
Abstract
Dysregulation of neurogenesis in the subgranular zone (SGZ) of the hippocampal dentate gyrus has been related to cognitive deficits and memory loss in neurodegenerative diseases, such as Alzheimer’s disease (AD). Members of the B group of SOX transcription factors play critical roles in regulating neurogenesis in the embryonic and adult nervous system, including maintaining the multipotency, renewal, and cell fate decision of neural stem/progenitor cells. The aim of the present study was to evaluate the expression patterns of selected SOXB proteins in the SGZ, of 8-week-old male and female 5xFAD mice, which represent a transgenic model of AD with a severe and very early development of amyloid pathology. Immunohistochemical analysis showed a significant decrease in the number of cells expressing SOX1, SOX2, and SOX21 transcription factors within the SGZ of 5xFAD mice in comparison to their non-transgenic counterparts which coincidences with reduced number of doublecortin immunoreactive immature neurons found in Tg males. Despite observed changes in expressional pattern of examined SOXB proteins, the proliferative capacity evaluated by the number of Ki-67 immunoreactive cells remained unaffected in transgenic mice of both genders. Based on our results, we suggest that SOXB proteins might be considered as new biomarkers for the detection of early impairments in adult neurogenesis in different animal models or/and new targets in human regenerative medicine.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is an age-associated progressive neurodegenerative disorder with a sex-related epidemiological profile, affecting two times more women than men. It is characterized by multiple neuropathological features, such as intraneuronal accumulation of neurofibrillary tangles, extracellular deposition of amyloid-β plaques, and synaptic and neuronal loss that gradually leads to cognitive deficits and memory loss [1].
The hippocampus, part of the limbic system important for learning and memory, is one of the earliest brain areas affected in AD [2, 3]. There is emerging evidence that alternations in hippocampal neurogenesis occur at the very early stage of AD pathology [4, 5]. It is widely accepted that neurogenesis significantly contributes to generation of new neurons during adult life [6]. Therefore, the disruption of this process is considered to be a main obstacle in endogenous neuronal repair mechanisms as AD pathology proceeds and could significantly contribute to related cognitive dysfunctions [2, 7].
So far, different animal models of AD have been used to study the impact of AD pathology on neurogenesis [2, 9]. These studies provided large number of conflicting evidences of neurogenesis dysregulation prior and following the onset of AD pathology [4, 10– 16]. As it has been extensively discussed, the discrepancies may come from different experimental conditions, including animal age, model type, age of disease onset, and markers used to characterize neurogenesis [2, 9].
In the adult hippocampus, neurogenesis occurs in the subgranular zone (SGZ) of the dentate gyrus (DG). It is currently accepted that it is initiated from “type 1 cells”, a population of quiescent and slow dividing neural stem cells that have radial glia properties. These cells, upon activation, give rise to intermediate progenitor pool, “type 2 cells” that mark a transition from cells with a highly proliferative ratio and multipotent stem cell phenotype to cells with features of the neuronal lineage, such as the expression of doublecortin (DCX) and NeuroD1 proteins [17]. Following a migratory neuroblast-like stage (“type 3 cells”), the lineage-committed cells exit the cell cycle and enter a maturation stage, during which they extend their dendrites into the molecular layer and their axon to the CA3 region of the hippocampus [17, 18].
Molecular mechanisms involved in regulation of cellular processes of distinct cell populations in DG under physiological and pathological conditions are largely unknown [2, 7]. Members of the B group of Sry-related Sox family of genes that encode transcription factors govern diverse cellular processes in the brain during development and adulthood [19–21]. Based on sequence homology and functional analysis, Sox genes are divided into ten distinct groups designated from A to J [22]. SoxB genes, in particular, are subdivided into subgroup B1 (Sox1, Sox2, and Sox3) and B2 (Sox14 and Sox21), acting as either activators or repressors, depending mostly on the cellular and genomic context [23, 24]. In the adult brain, SOX1-3 are predominantly expressed in cells within the neurogenic zones, maintaining progenitor multipotency and renewal [21, 25]. The proper SOX14 protein expression was described to be of importance during the development of neurons involved in circadian rhythm [26]. Its function in neurogenesis, however, remains largely unknown. Multiple roles of SOX21 transcription factor in the progression of neurogenesis have also been recently documented [27–29].
The aim of the present study was to evaluate the expression pattern of SOXB proteins in SGZ, the adult hippocampal neurogenic niche, in the 5xFAD transgenic mouse model of AD that rapidly develops severe amyloid pathology [30]. Expression pattern was analyzed in 8-week-old animals, at the time point when formation of amyloid-β (Aβ) plaques and first signs of impaired neuronal differentiation occur [30, 31]. Considering that development of AD pathology has been reported to be more progressive in females in this mouse model [30], animals of both genders were included in our study.
MATERIALS AND METHODS
Animals
5xFAD transgenic (Tg) mice and their appropriate controls (non-transgenic mice, non-Tg) were used in this study. The 5xFAD Tg model, developed by Oakley and colleagues [30], co-overexpresses five familial AD mutations: Swedish (K670N, M671L), Florida (I716V), and London (V717I) mutations in human amyloid-β protein precursor (AβPP695) and two mutations (M146L and L286V) in the human presenilin 1 (PS1) protein. The 5xFAD mice were purchased from the Jackson Laboratory (Bar Harbor, ME, USA) and maintained on a B6/SJL hybrid genetic background by crossing 5xFAD heterozygous males with B6/SJL F1 females (Jackson labs, Bar Harbor, ME, USA).
A total of 24 animals were included in the study: Tg male mice (n = 6), non-Tg male mice (n = 6), Tg female mice (n = 6) and non-Tg female mice (n = 6). All animal procedures were in compliance with the Directive 2010/63/EU on the protection of animals used for experimental and other scientific purposes, and were approved by the Ethical Committee for the Use of Laboratory Animals of the Institute for Biological Research “Siniša Stanković”, University of Belgrade. The animals were housed under standard conditions (23±2°C, relative humidity 60– 70%, 12 h/12 h light/dark cycle, free access to food and water) and their health status was routinely controlled.
Tissue collection, processing, and section preparation
At 8 weeks of age, animals were anesthetized with intraperitoneal injection of ketamine (100 mg/kg), followed by transcardial perfusion with 0.1 M phosphate-buffer saline (PBS, pH = 7.4). The brains were quickly removed from the skull and divided into left and right hemispheres on ice. The left hemispheres were used for immunohistochemical analysis. Therefore, left hemispheres were fixed in fresh 4% buffered PFA (PBS) for 24 h and further processed for paraffin embedding by standard procedure. Brain sections, 4μm thick, were cut on a microtome, transferred to a water bath, mounted on glass slides (Superfrost™ Ultra Plus Adhesion Slides, Thermo Scientific, USA) and dried for 2 h at 56°C. The level of sections was approximately – 1.656 to – 2.255 mm from the bregma according to the Allen Mouse Brain Atlas [32], while two hippocampal sections per animal (distance between each section was at least 200μm) were used for the detection and quantification of Ki-67, DCX, SOX1, SOX2, and SOX21 immunoreactive cells. Thus, for each of the mentioned antibodies two distant sections of the hippocampus per animal were immunohistochemically stained.
Immunohistochemistry
Brain sections were dewaxed with clearing agent (Xylene, Zorka Pharma-Hemija, Serbia), rehydrated in a series of decreasing ethanol solutions (Ethanol 100%, 96%, 70%, Zorka Pharma-Hemija, Serbia) and treated with citrate buffer (pH = 6.0) in a microwave (21 min, 90°C) for antigen retrieval. The endogenous peroxidase activity was neutralized with 3% hydrogen peroxide (H2O2), followed by 1-h incubation in normal rabbit or goat serum (VECTASTAIN Elite ABC HRP Kit - Peroxidase, Goat IgG, PK-6105; Peroxidase, Rabbit IgG, PK-6101, Vector Laboratories) depending on the antibody used in the reaction. Sections were incubated overnight in the following antibodies: anti-Ki-67 antibody (rabbit polyclonal IgG, Abcam, ab15580, dilution 1:3500); anti-DCX antibody (Doublecortin C-18, goat polyclonal IgG, Santa Cruz Biotechnology, sc-8066, dilution 1:100); anti-SOX1 antibody (rabbit polyclonal IgG, Cell signaling, 4194, dilution 1:100); anti-SOX2 antibody (rabbit polyclonal IgG, Active Motif, 39823, dilution 1:4000); anti-SOX21 antibody (goat polyclonal IgG, R&D systems, AF3538, dilution 1:1000). After overnight incubation, sections were further incubated for 1 h in biotinylated rabbit anti-goat IgG or biotinylated goat anti-rabbit IgG, followed by ABC-complex (VECTASTAIN Elite ABC HRP Kit - Peroxidase, Goat IgG, PK-6105; Peroxidase, Rabbit IgG, PK-6101, Vector Laboratories). Between each step, sections were washed in fresh PBS. Visualization of the immunoreactive sites was done by nickel enhanced 3,3’-diaminobenzidine chromogen (DAB Peroxidase (HRP) Substrate Kit (with Nickel), SK-4100, Vector Laboratories). Sections were counterstained with Mayer’s hematoxylin solution, dehydrated in a series of increasing ethanol solutions, cleared in xylene, mounted with DPX (44581, Sigma, St. Louis, MO, USA) and coverslipped.
Quantification of immunoreactive cells
Quantification of Ki-67, DCX, SOX1, SOX2, and SOX21 immunoreactive cells in stained hippocampal slices was performed using Leica DM4000 B LED microscope with digital camera Leica DFC295 and Leica Application Suite (LAS, v4.4.0) software system. The counting of immunoreactive cells was done in the SGZ of the hippocampal DG, which was defined as a border region between the hilus and the granule cell layer as described by Moon and colleagues [31]. The length of the SGZ was determined for each hippocampal section using the Leica Application Suite (LAS, v4.4.0) software system, and the number of immunoreactive cells was counted along the measured SGZ length. The counting of labeled cells in the SGZ was done under x40 objective, regardless of their size and shape. The number of counted immunoreactive neurons was expressed per 1 mm of length of the SGZ in order to standardize the number of counted cells [31]. The representative photomicrographs from each group of animals and for each antigen were made with OlympusBX41 light microscope and OlympusC5060A-ADU digital camera under x40 objective and x100 oil immersion objective.
Fluorescent immunohistochemistry staining

Immunohistochemical analysis of Ki-67 and DCX protein expression in the SGZ of non-Tg and Tg 5xFAD mice. A-D) Representative images of Ki-67 immunoreactivity in non-Tg (A) and Tg male mice (B) and non-Tg (C) and Tg (D) 5xFAD female mice. E) Statistical analysis of the Ki-67+ cells in different animal groups. F-I) Representative images of DCX immunoreactivity in non-Tg (F) and Tg male mice (G) and non-Tg (H) and Tg (I) 5xFAD female mice. J) Statistical analysis of the DCX+ cells in different animal groups. The number of cells is expressed per 1 mm of the SGZ length. Results are presented as mean±standard deviation. Objective magnification: 40x; Boxed images – 100x oil immersion. DCX, doublecortin. *p < 0.05, ***p≤0.001.
For immunofluorescent detection of SOX1, SOX2, PCNA, and DCX, sections were dewaxed with clearing agent, rehydrated in a series of decreasing ethanol solutions and treated with citrate buffer for 15 min at 121°C in the pressure cooker. Sections were subsequently blocked using 1% BSA, 10% normal horse serum (Sigma-Aldrich Co.) in PBS for 1 h at room temperature and next incubated with primary antibodies: rabbit anti-SOX1 (1:50), rabbit anti-SOX2 (1:1000), mouse anti-PCNA (1:100, mouse monoclonal IgG, sc-56, Santa Cruz Biotechnology) and goat anti-DCX (1:300) overnight at room temperature. After washing with PBS, sections were incubated for two hours in goat anti-rabbit secondary antibodies conjugated with Alexa Fluor® 488 (Invitrogen TM, diluted 1:500 in 1% BSA-PBS). The anti-DCX and anti-PCNA antibodies were first labeled with biotinylated donkey anti-goat IgG (Vector, Burlingame, CA, USA) or biotinylated goat anti-mouse IgG (Vector, Burlingame, CA, USA) for 1 h at room temperature, followed by DyLight® 594-streptavidin (Vector, Burlingame, CA, USA, dilution 1:1000) diluted in PBS for 1 h at room temperature. Nuclei were stained with 0.1 mg/ml diaminophenylindole (DAPI; Sigma-Aldrich, MO, USA). Sections were washed with PBS, and mounted with Mowiol (Calbiochem). Samples were analyzed and images were taken using a Leica TCS SP8 confocal microscope and Leica Microsystems LAS AF-TCS SP8 software (Leica Microsystems).
Western blot analysis
The entire right hippocampus was dissected on ice under magnifying glass, snap-frozen in liquid nitrogen, and stored at – 80°C for subsequent protein analyses. For whole-cell protein extracts, hippocampal tissue was homogenized and sonicated in 10 volumes of ice-cold RIPA buffer (50 mM Tris– HCl, pH 7.5, 150 mM NaCl, 1% NP-40, 0.1% SDS, 0.5% Triton x-100, 10 mM EDTA, 10 mM EGTA) containing protease and phosphatase inhibitors (Roche, Pleasanton, CA). Following the centrifugation at 20,000 g at 4°C for 30 min, the supernatants were collected and stored at – 80°C until use.
The protein concentration was determined using the Micro BCA Protein Assay Kit (Pierce Biotechnology, Rockford, IL, USA) and the Bovine Serum Albumin (BSA) as a standard. Equal amounts of proteins (15μg per lane) were separated by SDS-polyacrylamide gel electrophoresis (12%) and blotted onto PVDF membranes (GE Healthcare, Princeton, NJ, USA). To block non-specific binding, the membranes were incubated for 1 h at room temperature in 3% BSA/TBST (150 mM NaCl, 50 mM Tris, pH 7.4, and 0.1% Tween20). Following overnight incubation at 4°C in rabbit anti-BDNF antibody (N-20; Santa Cruz Biotechnology) membranes were incubated for 1 h in anti-rabbit Horse Radish Peroxidase (HRP)-conjugated secondary antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA). HRP-immunoreactive bands were visualized using enhanced chemiluminiscence (ECL, GE Healthcare, Princeton, NJ, USA) and film exposure (MXBE, Carestream Health, Inc., Rochester, NY, USA). Each blot was subsequently re-probed with mouse anti-β-actin antibody (Sigma-Aldrich Biotechnology, St. Louis, MO, USA). Signals were quantified densitometrically using Image Quant software (v. 5.2, GE Healthcare, Princeton, NJ, USA) and expressed as relative values (i.e., normalized to the corresponding β-actin signals). The levels of the target protein in Tg mice were determined as the fold change relative to the appropriate level detected in the control non-Tg mice that were assigned the value 1.
Statistical analysis
Data normality was tested using D’Agostino & Pearson omnibus normality test. To assess the differences between the number of immunoreactive cells in the SGZ of Tg and non-Tg control mice and to assess gender specific differences, two-way ANOVA with Tukey’s multiple comparisons post hoc test was used. For statistical analysis of immunoblot results, Kruskall-Wallis test was used. Results are represented as mean±standard deviation for immunohistochemistry results and as median with interquartile range for immunoblot results, while p values less than ≤0.05 were considered statistically significant.
RESULTS
Decrease in the number of immature granule cells in 5xFAD male mice was not preceded by a decrease in cell proliferation rate in SGZ
Adult hippocampal neurogenesis, formation of new neurons/excitatory granule cells in the DG, comprises series of sequential events such as cell self-renewal, asymmetric division, migration, differentiation and maturation [18]. Accordingly, in the first set of experiments, we have determined the number of proliferating cells and immature neurons in the SGZ. We analyzed the number of dividing cells positive for Ki-67, a protein present during all active phases of the cell cycle [33]. Quantitative analysis of immunohistochemistry results revealed no significant differences in the number of Ki-67-positive cells between non-Tg and Tg mice of both genders (F = 0.3666, p > 0.05) suggesting that cell proliferation was not affected in 5xFAD mouse model of AD at 8 weeks of age (Fig. 1A-E).
DCX is a microtubule-associated protein transiently expressed during neurogenesis in neuroblasts and newly generated neurons [8, 34]. Our immunohistochemical analysis revealed significant differences in the number of DCX-positive cells between non-Tg and Tg male animals (non-Tg versus Tg: 27.98±4.57 versus 17.33±5.05, p≤0.05). However, the number of immunoreactive cells between 5xFAD females and their healthy controls did not differ significantly (non-Tg versus Tg: 36.88±13.81 versus 34.02±8.07, p > 0.05). Notably, in female animals, regardless of the genotype, DCX-positive neurons displayed more pronounced and longer dendrites (Fig. 1H and I versus Fig. 1F and G) in comparison to male counterparts, suggesting important gender-related differences in morphology of hippocampal neurons in 8-week-old mice.
Interestingly, the results of our quantitative study demonstrated significant differences in the number of DCX immunoreactive cells between Tg female and male mice (Tg male versus Tg female: 17.33±5.05 versus 34.02±8.07, p≤0.001) (Fig. 1J). This result further implicates gender-specific differences in potential for neurogenesis in 5xFAD mice.
The number of SOX1-, SOX2-, and SOX21-positive neural stem/progenitor cells was significantly decreased in 5xFAD animal model of AD
Regarding the important roles of SOXB family members in the regulation of neurogenesis, we further analyzed their expression in the hippocampus of 5xFAD model of AD. Our results showed that, in the hippocampus of 8-week-old mice, four examined SOXB members (SOX1, SOX2, SOX3, and SOX21) were expressed within the SGZ with distinct immunohistochemistry patterns. SOX3 protein expression was weakly detected in a small number of cells (data not shown) and thereby was not taken into further analysis. Considering that there is no literature data on SOX14 protein expression in the cells inside neurogenic niches in the adult mice, this transcription factor was excluded from the analysis during the course of the study.
Our quantitative immunohistochemical analysis first revealed a significant decline in number of cells expressing SOX1 protein in 5xFAD mice compared to their control littermates (Fig. 2A-E). The number of SOX1 positive cells was decreased in both male (non-Tg versus Tg: 23.83±6.00 versus 17.47±5.20, p≤0.05) and female Tg animals (non-Tg versus Tg: 30.88±2.87 versus 24.83±5.11, p≤0.05) (Fig. 2E). Surprisingly, female mice of both genotypes showed a higher number of SOX1 positive cells than male mice (non-Tg male versus non-Tg female: 23.83±6.00 versus 30.88±2.87, p≤0.01; Tg male versus Tg female: 17.47±5.20 versus 24.83±5.11, p≤0.01). To the best of our knowledge, this is the first report indicating that the number of cells with SOX1 protein expression in the mouse hippocampus is gender-dependent.
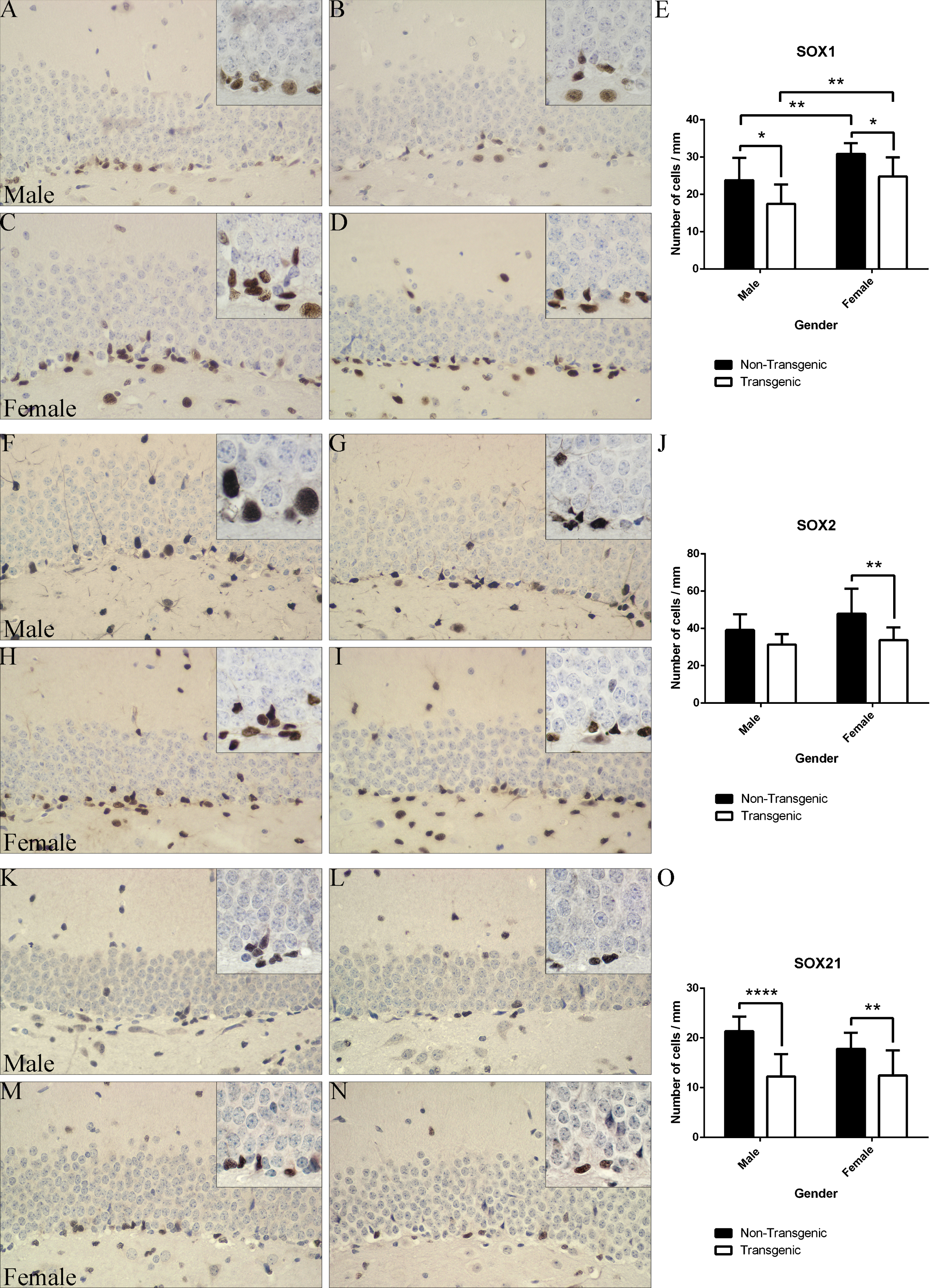
Immunohistochemical analysis of SOX1, SOX2, and SOX21 protein expression in non-Tg and Tg 5xFAD mice. A-D) Representative images of SOX1 immunoreactivity in non-Tg (A) and Tg male mice (B) and non-Tg (C) and Tg (D) 5xFAD female mice; E: Statistical analysis of the SOX1+ cells in different animal groups. F-I) Representative images of SOX2 immunoreactivity in non-Tg (F) and Tg male mice (G) and non-Tg (H) and Tg (I) 5xFAD female mice. J) Statistical analysis of the SOX2+ cells in different animal groups. K-N) Representative images of SOX21 immunoreactivity in non-Tg (K) and Tg male mice (L) and non-Tg (M) and Tg (N) 5xFAD female mice. O) Statistical analysis of the SOX21+ cells in different animal groups. The number of cells is expressed per 1 mm of the SGZ length. Results are presented as mean±standard deviation. Objective magnification: 40x; Boxed images – 100x oil immersion. SOX1, Sry-related HMG (high mobility group) box 1; SOX2, Sry-related HMG (high mobility group) box 2; SOX21, Sry-related HMG (high mobility group) box 21. *p < 0.05, **p≤0.01, ***p≤0.0001.
To further evaluate the expression of SOX1 protein in proliferative cells in SGZ of 8-week-old mice, we performed double immunofluorescent-staining on control (non-Tg) hippocampal tissue using SOX1- and cell cycle marker PCNA (proliferative cell nuclear marker)-specific antibodies. As presented in Fig. 3A1-A6, small population of cells in the SGZ was proliferating (PCNA-immunopositive), and all of the proliferating cells were expressing SOX1 protein. Our next aim was to examine the expression of SOX1 protein in neuroblasts and immature neurons, previously identified with DCX expression (Fig. 1F-J). Our immunofluorescence results revealed that, with exception of small number of cells (yellow arrowheads in Fig. 4), majority of SOX1 immunoreactive cells were not positive for DCX (Fig. 4).
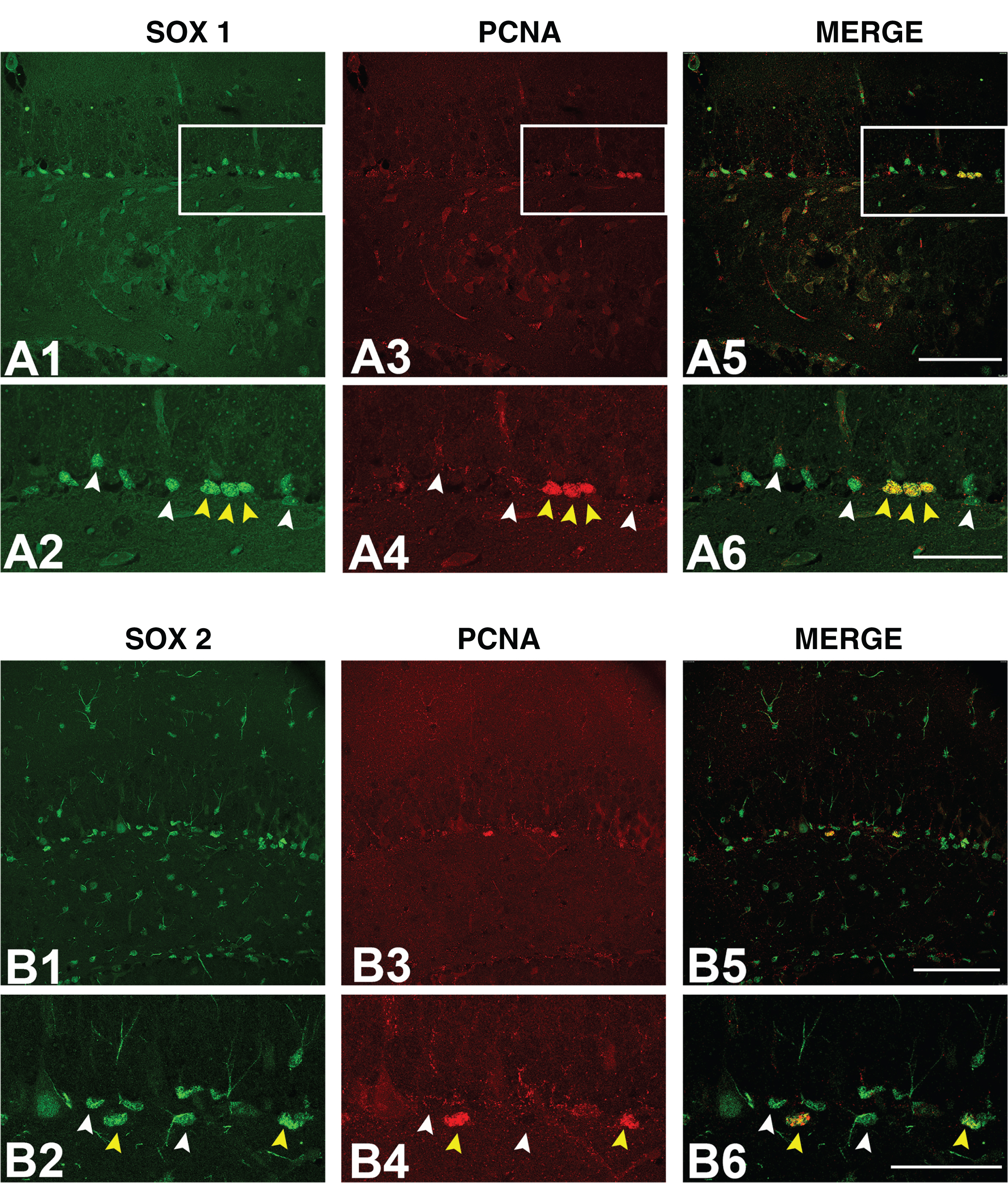
Immunofluorescence detection of SOX1 and SOX2 protein expression in proliferating cells. Representative confocal microscopy images of non-Tg hippocampus stained with anti-SOX1 (A1, A2), anti-SOX2 (B1, B2) and anti-PCNA (A3, A4, B3, B4) antibodies. Boxed regions in A1, A3, A5, B1, B3, and B5 are enlarged in A2, A4, A6, B2, B4, and B6 figures, respectively. White arrowheads in A2, A4, and A6 marks cells that are immunopositive for SOX1, white arrowheads in B2, B4, and B6 marks cells that are immunopositive for SOX2, while yellow arrowheads in A2, A4, A6, B2, B4, and B6 marks cells immunopositive for both proteins (SOX1 or SOX2 and PCNA). Scale bar: 100μm. SOX1, Sry-related HMG (high mobility group) box 1; SOX2, Sry-related HMG (high mobility group) box 2; PCNA, proliferative cell nuclear marker.
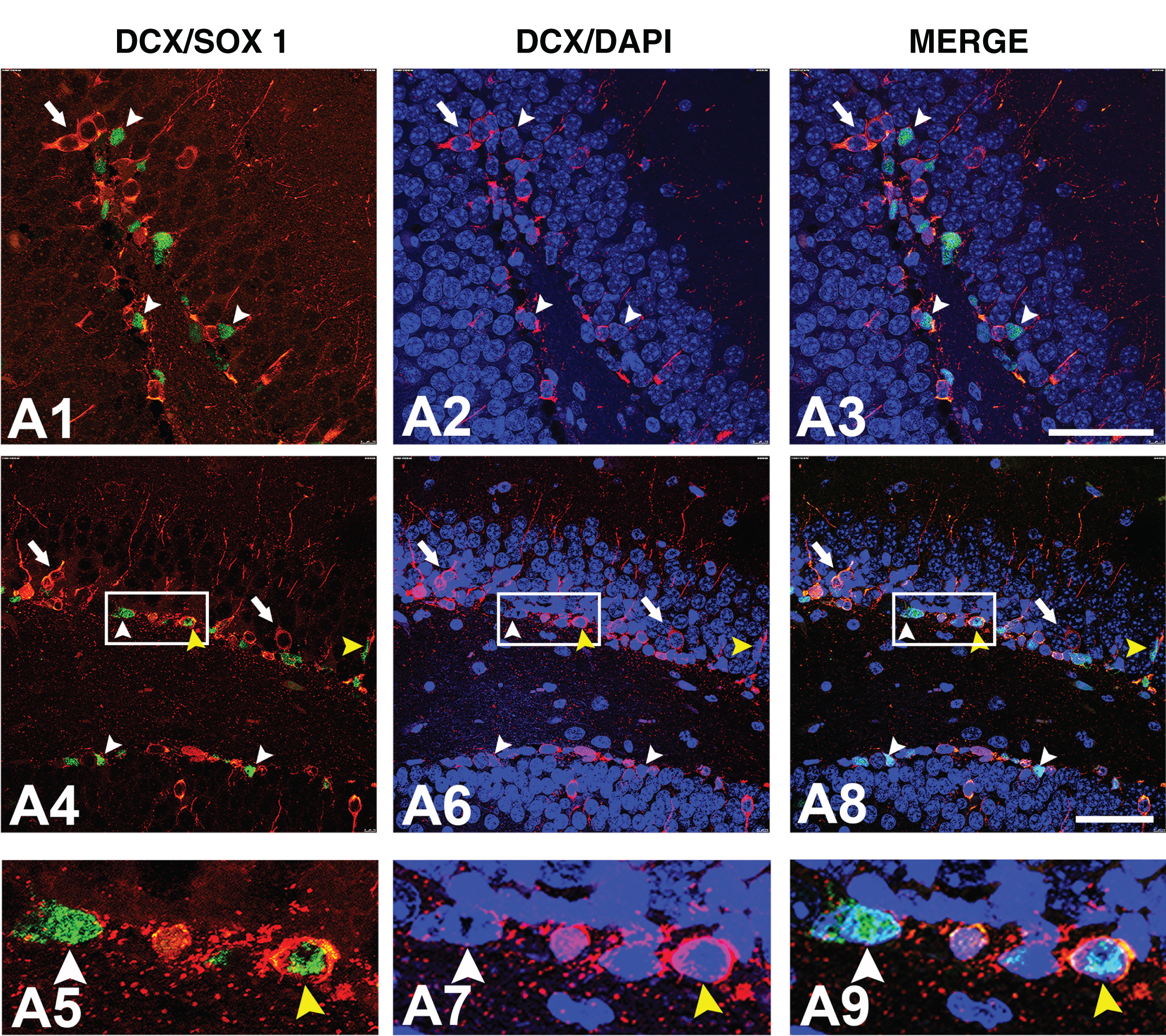
Immunofluorescence detection of SOX1 protein expression in immature granular cells. Representative confocal microscopy images of non-Tg hippocampus stained with anti-SOX1 and anti-DCX (A1-A9) antibodies. Boxed regions in A4, A6, and A8 are enlarged in A5, A7, and A9 figures, respectively. White arrowheads marks cells only immunopositive for SOX1, white arrows marks cells only immunopositive for DCX, while yellow arrowheads marks cells immunopositive for both proteins. Cell nuclei were counterstained with DAPI. Scale bars: 50μm. SOX1, Sry-related HMG (high mobility group) box 1; DCX, doublecortin.
We next evaluated the expression of SOX2 protein in the SGZ of 5xFAD and control mice (Fig. 2F-J and 3B1-B6). Statistical analysis of our results revealed considerable difference in the number of immunoreactive cells between 5xFAD and control females (non-Tg versus Tg: 47.83±13.48 versus 33.73±6.81, p≤0.01). In male animals, however, expression of SOX2 protein remained unaffected (non-Tg versus. Tg: 39.19±8.41 versus 31.3±5.6, p > 0.05; Fig. 2F-J). Furthermore, we did not detect any significant differences in the number of cells expressing SOX2 between different genders of the same genotype (Fig. 2J).
To assess the expression of SOX2 protein in proliferative cells in hippocampus of 8-week-old control (non-Tg) mice, we performed double immunofluorescent staining using SOX2 and PCNA antibodies. As presented on Fig. 3B1-B6, our results revealed different levels of SOX2 protein expression, from moderate to strong, in cells of SGZ. However, only cell nuclei displaying strong SOX2 immunoreactivity were positive to PCNA antibody (Fig. 3B6).
Finally, we examined the expression of SOX21 protein. Extensive quantitative analysis revealed significant decline in cells expressing SOX21 protein in the SGZ of the hippocampus of 5xFAD mice of both genders compared to their non-Tg controls (male animals, non-Tg versus Tg: 21.36±2.95 versus.12.25±4.48, p≤0.0001 and female animals, non-Tg versus Tg: 17.79±3.27 versus12.47±5.03, p≤0.01; Fig. 2K-O). At the same time, there was no statistically significant difference in the number of SOX21 positive cells between animals of the different genders (p > 0.05).
The level of BDNF protein expression in the hippocampus was not altered in 5xFAD mice
To assess whether BDNF, a neurotrophic factor involved in brain neuroplasticity, is changed in the 5xFAD model of AD, western blot analysis of hippocampal homogenates was performed. Statistical analysis showed no significant statistical difference between four groups of animals (p > 0.05), indicating that protein levels of BDNF were not affected in the present model of AD at 8 weeks of age regardless of the genotype and/or gender of mice used (Fig. 5).

BDNF protein levels in the hippocampus of non-Tg and Tg male and female mice. A, B) Representative immunoblots of hippocampal samples from male and female animals included in the study, respectively. C) Statistical analysis of western blot results showed that BDNF protein levels in hippocampal homogenates do not differ between 8-week-old male non-Tg (n = 6) and Tg mice (n = 6) and female non-Tg (n = 6) and Tg mice (n = 5). Results are presented as median with interquartile range. BDNF, brain-derived neurotrophic factor; Tg M, transgenic male; Tg F, transgenic female.
DISCUSSION
The major finding of the present study is a significant decrease in the number of cells expressing SOXB transcription factors with in the SGZ of 8-week-old 5xFAD transgenic mouse model of AD in comparison to their non-Tg counterparts. In Tg male animals, detected decline in the abundance of SOX1- and SOX21-positive cells coincided with reduced number of DCX-immunoreactive neuroblasts/immature neurons. In contrast, in female animals we detected a significant decrease in SOX1-, SOX2-, and SOX21-positive cells while the number of DCX immunoreactive neuroblasts/immature neurons remained the same between experimental groups. Finally, despite observed changes in expressional pattern of examined SOX proteins, the proliferative capacity evaluated by the number of Ki-67 immunoreactive cells remained unaffected in transgenic mice of both genders at this age.
5xFAD mice are known to be an early onset transgenic model as they recapitulate asymptomatic, prodromal-like, and symptomatic phases of AD pathology within the first 6 months of their life [35]. It has been demonstrated that Aβ depositions first emerge in the subiculum area of the hippocampus and in the cortical layer 5 between 8 and 16 weeks, while behavioral impairments starts at the age of 16 weeks, and cortical neuron loss at 9 months of age [30]. More prominent amyloid pathology and behavioral deficits were reported in females [30, 36]. As a robust model, it is considered convenient to assess Aβ impact on neuronal loss and neuroinflammation.
In line with the results obtained from different AD models [4, 5], early disruption of hippocampal neurogenesis, before the appearance of any visible Aβ aggregates in the DG, has been reported in 5xFAD mice [13, 31]. Moon and co-workers revealed that, in these Tg male mice, the number of DCX immunoreactive cells more progressively declines during aging, compared to their counterparts, starting from 8 weeks after birth [31]. Recent study on 5xFAD transgenic animals on different C57BL/6J genetic background reported somewhat different results regarding the hippocampal neurogenesis during aging [13]. A significant increase in the number of DCX immunopositive immature neurons in the DG in Tg male animals, compared to their non-Tg controls, was detected for the first time in 4-month-old mice. However, in older animals (8-month-old male mice), the number of DCX positive cells in the DG was substantially reduced, compared to the controls [13].
The present work demonstrated a significant decrease in the number of DCX immunoreactive cells in 8-week-old 5xFAD male mice in comparison to their non-Tg littermates (Fig. 1J), which is in line with results obtained by Moon and co-workers [31]. In addition to previous studies [13, 31], we analyzed the neurogenesis in the hippocampus of female mice since the development of AD pathology differs between genders [30]. Surprisingly, our quantitative results revealed that in 5xFAD female mice, the number of immature neurons in the DG was not affected at the examined age. Furthermore, immunohistochemical analysis revealed gender-specific differences in morphology of DCX immunoreactive cells (Fig. 1F-I). The pattern of DCX staining in female animals was much more pronounced with labeling of longer dendrites that could correspond to DCX class II immature postmitotic neurons [37, 38]. This further implies gender-specificities on surviving period of class I cells and their differentiation from class I to class II that can be intriguingly important considering the fact that female 5xFAD mice are more susceptible to develop AD-like phenotype [30]. The ability of Aβ peptide to induce an increase in the differentiation rate of newborn neurons in a dose-dependent manner was demonstrated [39]. On the other hand, dendrite development of adult-born granule cells is activity-dependent and regulated by autocrine action of BDNF [40]. Although we did not observe any changes in total BDNF levels, its role in regulation of more prominent dendrite arborization in females cannot be excluded.
In the SGZ of adult mice, transcription factor SOX2 is expressed in radial glial cells (type 1 cells), intermediate progenitor type 2a cells, and, to some extent, in intermediate progenitor type 2b cells [17]. Immunohistochemistry analysis in 8-week-old 5xFAD mice revealed SOX2 protein expression in higher number of cells throughout SGZ in comparison to other examined SOXB proteins (Fig. 2). Intensity of its immunoreactivity differed between cells, with highest signal observed in mitotically active (PCNA positive) cells (Fig. 3B6). These observations are in line with previous published data that different levels of SOX2 expression characterize distinct cell populations [17, 41]. Comparative analysis of immunopositive cells in SGZ of 5xFAD mice and their control littermates revealed significant decline in the number of SOX2 expressing cells in 8-week-old Tg female, but not male animals. Significance of improper SOX2 protein expression in the neural stem/progenitor cell fate specification and differentiation in adult brain has been previously documented. Ferri and colleagues [42] demonstrated that its deficiency impairs neurogenesis and induces neurodegeneration in adult mice brain. A recent study also revealed, by using a transgenic approach with conditional SOX2 expression, that deficiency in its expression in adult hippocampus causes reduction of neurogenesis by increasing apoptosis of neuroblasts and disruption of the maturation of newborn granular cells [43]. Furthermore, the level of SOX2 protein expression was decreased in the brain of a transgenic mouse model of AD [44] as well as in the brain of AD patients where this reduction correlated with the severity of the disease or the patient’s cognitive capacity [44, 45]. Finally, several reports demonstrated functional cross-talk between SOX2 and proteins involved in AD pathology and suggested involvement of this transcription factor in neuroprotective mechanisms [46, 47]. We may suspect that decline of SOX2 expression, detected only in 5xFAD female mice in the present study, is further evidence of earlier effects of AD pathology in this gender and that the compensatory mechanisms did not rely on mechanisms involving SOX2.
This work also provides the first evidence on reduced SOX1 protein expression in the SGZ of hippocampus in mouse model of AD (Fig. 2E). Results of our immunofluorescence analysis on 8-week-old mice (Fig. 3A1-A6, 4) identified SOX1 protein expression in different cell populations throughout SGZ, including proliferating (PCNA positive) “type 1 and 2 progenitor” cells, neuroblasts (DCX positive), and immature (DCX positive) neurons. However, the majority of SOX1 immunoreactive cells did not co-localize with markers used in this study. Based on previously published data on SOX1 expression in the hippocampus [48, 49], we may speculate that SOX1 expressing cells could partially represent a population of quiescent “type 1” cells and partially unidentified cell populations within the SGZ providing a transitional stage of neural differentiation. In contrast to SOX2, studying the role of SOX1 transcription factor in maintenance of progenitor pool and/or differentiation in neurogenesis is still insufficient and conflicting [50]. Initial studies showed that SOX1 promotes in vitro neuronal differentiation in P19 cells [25] and convert Xenopus ectodermal explants into neuronal tissue [51]. In contrast, studies by Elkouris and colleagues [52] showed that in the mouse developing cortex SOX1 regulates the maintenance of the undifferentiated state of progenitor cells via a mechanism involving suppression of Prox1– mediated cell cycle exit and neurogenesis. Latter results are in the line with data obtained from gain-of-function and loss-of-function experiments according to which all members of SOXB1 group block neurogenesis downstream of proneural factors [53, 54]. Finally, we show for the first time that there is a difference in the number of SOX1 protein expression between genders, in both non-Tg and Tg mice (Fig. 2E). We may speculate that this unique property of SOX1 protein could be a consequence of epigenetic-related mechanisms involved in brain differentiation [55], however further studies are needed to clarify this phenomenon.
The present study also provides first evidence on impaired SOX21 protein expression in the SGZ of hippocampus in mouse model of AD (Fig. 2K-O). Among all of the examined SoxB members, the cells expressing SOX21 protein are the most strikingly diminished. Evidence from several reports on expression and multiple function of SOX21 protein in regulation of embryonic and adult neurogenesis is, however, conflicting. Studies in mice showed that SOX21 protein is necessary for promoting neural differentiation in adult hippocampus, by inducing transition from neural progenitor cells type 2a to type 2b [28]. However, some studies suggested that SOX21, similarly to SOXB1 proteins, regulates maintenance of progenitor identity and that decrease of SOX21 expression may be necessary for differentiation to occur [56–58]. A model that reconciles both effects has been proposed suggesting that SOX21 regulates diverse mechanisms in dose-dependent fashion [29].
In summary, we demonstrated that there is a downregulation of SOX1, SOX2, and SOX21 expression in adult hippocampal neurogenic zone of 5xFAD mouse model of AD that proceeds disturbance of neural/progenitor cell proliferation [13] or, in the case of Tg female animals, decline in the number of DCX expressing neuroblasts and immature neurons. As previously reported, improper neural cell proliferation and differentiation is detected in various pathological conditions [59, 60]. Furthermore, even the minor alternations in SOXB1 expression level are associated with several human pathologies such as severe eye, urogenital, and esophageal defects and seizures [42, 62]. Therefore, further investigations of molecular mechanisms involved in impaired neurogenesis under pathological conditions that rely on SOXB genes, could contribute to better understanding of molecular mechanisms important for future strategies in the field of regenerative medicine.
Footnotes
ACKNOWLEDGMENTS
This work was supported by the Ministry of Education, Science and Technological Development of the Republic of Serbia (grants № ON175061, ON173056 and ON173051), NIH (grant № R03AG046216) and “MedSAP Research Network” project supported by the U.S. Embassy in Belgrade (Award Number: SRB100-15-GR-355).