
Abstract
Generation of amyloid-β peptides (Aβs) by proteolytic cleavage of the amyloid-β protein precursor (AβPP), especially increased production of Aβ42/Aβ43 over Aβ40, and their aggregation as oligomers and plaques, represent a characteristic feature of Alzheimer’s disease (AD). In familial AD (FAD), altered Aβ production originates from specific mutations of AβPP or presenilins 1/2 (PS1/PS2), the catalytic subunits of γ-secretase. In sporadic AD, the origin of altered production of Aβs remains unknown. We hypothesize that the ‘human chemical exposome’ contains products able to favor the production of Aβ42/Aβ43 over Aβ40 and shorter Aβs. To detect such products, we screened a library of 3500 + compounds in a cell-based assay for enhanced Aβ42/Aβ43 production. Nine pyrazole insecticides were found to induce a β- and γ-secretase-dependent, 3-10-fold increase in the production of extracellular Aβ42 in various cell lines and neurons differentiated from induced pluripotent stem cells derived from healthy and FAD patients. Immunoprecipitation/mass spectrometry analyses showed increased production of Aβs cleaved at positions 42/43, and reduced production of peptides cleaved at positions 38 and shorter. Strongly supporting a direct effect on γ-secretase activity, pyrazoles shifted the cleavage pattern of another γ-secretase substrate, alcadeinα, and shifted the cleavage of AβPP by highly purified γ-secretase toward Aβ42/Aβ43. Focusing on fipronil, we showed that some of its metabolites, in particular the persistent fipronil sulfone, also favor the production of Aβ42/Aβ43 in both cell-based and cell-free systems. Fipronil administered orally to mice and rats is known to be metabolized rapidly, mostly to fipronil sulfone, which stably accumulates in adipose tissue and brain. In conclusion, several widely used pyrazole insecticides enhance the production of toxic, aggregation prone Aβ42/Aβ43 peptides, suggesting the possible existence of environmental “Alzheimerogens” which may contribute to the initiation and propagation of the amyloidogenic process in sporadic AD.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is a major disease in countries with aging populations. Despite unresolved questions on the initial causes of AD and numerous clinical trial failures in recent years, the lack of satisfactory treatments and the extremely high prevalence of AD calls for fundamental research to determine its underlying molecular and cellular causes and mechanisms and for applied research to identify options for prevention, therapeutic targets and disease-modifying drug candidates (reviews in [1–4]).
Although the aggregation of specific forms of amyloid-β peptides (Aβs) into soluble oligomers is undoubtedly associated with the onset of AD, the upstream etiological events underlying this pathological process remain unclear, including whether there is an absolute, or relative, increase in the production of longer, aggregation-prone Aβ species. Aβs are derived from the successive action of two proteases, β-secretase and γ-secretase, on one of their numerous substrates, the transmembrane amyloid-β protein precursor (AβPP). According to its cleavage site, γ-secretase liberates Aβs of different sizes. While Aβ40 is considered as the main physiological product, the appearance of Aβ42 and Aβ43, or increase of the Aβ42-Aβ43/Aβ40 ratio is clearly associated with the onset of AD [5–7]. These longer Aβs show an increased propensity to form oligomers, which can assemble into large extracellular deposits, the amyloid plaques, a characteristic feature of AD. Aβ42/Aβ43 oligomers are now considered as the toxic, AD initiating elements rather than the more prominent plaques initially discovered by Aloïs Alzheimer. This ‘amyloid cascade theory’ is further supported by the identification of the genetic causes of early onset familial AD (FAD). These rare forms of AD (less than 1% of all AD cases) are indeed all associated with specific mutations of either AβPP or PS1 or PS2 (Presenilins) genes (the latter encode the catalytic subunits of the γ-secretase complex) (review in [8, 9]). In addition, a specific, protective mutation of AβPP was recently found to be associated with reduced risk for developing AD [10]. Furthermore, animal models that express mutated forms of AβPP and/or PS1, or human Aβ42/Aβ43, develop molecular, cellular and cognitive deficits reminiscent of AD ([6], review in [11, 12]). Yet, the level of the different forms of Aβs is not well correlated with cognitive deficits in mice, suggesting that Aβs may represent markers rather than inducers of AD in these models [13]. An altered processing of γ-secretase substrates other than AβPP might also be important in the pathogenesis of AD. Although FAD is clearly a consequence of genetic mutations of genes encoding the proteins responsible for the production of Aβs, the initial trigger(s) of late-onset, sporadic AD (>99% of AD cases) remain unknown, despite extensive genome-wide association studies (GWAS) and the identification of various risk factors (review in [14]).
Environmental neurotoxic agents including pesticides, organic solvents, metals and some natural toxins (cyanobacteria) are likely sources of AD-inducing factors (reviews in [15–25]). According to the US Environmental Protection Agency (EPA) Toxic Substances Control Act, over 84,000 chemicals are manufactured or imported at levels >10 tons per year, not including pesticides, cosmetics, food stuffs, and food additives which are covered by other legislations (http://www.epa.gov). It is estimated that we are exposed to over 85,000 substances which, along with all natural substances to which we are exposed from conception to death, constitute the ‘human chemical exposome’ (HCE) [26–34]. In search for potential “Alzheimerogens” (named by analogy with carcinogens), and encouraged by the discovery of Aftins (Amyloid β Forty-Two Inducers) [35–37], a class of synthetic compounds which specifically induce the production of Aβ42, we have started to assemble and screen a library of compounds, belonging to the HCE, for Aβ42-inducing products. Our first study identified some triazine herbicides as products stimulating the production of Aβ42/Aβ43 in numerous cell lines [38].
In this new study, we show that several members of the pyrazole class of insecticides, exemplified by fipronil, also increase the γ-secretase-dependent production of Aβ42/Aβ43 peptides in various cell lines and in a cell-free system with highly purified γ-secretase. Interestingly fipronil is metabolized to fipronil sulfone, a very persistent product that accumulates in adipose tissue and brain. These results show that the HCE contains various brain permeable products able to induce the production of pathogenic Aβs. If long exposures occur in real life and if accumulation of Aβ42/Aβ43 is sufficient to trigger neurodegeneration/neuroinflammation, such products may contribute to the initiation, development and acceleration of sporadic AD and might thus be collectively qualified as potential “Alzheimerogens”. Some of them might also be used to develop useful chemically-induced animal models of AD.
MATERIAL AND METHODS
Material and methods are described in full in the Supplementary Material They include: 1) Pyrazoles and other reagents; 2) Cell cultures: cell lines, iPSCs-derived neurons and HEK293-alcadeinα cells; 3) Purified γ-secretase preparation; 4) Aβ cell-based and cell-free assays; 5) Cell viability assay; 6) Pharmacokinetics studies; 7) Proteomics study.
RESULTS
Screening HCE compounds reveals pyrazole insecticides as Aβ42 inducers
We first screened a library of over 3,500 low molecular weight compounds representative of the HCE for their ability to induce the production and secretion of Aβ42 by N2a cells stably expressing AβPP695 (N2a-AβPP695). An MTS-based cell viability assay was run in parallel to assess cell survival. Although the vast majority of compounds were unable to induce Aβ42 production, a few active products including several triazine herbicides [38] and pyrazole insecticides (this article) were identified. We next assembled a small library of 18 pyrazoles (
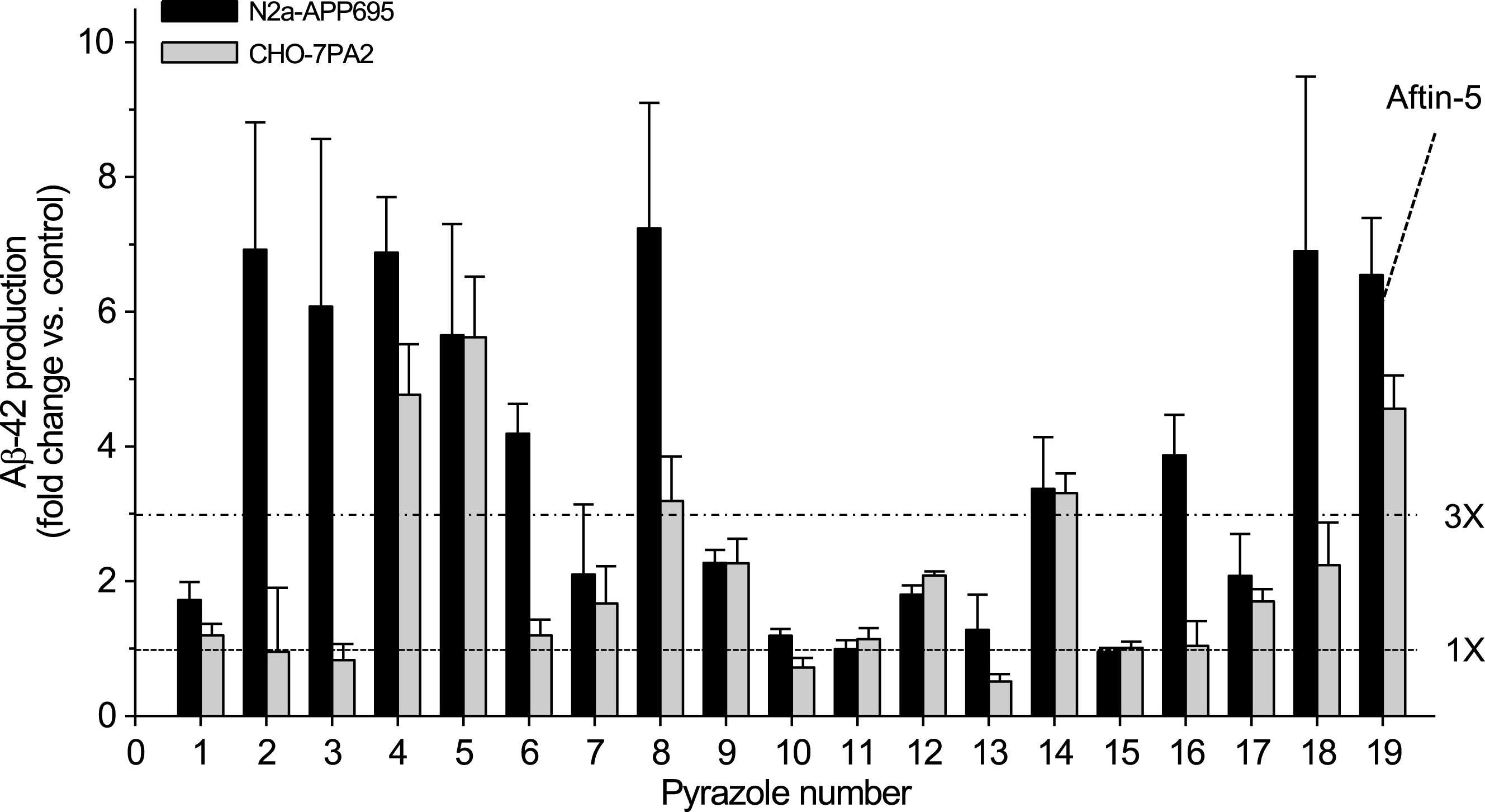
Some pyrazoles trigger the production of extracellular amyloid Aβ42. Effect of 18 pyrazoles on extracellular amyloid Aβ42 production by N2a-APP695 and CHO-7PA2-APPsw cells. Cells were treated with 100μM of each compound for 18 h and cell supernatants were collected for extracellular Aβ42 levels measurement by an ELISA assay. Aftin-5 was used as a positive control and the corresponding volume of vehicle (DMSO) was used as a negative control. Levels are expressed as fold change, ± SE, of Aβ42 levels over the Aβ42 level of control, vehicle-treated cells. Average of five experiments performed in triplicate. Horizontal dotted lines indicate levels for 1- and 3- fold increases in Aβ42 concentration.
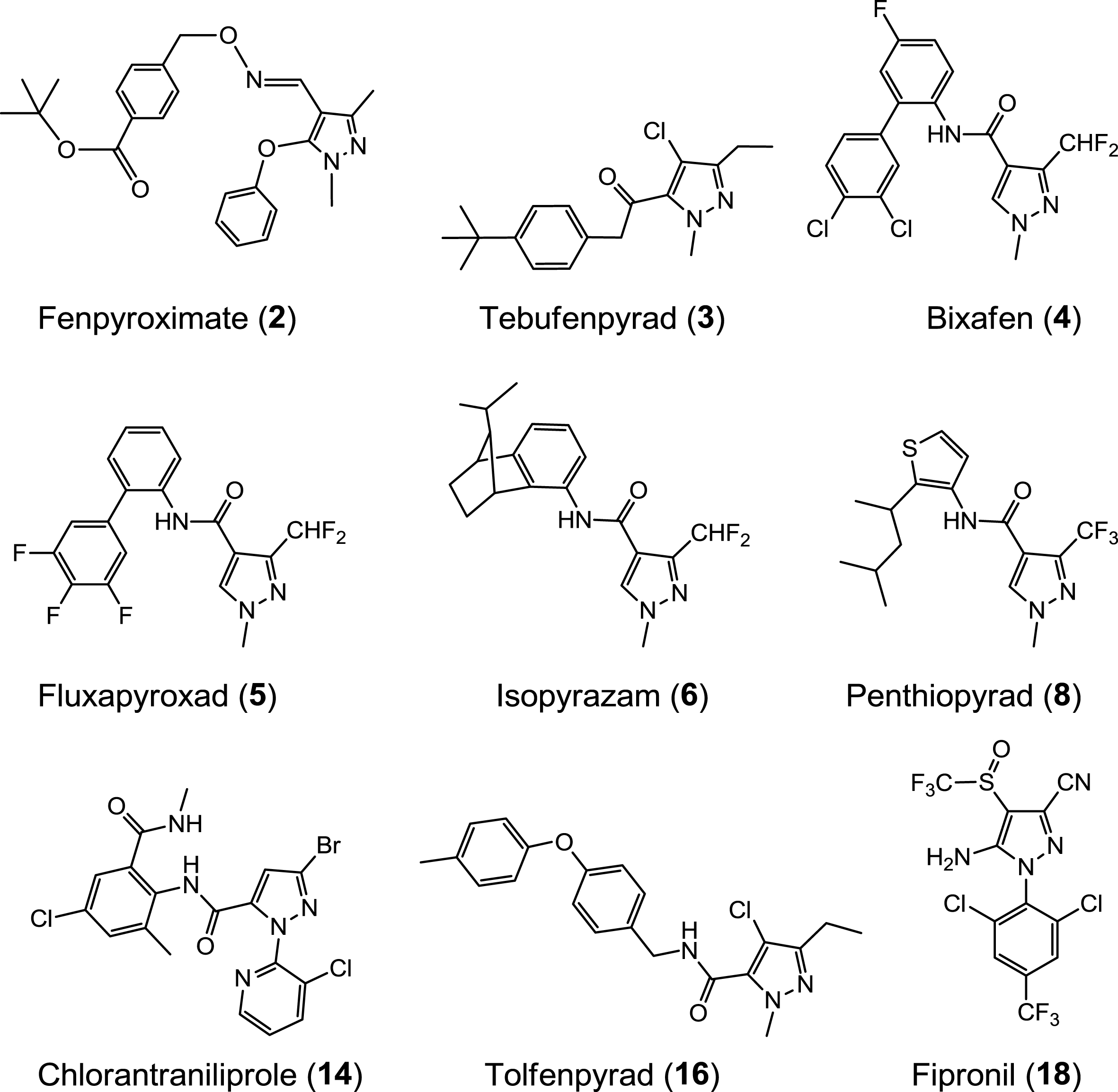
Molecular structure of the nine active pyrazoles.
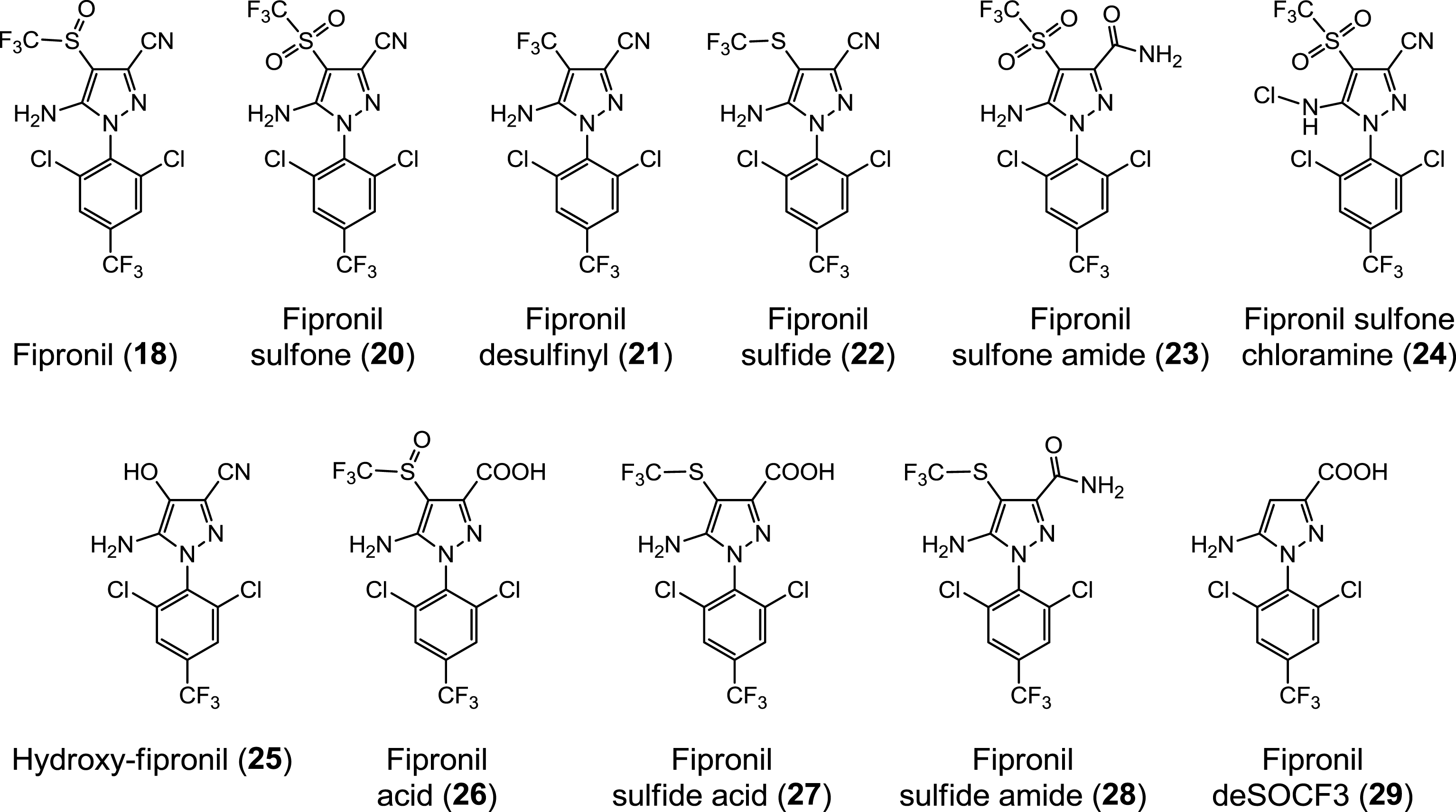
Molecular structure of fipronil and some of its metabolites and derivatives.
Concentrations of fipronil sulfone reached in mammals following fipronil administration (oral, ip, iv, general exposure). Fipronil sulfone (
Aβ42 production triggered by pyrazoles in N2a-APP695 cells requires β- and γ-secretase activity as demonstrated by the fact that it was strongly inhibited by β- (inhibitor IV) and γ-secretases (BMS 299897, DAPT) inhibitors and by a γ-secretase modulator (‘Torrey Pines’ compound, inducing an increase in Aβ38 and a decrease in Aβ42 and Aβ40) [48, 49] (Fig. 4). Aβ38 production was also strongly reduced following pyrazole treatment, while Aβ40 levels were only modestly affected, as measured by ELISA (data not shown) and by mass spectrometry (Fig. 5). Increased Aβ42 production triggered by pyrazoles was further confirmed in HEK293 cells stably expressing AβPPsw (data not shown) and neurons derived from human iPSCs (see below).
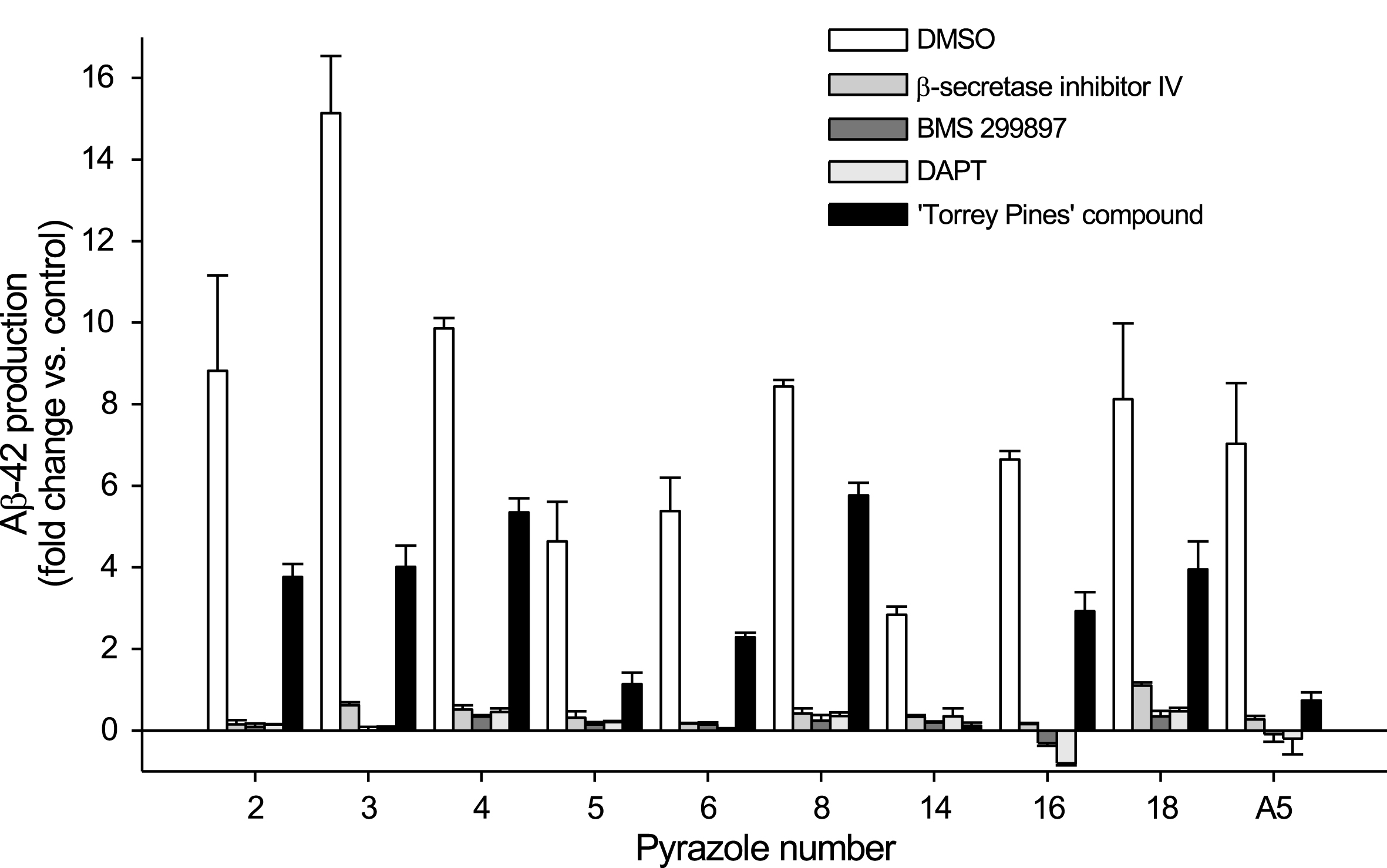
Extracellular Aβ42 production induced by pyrazoles is inhibited by β-secretase inhibitor IV, γ-secretase inhibitors DAPT and BMS 299897, and γ-secretase modulator ‘Torrey Pines’. N2a-APP695 cells were exposed to 10μM β-secretase inhibitor IV, 2μM BMS 299897, 2μM DAPT or 10μM ‘Torrey Pines’ compound. 1.5 h later cells were exposed to 50μM (
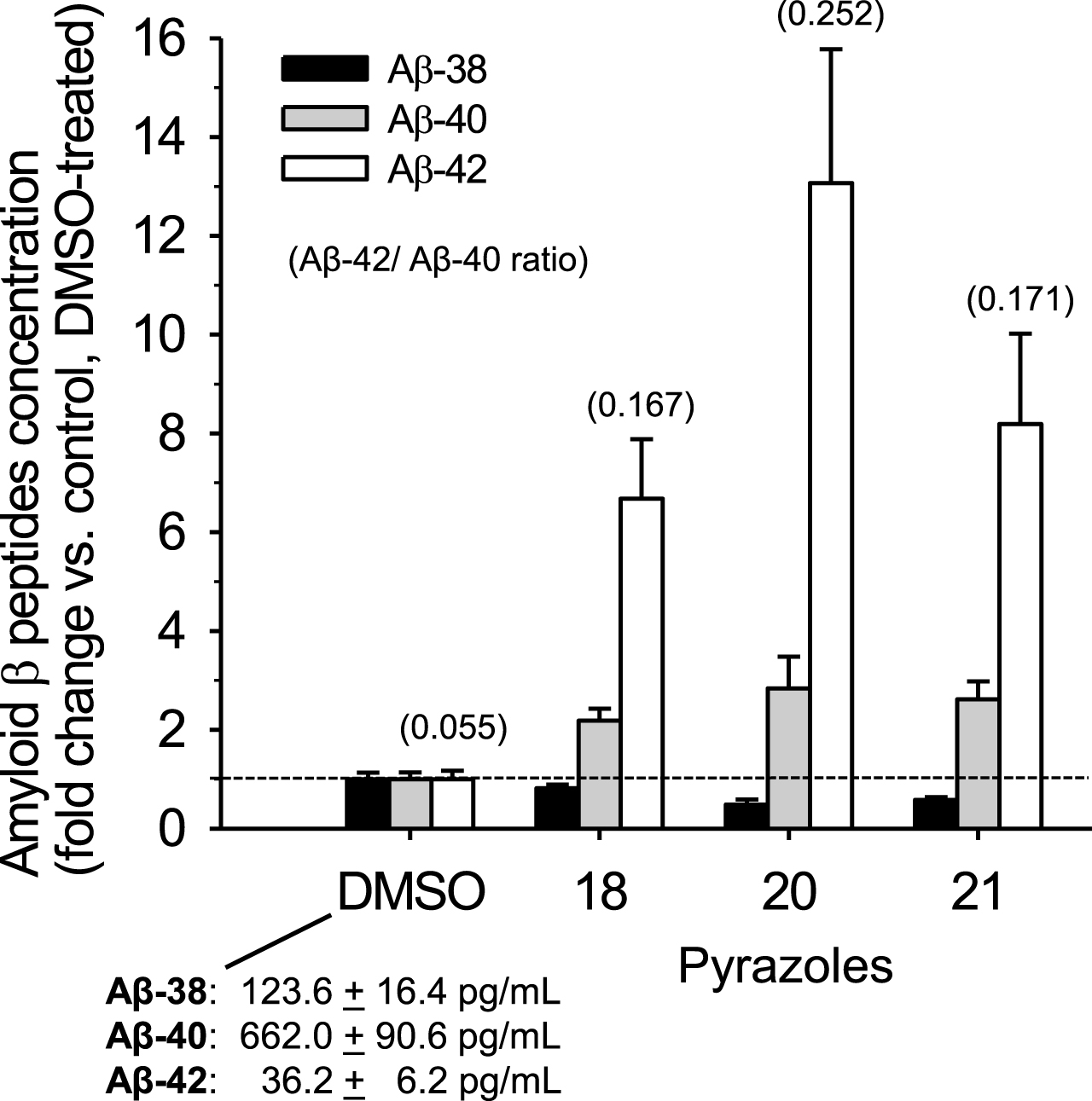
Absolute quantification of Aβ38, Aβ40, and Aβ42 using LC-MS/MS. Levels of the three Aβs were determined by mass spectrometry in supernatants of N2a-APP695 cells following 18 h treatment with DMSO, 100μM of pyrazoles
Mass spectrometric quantification and profile analysis of pyrazole-induced Aβs
Aβ38, Aβ40, and Aβ42 were quantified in N2a-AβPP695 cell culture supernatants using LC-MS/MS [50, 51]. Like aftins [35, 36] and triazines [38], pyrazoles induced a reduction in Aβ38 levels, a slight increase in Aβ40 levels and a strong increase in Aβ42 levels (Fig. 5).
We next analyzed, by IP-MS [52], the profile of Aβs produced by N2a-AβPP695 exposed to fipronil (
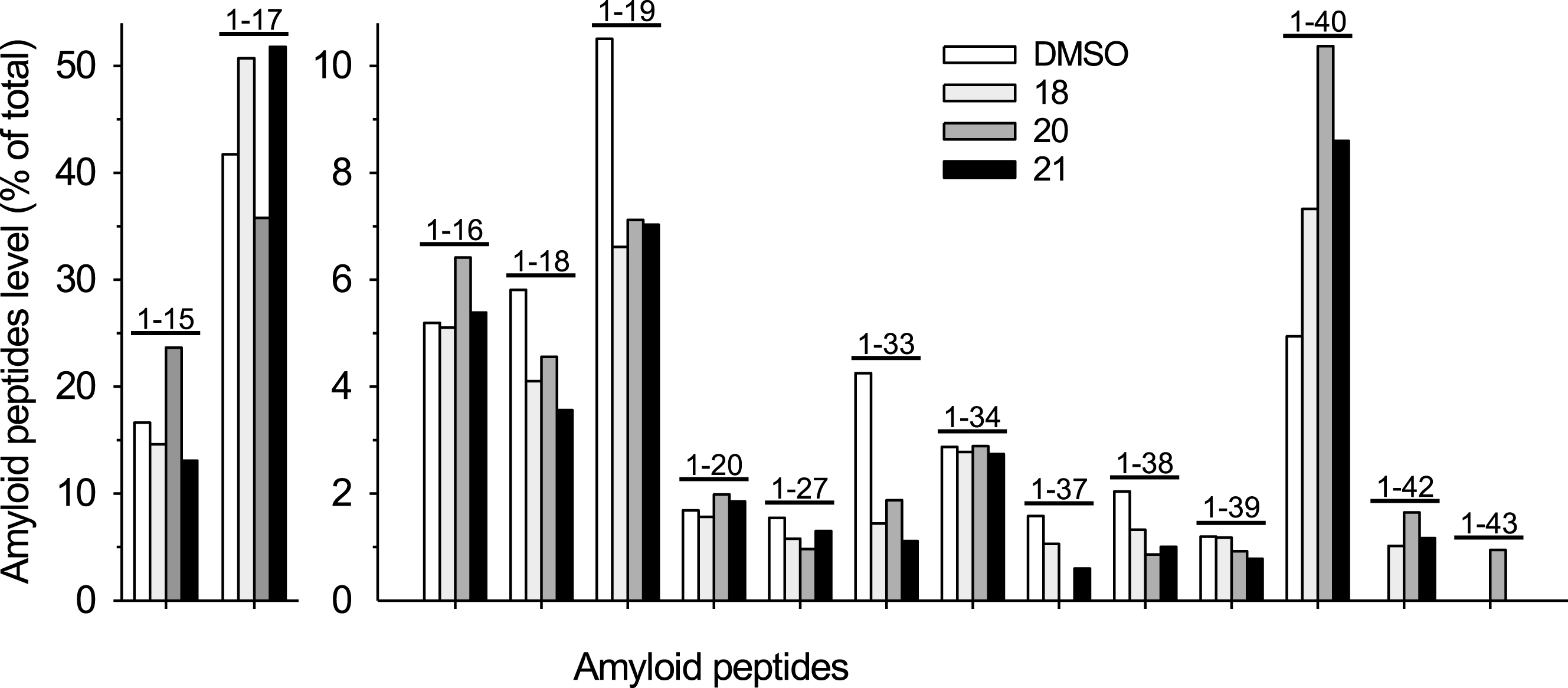
Pattern of Aβs produced by N2a-APP695 cells exposed to pyrazoles
Neurons differentiated from human iPSCs of AD patient and healthy control
We next tested the effects of the active pyrazoles and aftin-5 on neurons differentiated for 4 weeks from human iPSCs derived from a healthy individual (AβPP WT, wild-type) or from a patient with familial AD (AβPP K724N mutation) [56–58] (Fig. 7). AβPP K724N neurons produced more Aβ42 versus Aβ40 compared to AβPP WT neurons. As observed with triazines [38], addition of any of the active pyrazoles or aftin-5 resulted in further increase in Aβ42 production, in both AβPP WT and AβPP K724N neurons (Fig. 7).
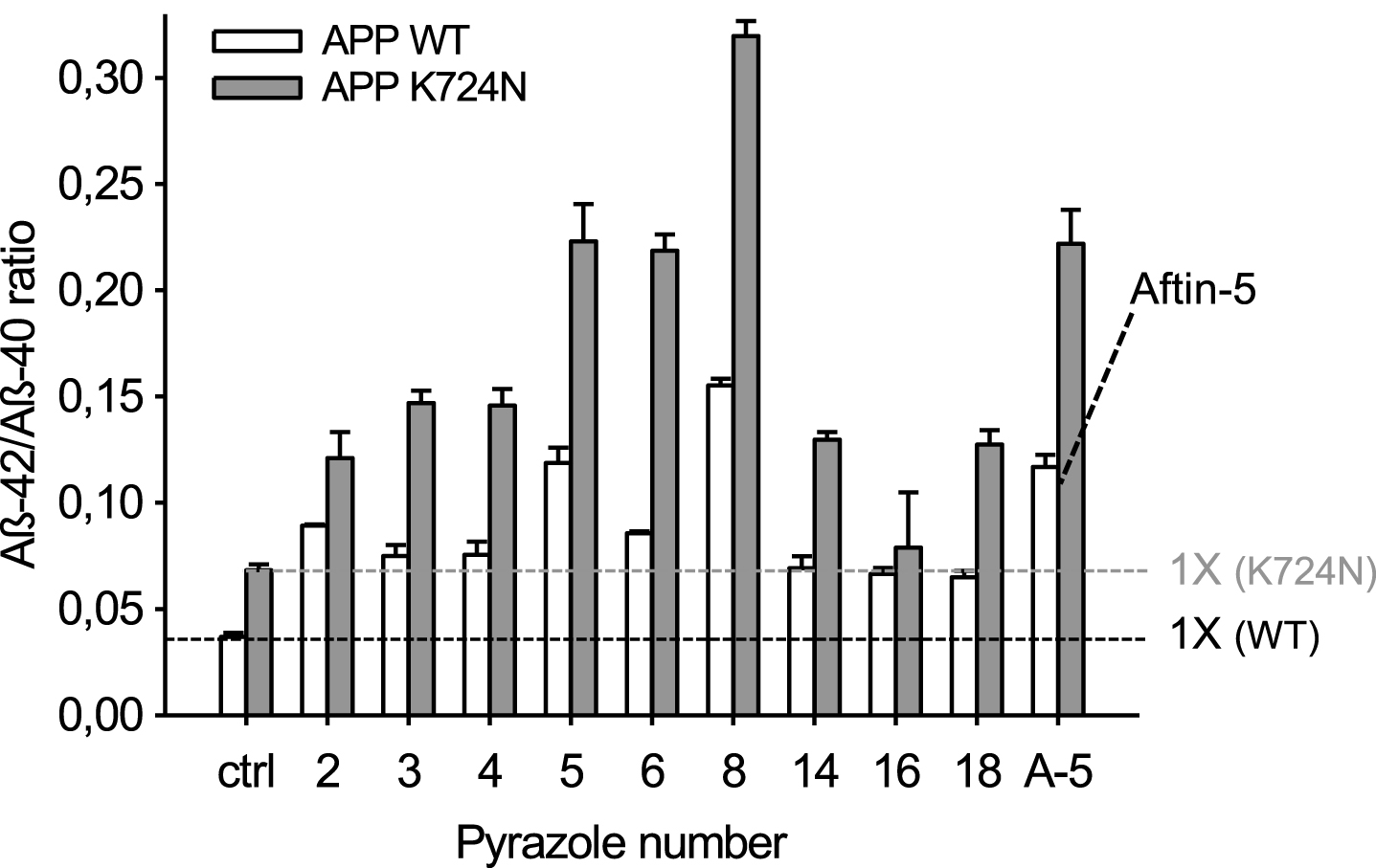
Pyrazoles trigger enhanced production of Aβ42 versus Aβ40 in neurons differentiated from human induced pluripotent stem cells. Neurons were derived from iPSCs obtained from healthy donor (APP WT, white bars) or from an AD patient with APP K724N mutation (grey bars). They were exposed for 24 h to DMSO (control), 100μM aftin-5 or the nine pyrazoles. Cell supernatants were collected for extracellular Aβ40 and Aβ42 levels measurement by an ELISA assay. Levels are expressed as Aβ42/Aβ40 ratios ± SE of triplicate values. Horizontal dotted line indicates level for basal Aβ42/Aβ40 ratios.
Pyrazoles affect the specificity of APP-C99 cleavage and Aβ production in a cell-free γ-secretase activity assay
In order to assess whether the pyrazole-based compounds modulate Aβ production through a direct effect on the specific processing of APP-C99 by the γ-secretase complex, we tested fipronil (
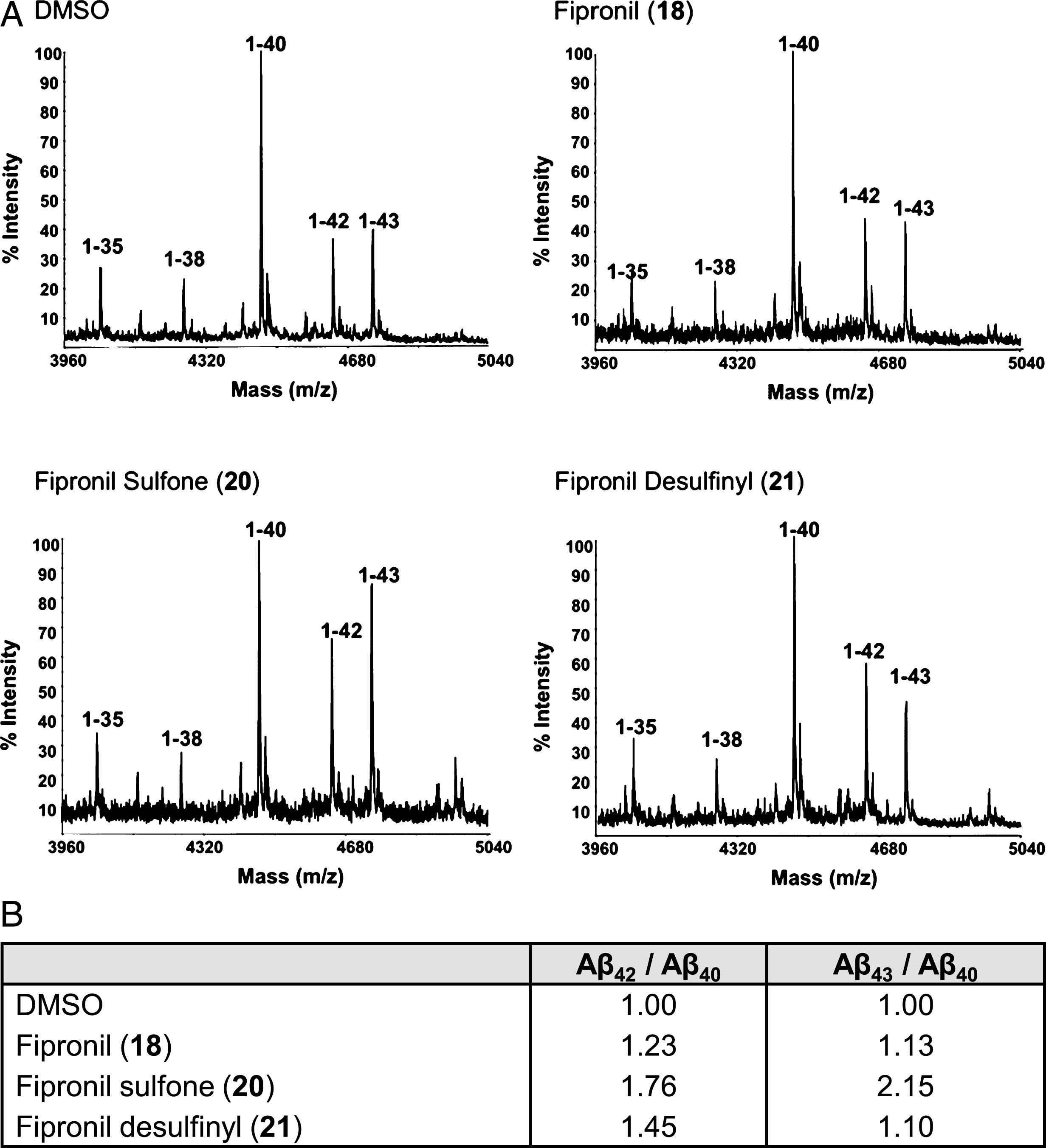
Mass spectrometric analysis of the Aβs generated in cell-free γ-secretase assays, in the presence of DMSO or fipronils
Pyrazoles shift the cleavage pattern of another γ-secretase substrate, alcadeinα
Like AβPP, alcadeins (Alcs) are sequentially cleaved by secretases, first by α-secretase, leading to N- and C-terminal fragments, the latter being then cleaved by γ-secretase to an intracellular domain and the p3-Alcs peptide, in a way similar to AβPP [61, 62] (see Fig. 6A in [38]). HEK293 cells stably expressing full length alcadeinα were used to investigate the effects of pyrazoles on alcadein cleavage. Alcadeinα is first cleaved on the N-terminal side (two possible sites) followed by cleavage by γ-secretase leading to p3-Alcα35 and p3-Alcα 2N+35, the latter representing the major peptide in cultured cells. HEK293-alcadeinα cells were grown till 70% confluence and treated with 100μM pyrazoles for 48 h. The secreted p3-Alcα peptides were recovered by immunoprecipitation and analyzed by MALDI TOF/MS (Fig. 9A). Quantification of p3-Alcα peptides showed that, compared to the p3-Alcα peptide profile in vehicle treated cells, concentrations of the main alcadeinα peptide (p3-Alcα2N+35) and p3-Alcα2N+37 peptide remained stable. In contrast, both p3-Alcα2N+34 and p3-Alcα2N+36 concentrations dropped significantly while the level of p p3-Alcα2N+38 peptide concentration strongly increased (Fig. 9B). These results show that, like for AβPP, pyrazoles induce a shift in the cleavage pattern of Alcadeinα, another γ-secretase substrate. Similar effects were seen previously with aftin-5 and triazines [38]. These observations further suggest that pyrazoles are more likely to interact with γ-secretase than with their substrates.
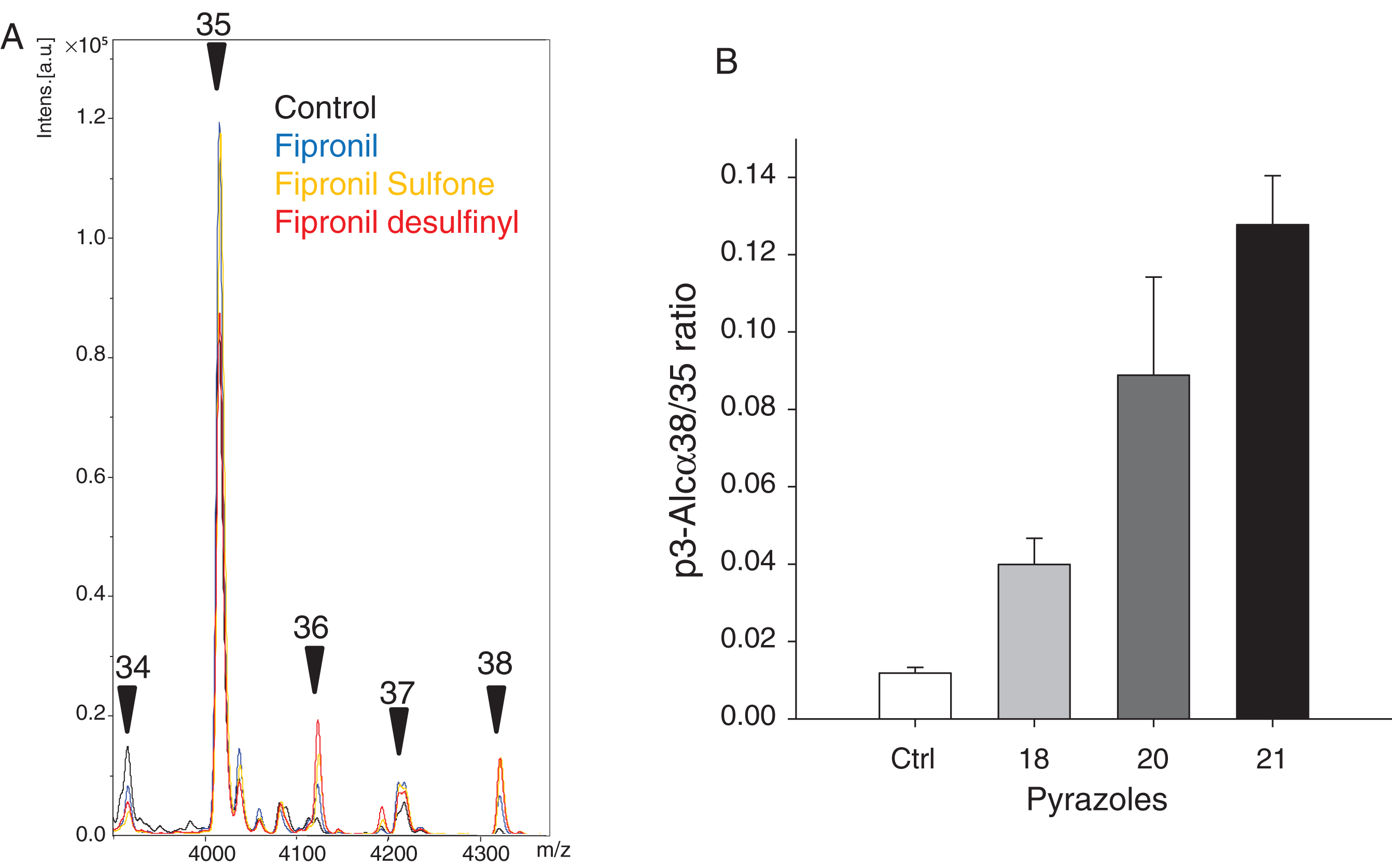
Pyrazoles alter the cleavage pattern of Alcadeinα, leading to increased p3-Alcα38 production. A) Immunoprecipitation/mass spectrometry spectra of p3-Alcα peptides produced by HEK cells expressing full length Alcadeinα exposed to various pyrazoles or DMSO (control). Cells were treated for 48 h with 100μM of each reagent and p3-Alc peptides were analyzed by MALDI-TOF/MS. A) Representative profile for each product showing the the p3-Alcα2N+34, p3-Alcα2N+35, p3-Alcα2N+36, p3-Alcα2N+37 and p3-Alcα2N+38 peaks. B) Relative quantification of p3-Alcα peptides produced by cells exposed to DMSO (control), fipronil (
Proteomics study of fipronil’s effects
In order to understand the molecular mechanisms of action of pyrazoles, we decided to analyze protein expression of cells exposed to fipronil sulfone (

Proteomics and phosphoproteomics analysis of N2a-APP695 cells exposed to fipronil sulfone
Fipronil is metabolized to fipronil sulfone, a stable metabolite which accumulates in adipose tissue and brain
We next investigated the pharmacokinetics and biodistribution of fipronil (
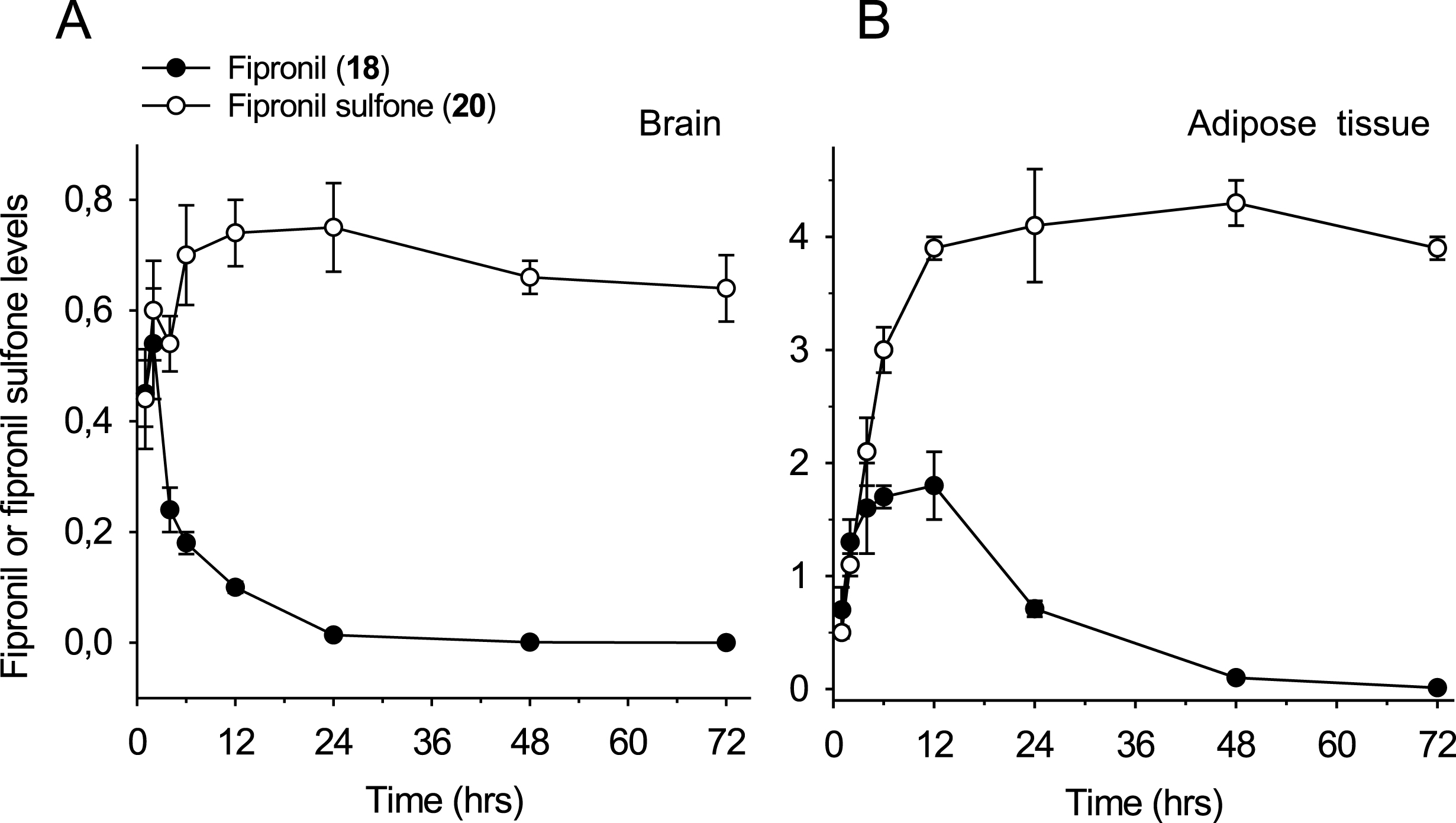
Short-term (72 hrs) time-course of fipronil (18) and fipronil sulfone (20) bioaccumulation in brain (A) and epididymal adipose tissue (B) following a single oral administration of fipronil

Long-term (56 days) time-course of fipronil sulfone (20) production and accumulation following single (A) or repeated (B) oral administrations of fipronil
DISCUSSION
This study reports on the induction of Aβ42/Aβ43 production, and the increased ratio of long versus short Aβs, in various cell cultures and by highly purified γ-secretase following exposure to several pyrazole insecticides. These results support previous results obtained with various drugs (fenofibrate, celecoxib, indomethacin, isoprenoids) [63], DAPT under certain conditions [64, 65], steroids [66], ceramide analogs [67], the peroxynitrite donor SIN-1 [68], Zinc [60], aftins [35–37] and several triazine herbicides [38]. Altogether these results have two main implications: They support the idea that the HCE contains several compounds able to shift the cleavage of AβPP toward the production of long, toxic, aggregation-prone Aβs, such as Aβ42/Aβ43, those which are classically associated with AD in humans and numerous animal models of AD. These compounds, if proven to modify this balance in animals (as suggested by the first preliminary results obtained with aftin-4 [28] and celecoxib or FT-1 [63]) and in human, may contribute to the onset, development and/or acceleration of AD. We suggest that they be collectively named “Alzheimerogens”, by analogy with cancer-inducing carcinogens. Identification of such compounds in our environment appears to be a priority to develop a prevention strategy. Some of these compounds, particularly the metabolically stable, brain-permeable molecules, constitute new pharmacological research tools to investigate the role of γ-secretase, its substrates and products, in the onset of AD, in cell lines and animal models. In this context, one of our objectives is, by analogy with MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine)-induced Parkinsonism, to develop a pesticide-induced animal model of AD, which would allow the study of AD onset in wild-type (WT) animals. This is exemplified by the orally available fipronil (
Mechanism of action of fipronil and other pyrazoles
As also observed with aftins [36] and triazines [38], pyrazoles insecticides, including fipronil derivatives, show a structure/activity relationship (Supplementary Table 1) in their effects on Aβ production – some are active, others are inactive-, showing that pyrazole insecticides devoid of Aβ42 induction properties can be developed. The limited number of pyrazoles and their wide structural diversity precludes a clear identification of key structural features interacting with the unknown target leading to Aβ42 production. Unspecific, detergent, hydrophobic, membrane, or protein structure disrupting actions are unlikely to account for the effects of pyrazoles. Several arguments support an effect on γ-secretase and/or its micro-environment: 1) induction of Aβ production by pyrazoles is inhibited by γ-secretase inhibitors or modulators (Fig. 4); 2) pyrazoles also affect the cleavage of another substrate of γ-secretase, alcadeins (Fig. 9); and 3) pyrazoles modify the cleavage pattern of AβPP by highly purified γ-secretase.
Our cell-free γ-secretase activity assays performed with highly purified enzyme and substrate (Fig. 8) show that fipronil (
Further quantitative experiments are required to explain the increased Aβ42/Aβ40 or Aβ43/Aβ40 ratios, caused either by increased Aβ42 or Aβ43 production, a reduced Aβ40 production or both. In any case, an altered Aβ profile in the presence of pyrazole insecticides strongly suggests that the compounds directly affect the positioning of the AβPP substrate in the lipid-bilayer, as previously observed for compounds known as inverse γ-secretase modulators (iGSM) [64]. Similarly to what we observed with fipronil (
Fipronil exposure and consequences
Fipronil has been developed a selective inhibitor of insect GABA-gated chloride channels (review in [44]). It is one of the most widely used pyrazole insecticides in agriculture (seed, culture, and crop treatment), in urban pest control (coackroaches, termites, wasps, flies, and ants management in and around buildings), and in veterinary applications (fleas and ticks control for pets and cattle). Fipronil and its metabolites are therefore widely found [73] in urban soil, dust, wastewater, and residential runoff [74], in rural soil, rivers, and atmosphere [46, 76], but also in various tissues of pet, farmland, and wild-life animals [75, 78] (only a very small number of references are cited). Although fipronil (
As shown in Figs. 11 and 12, fipronil (
In conclusion, we have added some pyrazole insecticides, especially the main metabolite of fipronil, in the growing list of HCE products which are able to alter the specificity of Aβ production. Our hypothesis is that these products may, on a long-term, cumulative and additive basis, possibly in parallel with microorganisms [101] and microorganisms-derived products, alter the Aβ production pattern sufficiently to lead to AD pathogenesis, and thus qualify as potential “Alzheimerogens”. Gradual accumulation in and chronic release from adipose tissue and transfer to the brain may contribute to the onset, development, and acceleration of sporadic AD.
Footnotes
ACKNOWLEDGMENTS
We are grateful to Laetitia Bailly (Laboratoire Cobra, University of Rouen) for purifying the two fipronil enantiomers. We are grateful to Jasna Padovan and Zeljko Javorscak (Fidelta) for the rat PK studies, to Rebecca L. McMahen, Mark J. Strynar for providing fipronil sulfone chloramine. This work was supported by ‘Fonds Unique Interministériel’ PHARMASEA/TRIAD projects, « Agence nationale de sécurité sanitaire, de l’alimentation, de l’environnement et du travail » (ANSES), Fondation “Jérôme Lejeune” and CRITT-Santé Bretagne /FEDER (LM). This research was also partly supported by an FP7-KBBE-2012 grant (BlueGenics) to LM. MC is CIFRE/ManRos Therapeutics PhD fellowship recipient. KB is a Torsten Söderberg Professor. HZ is a Wallenberg Academy Fellow. This work was supported in part by Grant-in-aid for Scientific Research 26293010 (TS) and 24790062 (SH) from the Ministry of Education, Culture, Sports, Science and Technology in Japan. HG, JF, and PCF were supported by the Fondation Eclosion. Partial support was provided by NIEH S/ROI ES002710, NIEH S Superfund Research Program P42 ES004699 and NIH/454 NS079202.
Although fipronil is not the most potent Aβ42 inducer, it was selected for more detailed studies, among other pyrazoles, for several reasons: 1) its very wide use all over the world (despite its ban in Europe in America for food plant cultures), 2) its wide use against fleas and ticks in domestic animals, 3) its wide presence in urban and rural environment, 4) its chemical stability and environmental persistence (especially that of its metabolite, fipronil sulfone), 5) the biological activity of the main metabolite, 6) the fact fipronil sulfone accumulates in adipose tissue, crosses the blood-brain barrier and accumulates in the brain, and 8) its availability in large quantities for very cheap prices (in preparation of our current long term exposure experiments the supply issue needed to be solved). Incidentally, fipronil was selected before the public health scandal of summer 2017 in Europe (contamination of eggs by fipronil, presumably even more by fipronil sulfone)!