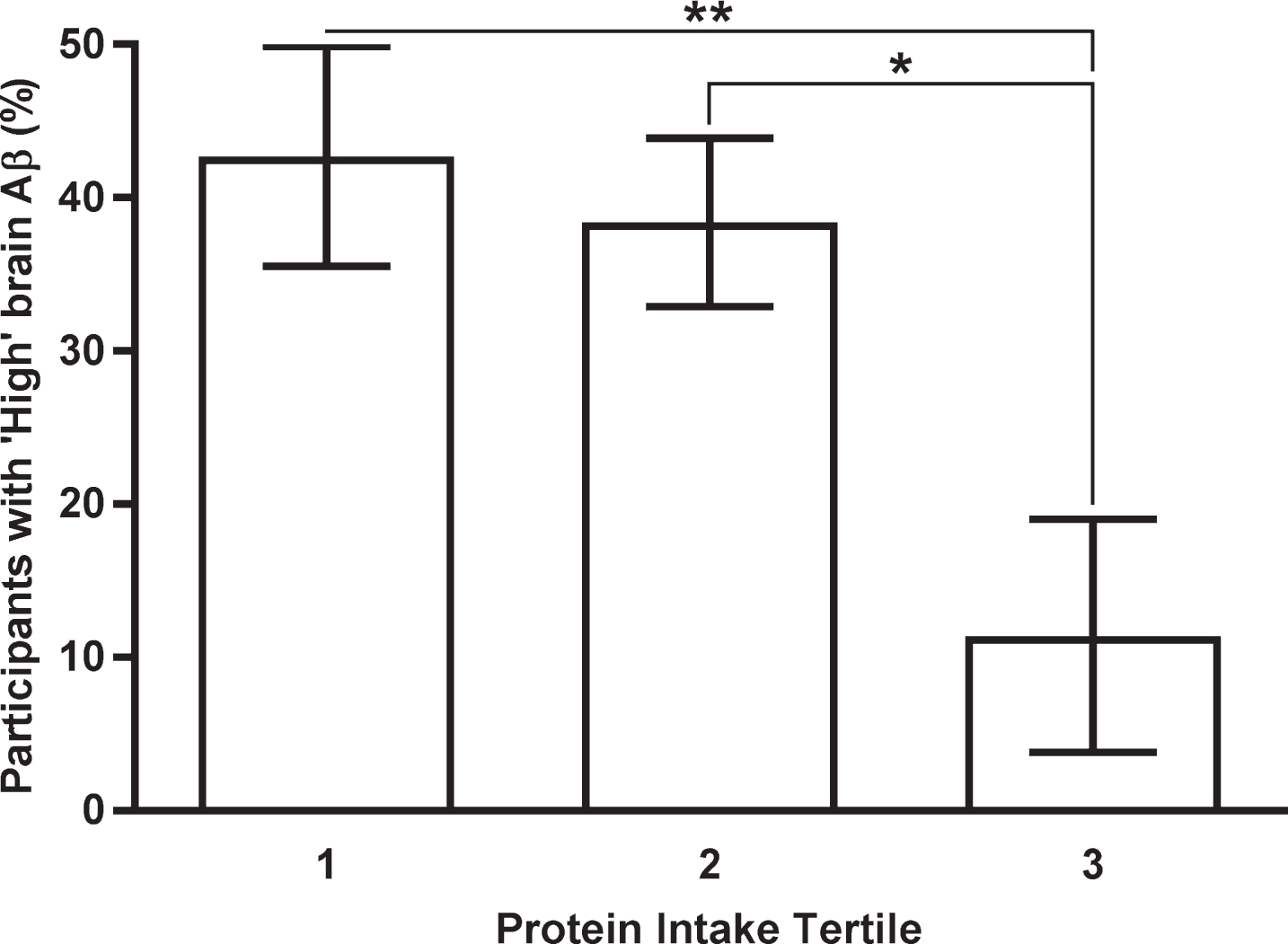
Percentage of participants in each protein intake tertile with ‘high’ brain amyloid beta burden. Means and standard deviations shown. Tertile 1 = lowest dietary intake, Tertile 3 = highest dietary intake. ‘High’ brain amyloid beta burden defined as PiB PET SUVR≥1.5. Means and standard deviations adjusted for age, gender, years of education, APOE ɛ4 allele carrier status, country of birth (Australia versus Other), body mass index, and energy intake. *p < 0.05, **p < 0.01. Aβ, amyloid-β; APOE, Apolipoprotein E; PiB PET, Pittsburgh compound-B positron emission tomography; SUVR, standardized uptake value ratio.
Odds ratios of demonstrating ‘high’ brain amyloid beta burden when comparing dietary intake tertiles of protein and fiber in the subset of cognitively normal cohort participants who underwent PiB PET (n = 162)
Multinomial logistic regression analysis used to calculate odds ratios. Tertile 1 = lowest dietary intake, Tertile 3 = highest dietary intake. ‘High’ brain amyloid beta burden defined as PiB PET SUVR≥1.5. Model adjusted for age, gender, years of education, APOE ɛ4 allele carrier status, country of birth (Australia versus Other), body mass index, and energy intake. Bold indicates statistical significance (*p < 0.05, **p < 0.01). †p-value remained significant (p < 0.05) after false discovery rate correction. APOE, Apolipoprotein E; CI, confidence interval; PiB PET, Pittsburgh compound-B positron emission tomography; SUVR, standardized uptake value ratio.
DISCUSSION
The objective of this study was to undertake a cross-sectional analysis to investigate the association of dietary protein and fiber intake, with plasma and brain Aβ (markers of AD pathology), in a highly-characterized cohort of cognitively normal older adults. Low and medium protein intakes were associated with increased likelihood of demonstrating significant brain Aβ burden (‘high’ Aβ) compared to high protein intake; indicating that in this cohort, the more protein participants consumed, the less likely they were to have ‘high’ Aβ burden in the brain. No other significant associations were observed. These data suggest that a diet high in protein may be potentially beneficial for maintaining brain health in Australian older adults.
To our knowledge, there have been no other human studies directly assessing the relationship between protein and fiber intake, and plasma and brain Aβ. While validation of our results is required longitudinally, and further substantiated via intervention, our findings regarding protein intake, appear to be consistent with related published studies. Several studies have linked high dietary protein intake to reduced blood pressure [21, 35–37], thereby presenting a potential mediator of positive effects of high protein intake on brain Aβ burden.
Hypertension is a risk factor for several diseases including CVD and AD, and CVD itself is proposed to increase AD risk [22–29]. Results from the INTERMAP study of 4,680 participants aged 40 to 59 years from four countries, found vegetable protein intake, but not animal protein or total protein intake, to be inversely related to blood pressure [21]. Appel et al. [35] conducted a randomized, three-period, crossover feeding study in 164 adults with prehypertension or stage 1 hypertension. Each feeding period lasted six weeks and the effect of three diets (rich in carbohydrates, rich in protein with half from plant sources, and rich in unsaturated fat with mainly monounsaturated fat) on blood pressure and serum lipids were compared. Blood pressure, low-density lipoprotein cholesterol and estimated coronary heart disease risk were all lower on each diet compared with baseline. Notably, compared with the carbohydrate diet, the protein diet further decreased mean systolic blood pressure by 1.4 mm Hg (p < 0.01) and by 3.5 mm Hg (p < 0.01) among those with hypertension. The authors concluded that partial substitution of carbohydrates with protein can lower blood pressure and reduce estimated CVD risk.
Prercisely how reduced blood pressure following high dietary protein intake would be associated with lower brain Aβ burden is unknown, although animal studies likely offer some mechanistic insight. Results from a study by Gentile et al. [38] using a mouse model of arterial hypertension suggest that chronic hypertension may result in Aβ deposition through increased blood-brain barrier permeability. Further, in another mouse model of hypertension, hypoperfusion and neuroinflammation preceded Aβ plaque deposition in the cortex and hippocampus [39]. Consistent with the studies reported above, in our total cohort (n = 541), increased protein intake was associated with decreased systolic blood pressure (SBP; partial correlation r = –0.087, p = 0.049, controlling for age, gender, years of education, APOE ɛ4 allele carrier status, country of birth (Australia versus Other), body mass index, and energy intake; data not shown). However, when conducting a mediation analysis on the subset of participants who underwent PiB PET (n = 162), we did not see a mediating effect of SBP on the relationship between protein intake and brain Aβ (path ab; bootstrapped indirect path = –0.004, CI = 0.17, 0.08 (data not shown); conducted using Hayes’ simple mediation model 4 [40]): future studies should investigate this association further.
Another potential mechanism that may explain why higher protein intake is associated with lower brain Aβ burden could relate to the fact that long-term ingestion of a diet high in protein has been shown to be associated with increased satiety and reduced adiposity [41]. Such improved body composition would result in reduced plasma insulin levels thereby lowering insulin resistance (reviewed in [42]): an important outcome given that higher insulin resistance has been associated with postmortem brain Aβ burden [43]. Furthermore, the reduction in body adiposity would result in decreased cytokines and other inflammatory agents known to be associated with increased Aβ burden in the brain [44, 45]; it should be noted, however, that high protein diets have been inconsistently associated with levels of inflammation determined by the non-specific acute phase reactant C-reactive protein [46–48].
Protein intake has also been shown to enhance memory performance in healthy elderly participants; findings of particular interest in the context of the current study. Twenty-two participants (50% male) aged 61–79 years consumed a 300 mL drink containing 774 kJ of pure protein (whey). Cognitive testing was administered 15 and 60 min after ingestion of the drink. Compared with placebo, the protein drink improved delayed paragraph recall and immediate paragraph recall and reduced the rate of ‘forgetting’ on the paragraph recall at 15 min (p < 0.05) [49].
Published studies propose a link between increased fiber intake and decreased risk of AD and AD-related diseases including CVD, diabetes, and obesity [6, 51]. Moreover, transgenic AD mice fed diets rich in fiber have demonstrated improved memory and learning [52, 53], and lower levels of plasma Aβ [53]. Unexpectedly, however, we did not find an association between fiber intake and measures of Aβ in the current study. Our study examined the association between total fiber intake and measures of plasma and brain Aβ; it is conceivable that a specific type or source of fiber (e.g., soluble, insoluble, cereal, fruit, or vegetable fiber) mediates the positive effects reported by others, and that such an effect is concealed when all fiber types and sources are grouped together. Future studies may benefit from examining fiber types or sources individually. Furthermore, the Australian National Health and Medical Research Council (NHMRC) and the New Zealand Ministry of Health (MoH) define an Adequate Intake (AI) of dietary fiber as 30 and 25 grams per day for adult men and women, respectively [54, 55]. However, in our cohort, mean daily fiber intake was 22.96 grams per day; suggesting that fiber intake may have been too low to confer significant benefit in the context of brain Aβ. By contrast, mean daily protein intake in our cohort (84.44 grams per day) was above Recommended Dietary Intake (RDI; 64–81 and 46–57 grams per day for older men and women, respectively) [54, 55].
There are limitations to our report; this is a cross-sectional, observational study; therefore, we can draw no conclusions regarding causality or rates of brain Aβ accumulation. Further, the CCVFFQ utilized in this study relies on participants’ estimations of food intake over the previous year; this is a common limitation of studies of diet and can potentially lead to misclassification of dietary intake due to limited accuracy. This is particularly important with regards to cognitively impaired participants, thus, to circumvent this limitation, only cognitively normal older adults were included in the current analysis. Nevertheless, we recognize that ideally, an objective measure, such as a blood-based biomarker panel, that reflects intake levels of the nutrients of interest, would be utilized as an additional approach to evade the limitations of self-report.
Many aspects of our study, however, provide confidence in our findings. We have utilized a well-characterized cohort thereby increasing the internal validity of our results. We have taken a conservative approach to analysis by controlling for a range of demographic variables. Moreover, the dietary data were collected using an instrument previously validated in epidemiological studies [56].
In summary, the results of this study highlight the potentially beneficial impact of high dietary protein intake on brain Aβ burden in older cognitively unimpaired adults. While validation of these results is required longitudinally, these findings may assist in the development of approaches or interventions aimed at preventing or delaying AD onset.
Footnotes
ACKNOWLEDGMENTS
The AIBL team wish to thank the participants of the AIBL study for their commitment and dedication to helping advance research into the early detection and causation of AD, and the clinicians who referred patients to the study.
Funding for the study was provided by the CSIRO Flagship Collaboration Fund and the Science and Industry Endowment Fund (SIEF) in partnership with Edith Cowan University (ECU), The Florey Institute of Neuroscience and Mental Health, Alzheimer’s Australia (AA), National Ageing Research Institute (NARI), Austin Health, CogState Ltd., Hollywood Private Hospital, and Sir Charles Gairdner Hospital. The study also received funding from the National Health and Medical Research Council (NHMRC), the Dementia Collaborative Research Centres program (DCRC2), the Cooperative Research Centre (CRC) for Mental Health – funded through the CRC Program (an Australian Government Initiative, Grant ID:20100104), and the McCusker Alzheimer’s Research Foundation, and Operational Infrastructure Support from the Government of Victoria.