
Abstract
Alzheimer’s disease (AD) is characterized by neuritic plaques and neurofibrillary tangles. It is reported that enzymatic degradation of amyloid-β (Aβ) plays a pivotal role in Aβ accumulation and type-2 cannabinoid receptor (CB2R) participates in Aβ processing in the brain; however, the underlying mechanisms remain unclear. We determined that Aβ degradation-related proteins are significantly different between CB2R–/– mice and wild-type (WT) mice via proteomic analysis. Moreover, the data demonstrated that the angiotensin converting enzyme (ACE) and insulin-degrading enzyme (IDE) levels are substantially attenuated, and the Aβ level is significantly enhanced in CB2R–/–-Aβ1 - 42 mice compared with that of WT-Aβ1 - 42 mice. Furthermore, Aβ-mediated synaptic dysfunction, the loss of memory associated proteins, and the suppression of glutamatergic transmission are more severe in CB2R–/–-Aβ1 - 42 mice than that in WT-Aβ1 - 42 mice. CB2R activation could decrease Aβ1 - 40 and Aβ1 - 42 levels and enhance ACE and IDE levels with its selective agonist JWH133; however, AM630 (CB2R antagonist) abrogates all changes induced by JWH133 in N2a cells with AβPP overexpression. Taken together, our study demonstrated that the deletion of CB2R reduces exogenous Aβ degradation and aggravates the toxicity of Aβ via the reduction of ACE and IDE, which suggests that CB2R is involved in the onset of AD and a potential therapeutic target for AD.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is a progressive neurodegenerative disease, which is characterized by the amyloid-β (Aβ) aggregates formation of neuritic plaques [1] and neurofibrillary tangles composed of hyperphosphorylated tau protein [2]. The Aβ accumulation results from an imbalance between Aβ production and Aβ degradation and clearance in AD progression [3]. Aβ aggregation leads to neuronal cell loss and synaptic dysfunction [4]. Aβ degradation and clearance play a pivotal role in Aβ accumulation [5, 6], the mechanisms of which include non-enzymatic and enzymatic pathways. The non-enzymatic pathway contain 1) receptor-mediated clearance of Aβ, such as low-density lipoprotein receptor-related protein (LRP) [7, 8], and receptor for advanced glycation end products (RAGE) [9], 2) the uptake and phagocytosis of Aβ by cells such as microglia, astrocytes and macrophages [10], and 3) the autophagy pathway, which is critical for Aβ degradation [11]. Furthermore, enzymes-mediated-degradation and clearance of Aβ are composed of numerous enzymes including but not limited to, angiotensin converting enzyme (ACE) [12], insulin-degrading enzyme (IDE) [13, 14], and neprilysin (NEP) [13].
Two cannabinoid receptors (CB1R and CB2R) have been identified and recognized as key members of endocannabinoid signaling. CB1R is the most abundant cannabinoid receptor in the brain [15]; however, the activation of CB1R is accompanied by psychoactive effects [16]. In contrast, CB2R has none of these side effects. CB2R was initially identified in the immune system [17], whereas accumulating evidence has demonstrated that CB2R is also present in the central nervous system, such as the hippocampus, cerebral cortex [18, 19], ventral tegmental area [20], nucleus accumbens, dorsal striatum [21], and cerebellum [22]. The roles of CB2R in anxiety, schizophrenia, drug addiction, depression, and Huntington’s disease have been demonstrated in different laboratories [23–26], suggesting that CB2R is involved in the regulation of brain function. Evidence has suggested a direct role of CB2R in the modulation of Aβ peptide levels in brain [27–30]. For example, MDA7 (CB2R agonist) reverses Aβ-induced memory deficiency via promoting Aβ clearance [27]; the soluble Aβ1 - 42 and plaque deposition are significantly increased in J20 CB2R–/– mice relative to J20 CB2R +/+ mice [29]. Moreover, a lack of CB2R exacerbates the cortical Aβ deposition and increases the levels of soluble Aβ1 - 40 in APP/PS1 mice [28]. Taken together, previous studies have suggested that CB2R plays a crucial role in the process of Aβ degradation. However, the mechanism of CB2R regulating Aβ degradation remains elusive.
To investigate the roles of CB2R in Aβ degradation and its underlying mechanism, we established exogenous Aβ-induced AD-like models in CB2R–/– or wild-type (WT) mice. In this study, we determined that the levels of Aβ are substantially increased and the levels of ACE and IDE are significantly decreased in CB2R–/–-Aβ mice compared with those in WT-Aβ mice. Furthermore, CB2R–/–-Aβ mice display a more severe response to Aβ neurotoxicity than that of WT-Aβ mice. We also showed that CB2R inactivation could abrogate the CB2R activation induced Aβ reduction and augmentation of ACE and IDEin vitro.
MATERIALS AND METHODS
Animals
The generation of mice that lack CB2R has been previously described (PMID: 10822068). The male CB2R–/– mice on a C57BL/6 congenic background were kindly provided by Nancy E. Buckley (Cal State Polytechnic University, Pomona, CA). Non-transgenic WT mice and CB2R–/– mice were housed in cages (4-5 mice per cage), received food (standard chow) and water ad libitum, and were maintained on a reverse 12-h light/12-h dark cycle with lights on at 07:00 am and off at 07:00 pm, at 24±2°C room temperature. In all experiments, the mice used were matched for age (2.5–3 months old) and weight (25–30 g). All experiments were approved by the Institutional Ethics Committee of Tongji Medical College, Hua-Zhong University of Science and Technology, and were conducted in accordance with the “Policies on the Use of Animals and Humans in Neuroscience Research” revised, approved by the Society for Neuroscience in 1995.
Aβ1 - 42 peptide preparation
Aβ1 - 42 peptide were purchased from Sigma (St Louis, MO, USA, A9810, SCP0048) and oligomerized by the procedure described previously [31]. In brief, the Aβ1 - 42 peptides were dissolved in dimethyl sulfoxide (DMSO) and diluted in sterile physiological saline to a final concentration of 1.0 mg/ml and 1% (vol/vol) DMSO. The solution was subsequently incubated for oligomerization at 37°C in the dark for 1 week prior to use. The Aβ1 - 42 oligomers were confirmed by western blot.
Intracerebroventricular surgery
The CB2R–/– mice at the age of 3 months and 3-month-old WT mice were positioned in a stereotaxic instrument; then, 5μl of the solution with Aβ1 - 42 or without Aβ1 - 42 were infused into the left lateral ventricle (anteroposterior, –0.3 mm; mediolateral, –1.0 mm; dorsoventral, –2.5 mm; relative to the bregma) at a rate of 0.2μl/min. The needle was maintained in place for 10 min prior to being withdrawn to avoid flow back of the solution.
Antibodies
An overview of all antibodies employed in this study is given in Table 2.
Cell cultures and treatments
N2a cells were cultured in DMEM with 5% heat-inactivated fetal bovine serum and were incubated at 37°C in a humidified atmosphere with 5% CO2. For AβPP overexpression, N2a cells were transfected with plasmids that encoded Myc-tagged human AβPP (generously provided by Dr. Yizheng Wang) for 48 h. The transfection was conducted according to the manufacturer’s instructions. In brief, the plasmids (5μg) were mixed with Lipofectamine-2000 (5μl) in 250μl OPTI-MEM followed by incubation at room temperature for 25 min; the Lipofectamine-DNA complex was subsequently added to the N2a cells. Following the administration of CB2R agonist (JWH133) or united with CB2R antagonist (AM630), the cells were incubated at 37°C for 24 h. Cells were harvested for furtheranalysis.
Measurement of Aβ, ACE, and IDE levels
ELISA kits for Aβ1 - 40 and human Aβ1 - 42 (Elabscience Biotechnology Co., Ltd, catalog NO: E-EL-M0067c, E-EL-H0542c and E-EL- H0543c) were used to detect the Aβ1 - 40 or Aβ1 - 42 levels. ELISA kits were employed for mouse ACE and IDE (MLBIO Biotechnology Co., Ltd, catalog #SU-B20735 and SU-B20779). In brief, the hippocampus of the mice was homogenized in a 9×mass of cold RIPA buffer (which contained 50 mM Tris, pH 7.4;150 mM NaCl; 1% Triton X-100; 1% sodium deoxycholate; 0.1% SDS,1 mM phenylmethylsulfonylfluoride, and 10μg/ml protease inhibitor cocktail) for ELISA. The supernatants were collected after centrifugation (12,000×rpm for 10 min at 4°C). The cell lysate was prepared to assess the levels of Aβ1 - 40, Aβ1 - 42, ACE and IDE with ELISA. The homogenates were subjected to ELISA analysis using the kit according to the manufacturer’s instructions.
Western blot
Western blot was performed as described previously [32]. In brief, the dissected hippocampal tissue and the cell extracts were homogenized in RIPA buffer (which contained 50 mM Tris, pH 7.4, 150 mM NaCl, 1% Triton X-100, 1% sodium deoxycholate, 0.1% SDS, 1 mM phenylmethylsulfonylfluoride, and 10μg /ml protease inhibitor cocktail), followed by boiling for 10 min and ultrasonic waving for 10 times (60 Hz), after centrifugation at 12,000×rpm for 10 min, the supernatants were collected and the protein levels were analyzed. The proteins in the extracts were separated by 8–12% SDS-PAGE and transferred to nitrocellulose membranes. The membranes were subsequently blocked in 5% non-fat milk for 30 min at room temperature and incubated with primary antibody at 4°C overnight. The blots were then incubated with anti-rabbit or anti-mouse IgG conjugated to IRDyetrademark (800CW) (1:10000) for 1 h at room temperature. Immunoreactive bands were visualized using the Odyssey Infrared Imaging System (Licor biosciences, Lincoln,NE, USA).
Immunohistochemistry
Mice were anesthetized and perfused through the aorta with 300 ml normal saline, followed by perfusion with 4% paraformaldehyde (PFA) in 10 mM phosphate-buffered saline (PBS). The brain was removed and post-fixated in the same paraformaldehyde for an additional 48 h. Coronal slices (30μm) were subsequently cut with a freezing microtome. Immunohistochemistry was performed as previously described [33]. The sections were blocked with 0.3% H2O2 that contained 0.5% Triton X-100 for 30 min; nonspecific sites were blocked with 5% bovine serum albumin for 60 min at room temperature and incubated with primary antibodies overnight at 4°C. After washing with phosphate-buffered saline, the sections were incubated with biotin-labeled secondary antibodies for 1 h at 37°C. The immunoreaction was detected using horseradish peroxidase labeled antibodies for 1 h at 37°C and was visualized with the DAB tetrachloride system for brown color. The images were observed using a microscope (Olympus BX60, Tokyo, Japan) at identical settings for each of the conditions. Image analysis was performed as described in a previousstudy [34].
Morris water maze test
The Morris water maze (MWM) test was used to assess the spatial learning and memory. The water maze (180-cm diameter, 60-cm high) was filled with water (22±2°C) and made opaque by the addition of milk powder. For spatial learning, one week after the injection of Aβ1 - 42, the mice were trained in the water maze to find a hidden platform for 6 consecutive days, 4 trials per day with a 2 h interval from 14:00 to 22:00 pm. On each trial, the mice started from one of the four quadrants facing the wall of the pool and ended when the animal climbed on the platform. If the mice did not locate the platform within 60 s, they were guided to the platform and remained on the platform for 30 s uniformly. The swimming paths and the time used to find the platform (latency) were recorded each day. The spatial memory was tested on the 7th day. The platform was removed, and each mouse was allowed to swim freely for 60 s; the platform crossings, the time in the target quadrant, and the swimming paths were recorded.
Electrophysiology
Mice were decapitated, and the brains were immediately removed and immersed in ice-cold artificial cerebrospinal fluid (ACSF, 126 mM NaCl, 2.5 mM KCl, 1.2 mM MgSO4, 2.0 mM CaCl2, 1.25 mM NaH2PO4, 10 mM glucose, and 26 mM NaHCO3), which was continuously bubbled with 95% O2 and 5% CO2. Parasagittal sections (300 mm) were cut using a vibrating microtome (Leica). Slices were incubated in oxygenated ACSF at room temperature for at least 2 h for recovery. A planar multi-electrode recording setup (MED64 system, Alpha Med Sciences) was used to record the fEPSPs. Each slice was superfused with 100 ml oxygenated artificial cerebrospinal fluid, which was recirculated at a flow rate of 1.0–1.5 ml/min. The fEPSPs were recorded from the dendritic layer of CA1 neurons by choosing an electrode in the Schaffer collateral pathway as the stimulating electrode. After establishing a stable baseline of 30 min, an induction protocol that evoked long-term potentiation was induced by applying 1 train of high frequency stimulation (100 Hz, 1 s duration at test strength). The field potential was monitored for at least 1 hour after the conditioning stimuli. The values obtained after HFS were expressed as percentages of their respective baseline recordings, and the last 20-min blocks of recordings were analyzed.
Proteomic analysis
Hippocampal proteins were extracted and concentrated using a 2-D Quant kit (GE Healthcare) according to the manufacturer’s instructions, with trypsin digestion. The obtained peptides were subsequently labeled by iTRAQ-6plex reagents on the basis of the manufacturer’s protocol [35]. The efficiency of iTRAQ labeling was greater than 95%. The labeled peptides were subsequently fractionated into fractions by high pH reverse-phase HPLC. For the LC-MS/MS analysis, the peptides were dissolved in 0.1% FA and directly loaded onto a reversed-phase pre-column (Acclaim PepMap 100, Thermo Scientific); the peptides separation was subsequently performed by a reversed-phase analytical column (Acclaim PepMap RSLC, Thermo Scientific). The obtained peptides were analyzed by a Q ExactiveTM hybrid quadrupole-Orbitrap mass spectrometer (ThermoFisher Scientific). To identify and quantify proteins, the resulting MS/MS data were processed using Mascot search engine (v.2.3.0) and tandem mass spectra were searched against the Swissprot mouse database (16,717 sequences). We defined observed proteins with iTRAQ ratios of >1.2 and <0.83 as differentially expressed.
Bioinformatics analysis
To investigate the proteins with differential expression, gene ontology (GO) annotations were used, which covers three categories: cellular compartment, molecular function and biological process. For each category, a two-tailed Fisher’s exact test was employed to test the enrichment of the differential expression protein against all identified proteins. Correction for multiple hypothesis testing was performed using standard false discovery rate control methods. A GO with a corrected p-value <0.05 is considered significant. The GO annotation proteome originated from the UniProt-GOA database (http://www. http://www.ebi.ac.uk/GOA/). First, the identified protein ID was converted to the UniProt ID and then mapped to GO IDs by protein ID. If identified proteins were not annotated by the UniProt-GOA database, the InterProScan software was used to annotated the protein’s GO function based on the protein sequence alignment method. Proteins were then classified by Gene Ontology annotation based on three categories: biological process, cellular component and molecular function.
Statistical analysis
All data were expressed as mean±standard error of the mean and analyzed using the SPSS 17.0 statistical software (SPSS Inc, Chicago, IL, USA) and the statistical graphs were produced by GraphPad Prism6 (GraphPad Software, Inc., La Jolla, CA). For two conditions, the significance of data was assessed by the 2-tailed student unpaired t test. For comparison of multiple groups, the one-way ANOVA of variance or two-way analysis of variance procedure followed by post hoc Tukey’s test. The test with p < 0.05 was considered as statistically significant. All experiments were performed at least threetimes.
RESULTS
Differentially expressed proteins and functional classification of these proteins in hippocampus of CB2R–/– and WT mice
Previous studies have shown that CB2R–/– mice display AD-associated pathological alternations and cognitive disorder [36, 37]; however, proteomic data in the hippocampus of CB2R–/– mice have not been previously reported. Data regarding differentially expressed proteins and functional classification of these proteins may provide a crucial clue for exploring the CB2R role and molecular mechanism in the onset of AD. Thus, we initially measured the protein expression in the hippocampus of CB2R–/– and WT mice with proteomic analysis. The results showed that 4,704 proteins were identified and 2,949 proteins were quantified in total. When a quantification ratio of >1.2 was set as the upregulated threshold and <0.83 was set as the downregulated threshold, we determined that there are thirty-seven differentially expressed proteins, including 12 cytosol, 15 nuclear, 7 extracellular, 3 plasma-membrane, and 2 mitochondrial proteins, between the CB2R–/–and WT mice using UniProt and DAVID database analysis, however, only eighteen proteins were significantly different (p < 0.05) between the CB2R–/– and WT mice (Table 1).
Differentially expressed proteins present in WT and CB2R–/– mice
*p < 0.05, **p < 0.01, ***p < 0.001; extr, extracellular; nucl, nuclear; cyto, cytosol; plas, plasma membrane; mito, mitochondria.
Antibodies employed in this study
To further understand the function and feature of the differential proteins, their biological processes and signal transduction pathways were examined. The identified proteins were classified into 11 biological process categories on the basis of their GO functional annotations, including cellular processes, single-organism processes, metabolic processes, biological regulation, response to stimulus, localization, cellular component organization or biogenesis, multicellular organismal processes, developmental processes, signaling and other mechanisms. According to the proteomic data, we verified that most of these differential proteins were associated with cellular process, metabolic process and the response to stimuli (Fig. 1a), which may relate to Aβ degradation and cause aggregation of Aβ in CB2R–/– mice. The detailed signal transduction pathways related to these proteins are shown in Fig. 1b.
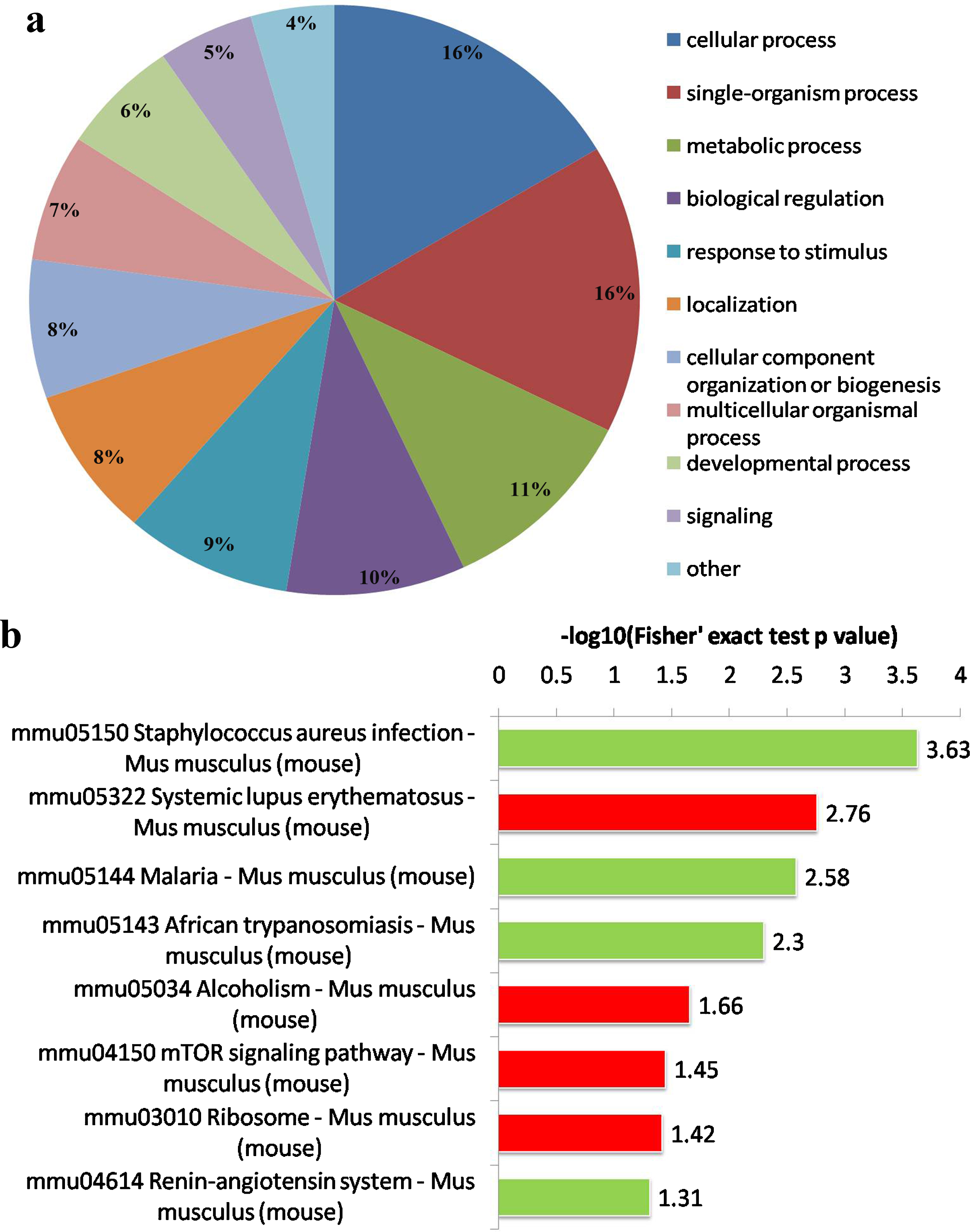
Functional classification of the differentially expressed proteins in the hippocampus between CB2R–/– and WT mice. a) Pie charts of functional classification of differentially expressed proteins. The 11 biological processes in a ranking from gene ontology (GO). Percentage of the number of proteins associated with each category is presented. b) The differentially expressed proteins are associated with signal transduction pathways. The red and green color codes represent the remarkable up-regulation and down-regulation signal pathways respectively.
The levels of Aβ are enhanced and levels of ACE and IDE are decreased in CB2R–/– -Aβ mice
The bioinformatic analysis of proteomics showed that the levels of Aβ degradation-related proteins are significantly altered between the WT and CB2R–/– mice; however, whether these protein alternations result in the dysfunction of Aβ degradation must be verified. The Aβ mouse model induced by ICV-injection oligomers Aβ1 - 42 has been broadly accepted to investigate the Aβ pathology and mechanism in AD; this model is a simple tool used to evaluate the effect of CB2R on Aβ degradation and excludes other potential factors, such as multiple transgenic models that result in proteomic changes [38]. The western blot image showed that the soluble Aβ1 - 42 had formed the oligomers Aβ1 - 42 (Fig. 2a), which were injected into the left lateral ventricle to establish the Aβ mouse model. Immune staining of 6E10 was substantially stronger in the mice injected with oligomers Aβ1 - 42 than in the control mice injected with the solvent of Aβ1 - 42 (1% DMSO); moreover, the staining of 6E10 in the CB2R–/–-Aβ mice was significantly enhanced compared with that in the WT-Aβ mice (Fig. 2b, c). To further verify the content of Aβ, we detected the levels of Aβ1 - 40/1 -42 using ELISA; the Aβ1 - 40 and Aβ1 - 42 levels were not different between the WT mice and CB2R–/– mice (Fig. 2d, e). The contents of Aβ1 - 42 in the CB2R–/–-Aβ mice were also significantly enhanced compared with those in the mice injected with the solvent of Aβ1 - 42; moreover, the levels of Aβ1 - 42 were higher in the CB2R–/–-Aβ mice than in the WT-Aβ mice (Fig. 2e). The levels of Aβ1 - 40, which represent endogenous Aβ metabolism, had a tendency to increase; however, there was no statistical significance (Fig. 2d).

The levels of Aβ were increased in CB2R–/–-Aβ mice compared with those in WT-Aβ mice. After performing the behavior test, the mice were sacrificed, and extracts of hippocampal protein were prepared to measure the levels of Aβ1 - 40 and Aβ1 - 42 with ELISA or immunohistochemistry. a) Monomeric (mono) or oligomerized (oligo) synthetic human Aβ1 - 42 peptides were analyzed by western blot with antibody to Aβ (6E10). b, c) Representative immunostaining images with 6E10 and quantitative analysis. Scale bars: 500μm. d) The levels of Aβ1 - 40 were measured by ELISA, mouse Aβ40 (E-EL-M0067c) ELISA kit was used. e) The levels of Aβ1 - 42 were measured by ELISA, human Aβ42 (E-EL- H0543c) ELISA kit was used. N = 6 mice per group for ELISA, N = 3 mice for each group for immunohistochemistry, one-way ANOVA with a post hoc Tukey’s test, *p < 0.05; ***p < 0.001 versus WT mice; # p < 0.05 versus WT-Aβ mice. Data were expressed as mean±SEM.
Our bioinformatic analysis only displayed a reduction of ACE expression; however, we considered the limitation of the proteomics analysis and the previous studies in Aβ enzymatic-degradation-protein [39, 40], and we detected the expression of ACE and IDE. The levels of ACE were notably decreased in the CB2R–/–-Aβ mice compared with those in the WT-Aβ mice (Fig. 3a), which confirmed the results obtained in the proteomic analysis (Table 1). The change of IDE was similar to that of ACE (Fig. 3b), however, the change was not displayed in the proteomic analysis (Table 1). These data suggested that the Aβ mouse models were successfully established via the ICV-injection oligomers Aβ1 - 42, and the CB2R knockout should be responsible for the reduction of ACE and IDE and more exogenous Aβ accumulation in the CB2R–/–-Aβ mice; these findings suggest that the impairment of Aβ degradation is associated with the reduction of ACE and IDE in CB2R–/– mice.
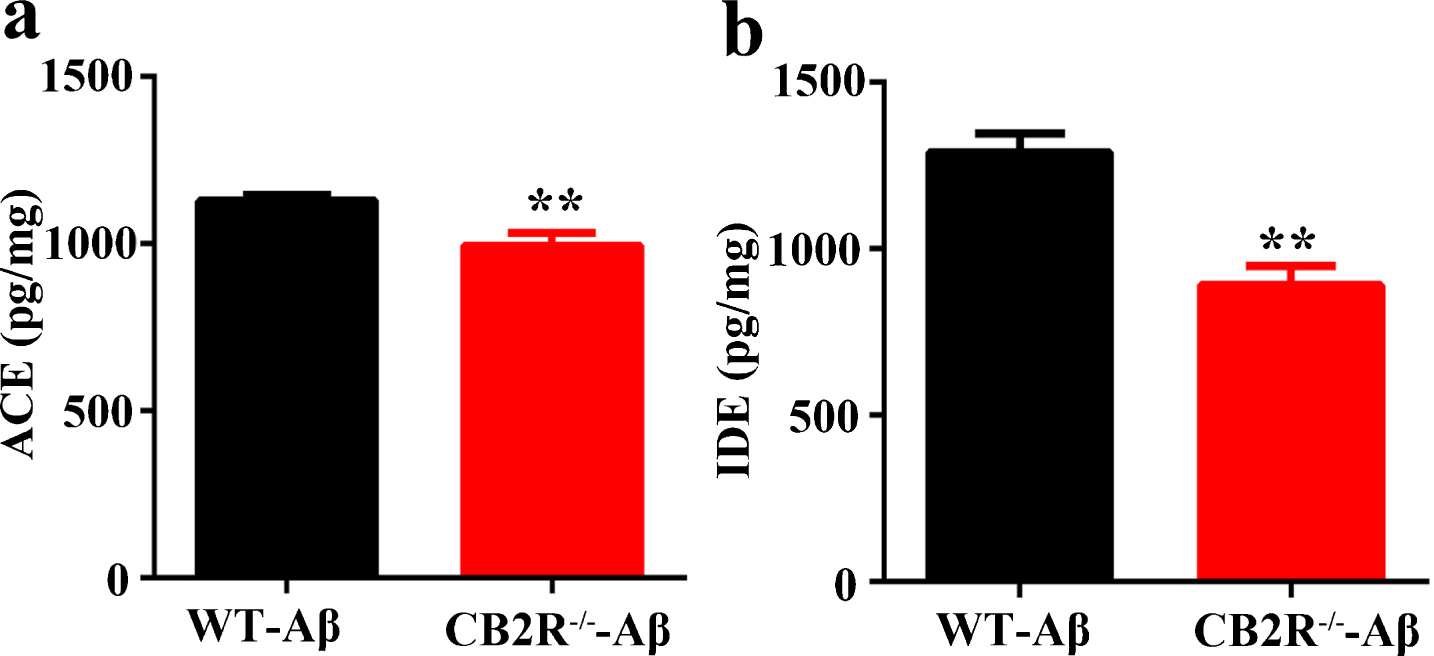
The levels of ACE and IDE were reduced in CB2R–/–-Aβ mice compared with those in WT-Aβ mice. After performing the behavior test, the mice were sacrificed, and extracts of hippocampal protein were prepared to measure the levels of ACE and IDE with ELISA. a) The levels of ACE were measured by ELISA in WT-Aβ mice and CB2R–/–-Aβ mice. b) The levels of IDE were detected by ELISA in WT-Aβ mice and CB2R–/–-Aβ mice. N = 6 mice per group for ELISA, student unpaired t test, **p < 0.01 versus WT-Aβ mice. Data were expressed as mean±SEM.
Lack of CB2R exacerbates exogenous Aβ-induced synapse impairments
To explore the underlying mechanisms of behavioral impairment, we further measured the synaptic transmission in the hippocampal CA1 regions via the long-term potentiation (LTP), the activity-dependent change in synaptic plasticity, which is related to learning and memory [42, 43], by recording the fEPSPs after tetanic stimulation. We determined that the slope of the fEPSPs in the CB2R–/–-Aβ mice was significantly decreased compared with that in the WT-Aβ mice (Fig. 5a, b). Dendritic spine density plays a crucial role in cognitive function and synaptic plasticity. We subsequently examined the dendritic spine structure in the CB2R–/–-Aβ mice and WT-Aβ mice via Golgi staining. We identified a decreased dendritic spine density in the hippocampal CA1 regions of the CB2R–/–-Aβ mice compared with that of the WT-Aβ mice (Fig. 5c, d). Furthermore, we measured the levels of synapse-associated proteins, and our data showed that the levels of postsynaptic density-93 (PSD93) and postsynaptic density-95 (PSD95) (Fig. 6a, b), as well as N-methyl-D-aspartate receptor type 2A (NR2A), N-methyl-D-aspartate receptor type 2B (NR2B) and N-methyl-D-aspartate receptor subunit NR1 (NR1) (Fig. 6a, c), were reduced in the CB2R–/–-Aβ mice, compared with those in the WT-Aβ mice; however, the presynaptic proteins (synapsin I, synaptophysin) (Fig. 6a, b), AMPA-selective glutamate receptor 1 (GluR1) and AMPA-selective glutamate receptor (GluR2) (Fig. 6a, c) were not significantly different between the CB2R–/–-Aβ mice and WT-Aβ mice. Although previous studies have shown that Aβ-induced LTP impairments always accompany synaptic proteins changes in presynaptic and postsynaptic proteins, we showed that only postsynaptic proteins exhibit clear changes between CB2R–/– and WT mice injected with Aβ1 - 42, which suggests that postsynaptic proteins are more crucial for exogenous Aβ-induced impairments than presynatic proteins in CB2R–/– mice.
CB2R–/–-Aβ mice display severe impairment of hippocampus-dependent spatial memory
We used the MWM test to evaluate the spatial learning and memory ability of all experimental mice; this test is considered a canonical approach to measure the capacity of hippocampus-dependent spatial learning and memory in rodents [41]. We first assessed the ability of learning and memory in 3-month-old WT mice and CB2R–/–mice. There was no difference in learning and memory between these two groups of mice in the MWM test (Fig. 4a-e). It is established that exogenous Aβ leads to the impairment of hippocampus-dependent spatial memory in mice. Thus, we evaluated whether more accumulation of Aβ induces more severe impairments of brain function in CB2R–/–-Aβ mice than in WT-Aβ mice. In the MWM test, the learning ability was evaluated using 6-day consecutive training and learning, and the CB2R–/–-Aβ mice spent a longer time to find the hidden platform on the 5th and 6th days than the WT-Abeta mice (Fig. 4a). The memory retention ability was assessed by the hidden platform trials on day 7. Compared with the WT-Aβ mice, the CB2R–/–-Aβ mice displayed a curved line toward the platform (Fig. 4b), stayed a shorter time in the target quadrant (Fig. 4d) and decreased the target platform crossings (Fig. 4e). The swimming speed was not different between the WT-Aβ mice and CB2R–/–-Aβ mice (Fig. 4c); thus, we considered that the behavior change is not associated with locomotor activity in CB2R–/–-Aβ mice. Taken together, the results of the behavioral assay supported that a lack of Aβ degradation could reinforce exogenous Aβ-caused brain dysfunction in CB2R–/–-Aβmice.

The hippocampus-dependent spatial memory is impaired in CB2R–/–-Aβ mice. After microinjection of Aβ1 - 42 in the left lateral ventricle of mice for 1 week, the spatial learning and memory ability were measured with the MWM test. In the MWM test, mice underwent a 6-day consecutive training to measure the learning ability. a) The latency to find the hidden platform each day. Memory retention ability was assessed by the hidden platform trials on day 7, the representative swimming routes (b), the swim speed (c), the time spent in the target quadrant (d), and the target platform crossings (e). N = 15 mice in per group, two-way repeated-measures ANOVA with a post hoc Bonferroni’s test in (a), one-way ANOVA with a post hoc Tukey’s test in others, *p < 0.05 versus WT mice; # p < 0.05 versus WT-Aβ mice. Data were expressed as mean±SEM.

The synaptic plasticity is decreased in CB2R–/–-Aβ mice compared with that in WT-Aβ mice. a) LTP measurements in the hippocampal CA1 region of acute slices from WT-Aβ mice and CB2R–/–-Aβ mice. b) Quantitative analyses of the magnitude of LTP. c,d) Dendritic spine density in hippocampal CA1 of WT-Aβ mice and CB2R–/–-Aβ mice was detected by Golgi staining. Representative shaft dendrite s(c) and the quantification of spine density (d) in hippocampal neurons of the CA1 region. Scale bars: 10μm. N = 8–10 slices from 5 mice for each group for LTP, N = 3 mice for each group for Golgi staining, student unpaired t test, **p < 0.01 versus WT-Aβ mice. Data were presented as mean±SEM.
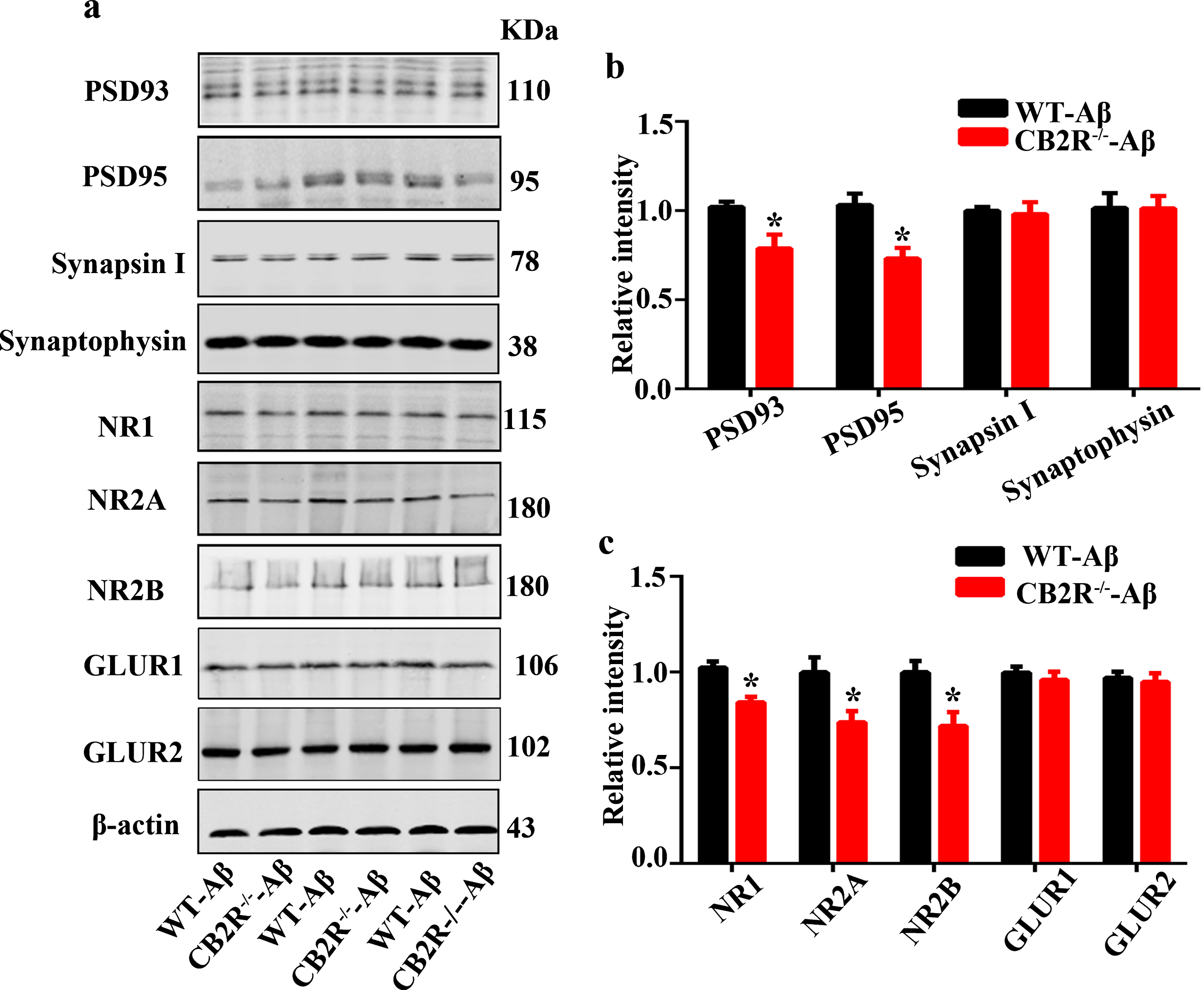
The expressions of synaptic-related proteins are decreased in CB2R–/–-Aβ mice compared with those in WT-Aβ mice. After performing the behavior test, the mice were sacrificed, and extracts of the hippocampus were prepared for protein level measurement with western blot. a) Representative images show the expression levels of PSD93, PSD95, Synapsin I, Synaptophysin, NR1, NR2A, NR2B, GLUR1, and GLUR2 in the hippocampus. b,c) Quantitation of the immunostaining densities shown in (a). N = 6 mice in per group, student unpaired t test, *p < 0.05 versus WT-Aβ mice. Data were presented as mean±SEM.
JWH133 reduces the levels of Aβ1 - 42 and Aβ1 - 40, and increases the levels of ACE and IDE in N2a cells with AβPP overexpression
To confirm that the accumulation of Aβ is mediated by the deletion of CB2R in CB2R–/–-Aβ mice. N2a cells were transfected with plasmids that encoded Myc-tagged human APP (APP-Myc) for 24 h and were subsequently administered JWH133 or combined with AM630 for 24 h. JWH133 increased the levels of CB2R (Fig. 7a, b), ACE (Fig. 7e), and IDE (Fig. 7f) and decreased the levels of Aβ1 - 40 (Fig. 7c) and Aβ1 - 42 (Fig. 7d). In contrast, AM630 mitigated the activation of CB2R induced-protective effects, such as the decrease of Aβ and enhancement of IDE and ACE (Fig. 7c-f). These results may explain why CB2R–/– mice more easily form Aβ accumulation and are susceptible to exogenous Aβ-induced cytotoxicity.

The activation of CB2R with JWH133 reduces the levels of Aβ1 - 40 and Aβ1 - 42 and increases ACE and IDE; these effects disappeared when united with AM630. N2a cells were transfected with APP-Myc for 24 h and were subsequently administered JWH133 or combined with AM630 for 24 h. The cell lysates were prepared for western blot or ELISA. a,b) The expression levels of CB2R, Myc and APP in N2a cells and quantitative analysis. c,d) The levels of Aβ1 - 42 and Aβ1 - 40 were measured by ELISA, human Aβ40 (E-EL-H0542c) and human Aβ42 (E-EL- H0543c) ELISA kits were used. e,f) The levels of ACE and IDE were measured by ELISA. The experiments were performed at least three independent assays (N = 3), one-way ANOVA with a post hoc Tukey’s test, $$p < 0.01 versus CON; *p < 0.05; **p < 0.01 versus APP+DMSO, # p < 0.05; # # p < 0.01 versus APP+JWH. Data were expressed as mean±SEM.
DISCUSSION
In the present study, we showed that CB2R deletion causes differential expression of special proteins associated with Aβ degradation via proteomic analysis. We subsequently determined that Aβ1 - 42 accumulation is clearly increased in CB2R–/–-Aβ mice compared with that in WT-Aβ mice. For the first time, we demonstrated that the levels of ACE and IDE decrease and contribute to Aβ accumulation in the CB2R–/–-Aβ mouse model. The current study supported the previous opinion that CB2R is involved in Aβ degradation rather than in Aβ production and Aβ aggregation [28, 30]. CB2R activation with its specific agonist, JWH-015, stimulates Aβ removal by human macrophages [44]; chronically administration of the CB2R agonists JWH-133 and WIN55,212-2 showed marked beneficial effects and increased Aβ clearance in Tg APP 2576 mice [30]. APP/PS1/CB2R–/– mice (CB2R–/– bred with APP/PS1 mice) exhibit increased Aβ deposits in the cerebral cortex and hippocampus of the brain at 6 months of age, accompanied by a reduction of microglia [28]. Taken together, our current and previous studies demonstrated that CB2R plays a crucial role in Aβ degradation.
ACE is a zinc-dependent peptidase responsible for converting angiotensin I into the vasoconstrictor angiotensin II. It is also a potential susceptibility gene of AD [45]. ACE overexpression in human neuroblastoma cells promotes the degradation of Aβ [12, 46]. In the current study, we demonstrated that the ACE content is significantly lower in CB2R–/–-Aβ mice than in WT- Aβ mice based on the data of the proteomic analysis and ELISA assay. Increasing evidence has shown that proteomic analysis is a better approach to riddle valuable protein; however, the complexity of proteomics and technical limitation of proteomic tests make it necessary to prudently interpret data from proteomic analyses. In general, positive results from a proteomic analysis may provide a valuable clue for further research in contrast to negative results, moreover, the results must be confirmed by others assay [47]. Apart from ACE, Aβ may also be degraded by another degradative enzyme referred to us IDE; IDE is a metalloendopeptidase that hydrolyzes several regulatory peptides such as insulin, Aβ and AβPP intracellular domain [48]. Increasing evidence has shown that IDE plays a key role in Aβ degradation [14, 50]. We attempted to determine the IDE content with ELISA despite the lack of change in IDE by proteomic analysis. Interestingly, our results showed that the levels of IDE also significantly decrease and the levels of Aβ1 - 42 significantly increase in CB2R–/–-Aβ mice compared with those in WT-Aβ mice. Moreover, the levels of Aβ1 - 40 exhibited an increasing trend; however, there was no statistical significance. These data support the previous opinion regarding ACE and IDE degrading Aβ and suggest that the reduction of ACE and IDE content results in a dysfunction of Aβ degradation and more accumulation of exogenous Aβ in CB2R–/–-Aβ mice.
In the current study, our data showed that there is more accelerated behavior impairment in the CB2R–/–-Aβ mice than in the WT-Aβ mice, which are consistent with previous studies; for example, CB2R activation restored cognition and memory impairments induced by bilateral microinjection of Aβ1 - 40 into the hippocampal CA1 area of rats [27]. The specific CB2R agonist JWH-133 induced cognitive improvement in a transgenic mouse model of AD (APP/PS1 mice) [51]. However, several studies have indicated that a lack of CB2R does not accelerate memory impairment in APP/PS1 mice [28, 52]. This discrepancy might be because of differences in the ages of the mice and the experimental conditions.
Hippocampal LTP is a benign form of synaptic plasticity and has been widely used as an electrophysiological model of learning and memory [53]. Aβ-induced LTP impairments in vitro and in vivo have been reported [53–55]. In the present study, we recorded hippocampus LTP, and demonstrated that CB2R deficiency exacerbated the impairments of hippocampus LTP induced by Aβ1 - 42. Our results were in consistent with previous studies which demonstrated that synaptic plasticity impairment induced by the deletion of CB2R [36] and systemic administration of CB2R agonist significantly ameliorated the impaired synaptic plasticity in the hippocampus induced by Aβ [27]. Furthermore, Aβ-induced LTP impairments always accompany synaptic protein changes in presynaptic and postsynaptic proteins [56, 57]. We attempted to clarify whether presynaptic and/or postsynaptic proteins modulates the synaptic transmission. The result indicated that the level of postsynaptic proteins decreased, whereas the presynaptic proteins had no effects, which suggests that the more impairments of synaptic plasticity induced by Aβ in CB2R–/– mice than in WT mice are attributed to the reduction of postsynaptic proteins.
In N2a cells with transfected APP-Myc, JWH133 increased the levels of IDE and ACE and decreased the levels of Aβ1 - 40 and Aβ1 - 42. The effects of JWH133 disappeared under the condition of CB2R inactivation. Therefore, our study indicated that a lack of CB2R results in Aβ accumulation because of deficits in IDE and ACE expression in CB2R–/– mice.
Taken together, our study demonstrated that CB2R–/– mice display more susceptibility to the toxicity of Aβ as a result of dysfunction of Aβ degradation and suggest that CB2R may represent a new target for AD therapy.
Footnotes
ACKNOWLEDGMENTS
This work was supported by the National Nature Scientific Fund of China (81671262) and Self-innovation fund of HUST and Integrated Innovative Team for Major Human Diseases Program of Tongji Medical College, HUST. The authors would like to thank Dr. Nancy E. Buckley for providing CB2R–/– mice.