
Abstract
Alzheimer’s disease (AD) is a neurodegenerative condition affecting millions of people worldwide. It is associated with cerebral amyloid-β (Aβ) plaque deposition in the brain, synaptic disconnection, and subsequent progressive neuronal death. Although considerable progress has been made to elucidate the pathogenesis of AD, the specific causes of the disease remain highly unknown. Recent research has suggested a potential association between certain infectious diseases and dementia, either directly due to bacterial brain invasion and toxin production, or indirectly by modulating the immune response. Therefore, in the present review we focus on the emerging issues of bacterial infection and AD, including the existence of antimicrobial peptides having pore-forming properties that act in a similar way to pores formed by Aβ in a variety of cell membranes. Special focus is placed on oral bacteria and biofilms, and on the potential mechanisms associating bacterial infection and toxin production in AD. The role of bacterial outer membrane vesicles on the transport and delivery of toxins as well as porins to the brain is also discussed. Aβ has shown to possess antimicrobial activity against several bacteria, and therefore could be upregulated as a response to bacteria and bacterial toxins in the brain. Although further research is needed, we believe that the control of biofilm-mediated diseases could be an important potential prevention mechanism for AD development.
INTRODUCTION
Alzheimer’s disease
In recent years, considerable progress has been made regarding the understanding of the pathogenesis, diagnosis, and treatment of Alzheimer’s disease (AD). However, it still remains a fatal condition with significant negative impacts on economic and social issues. Until now, there are no effective therapeutic approaches to prevent, delay, or even reverse the condition, which starts with an initial mild loss of cognitive function and progresses toward language problems, memory loss, motor difficulties, and ultimately death [1]. The shift in demographics toward a highly prevalent elderly population is further increasing the prevalence of AD worldwide and has increased the financial burden of the disease (e.g., medications, nursing care). Currently, it is estimated that in the US alone, more than 5 million people aged 65 or older suffer from AD, and the costs for treating the disease are estimated at over US$180 billion per year [2, 3].
AD is a neurodegenerative disease, which is associated with neuronal loss in several areas of the brain, such as frontal cortex and hippocampus, and can be classified as either “early-onset” (also known as “familiar”) or “late-onset” [4]. The late-onset form of the disease is the most prevalent and is considered to be the main worldwide cause of dementia in the elderly population [5]. Although it is believed that late-onset AD is mostly determined by environmental factors, the presence of the apolipoprotein E (APoE) ɛ4 allele has been demonstrated to increase the risk of developing AD [6].
In the present review, we focus on the emerging issues of brain bacterial infection and AD, and on the existence of antimicrobial peptides having pore-forming properties that function in a similar way to the pore formed by amyloid-β (Aβ) in a variety of cell membranes. Thus, it is possible that brain microbial infection might accelerate/precipitate the onset of AD and that Aβ might act as an antimicrobial or as an enhancer of the neurodegenerative process.
The amyloid cascade as a cause of Alzheimer’s disease
There is abundant evidence, based on genetic and animal data, suggesting that Aβ peptide accumulation first initiates and then might be acting as a causative agent of the disease. For instance, the existence of mutations in genes for the amyloid precursor protein (APP) (chromosome 21) and presenilins 1 (chromosome 14) and 2 (chromosome 1) [7] has been reported in a small number of AD patients. Furthermore, as mentioned above the ApoE E4 allele has been associated to sporadic AD [8]. On the other hand, environmental factors and lifestyle habits such as diet, physical exercise, and education are also believed to affect the onset of AD [9]. One of the main histological markers of AD are senile plaques that are made up mainly of Aβ [10]. Although its function is still unknown [11], Aβ is a 39–42 amino acid peptide generated by the enzymatic processing of amyloid-β protein precursor (AβPP), which has sites for α- β- and γ-secretases, of which β− and γ− secretases produce Aβ [12]. Aβ is mainly found in the extracellular matrix in the brain and cerebrospinal fluid at nanomolar concentrations and is widely accepted as the main neurotoxic agent in the disease [13]. Early manifestations of AD are associated with the synaptotoxic effects produced by soluble oligomeric forms of Aβ [14].
The central dogma is that monomers of Aβ aggregate to form small molecular weight species, also referred to as oligomers [14], which are believed to be highly neurotoxic [15]. Although there is no final conclusion, diverse forms from dimers to 56 kDa oligomers appear to be the most important causal agents in the disease [16, 17]. It is believed that before affecting neuronal function, the peptides have to associate with the cell membrane. It was recently shown that Aβ association to the membrane resulted in the leakage of important cellular components, as well as entry of calcium into the cell [15]. The unregulated entry of calcium can by itself cause alterations in synaptic transmission, cell toxicity, and death [18]. This experimental evidence points to the idea that Aβ toxic events are multiple and that one/several of them might serve as a therapeutic target. Nowadays, there is a reasonable consensus that Aβ will provide the first, early cellular toxic stress and that tau hyperphosphorylation will subsequently orchestrate the final deadly blow to the neuron [10].
Pathogenesis of AD
In terms of pathogenesis, AD shows two distinct structures in affected neurons known as neurofibrillary tangles (NFT) and the previously described senile plaques [19, 20]. NFT are a characteristic structure found in the brain of AD patients and correspond to intracellular accumulations of hyperphosphorylated and misfolded tau protein [21, 22]. Tau protein is normally found in the cytoplasm of neuronal cells and along their axons within the CNS [23] and when phosphorylated has the tendency to form paired helical filaments that accumulate as NFT [24]. There are six isoforms of the tau protein, and all six are hyperphosphorylated in AD (reviewed in [25]). Therefore, potential therapies against tau pathogenesis are mostly directed against hyperphosphorylation, although they also include approaches to inhibit aggregation, as well as tau-directed immunotherapies [26]. Despite the above knowledge, the consequence of NFT accumulation in the central nervous system (CNS) is not yet fully understood.
On the other hand, and as previously mentioned, senile plaques are constituted by Aβ deposits generated by the cleavage of the AβPP. Reasons for the presence of the precursor protein in the CNS and why it is cleaved to Aβ are not well understood. However, it is known that Aβ is able to rupture the neuronal plasma membrane by the formation of pores leading to cytoplasmic leakage and cell death [27, 28], by either direct lipid disruption or by its interaction with ion channels in the membrane [15, 29]. Some studies have shown that reduced cerebrospinal fluid levels of Aβ42 in patients showing mild cognitive impairment is strongly indicative of disease progression [30 –32].
Interestingly, it has been recently shown that Aβ functions as an antimicrobial peptide, which has the capacity to kill various strains of bacteria such as Escherichia coli (E. coli), Staphylococcus aureus (S. aureus), Streptococcus pneumoniae (S. pneumoniae), Enterococcus faecalis (E. faecalis), and the fungus Candida albicans (C. albicans) [33]. The mechanism of action of Aβ as an antimicrobial peptide is believed to stem from its ability to entrap microbial cells and avoid their adhesion to host cells [34]. There has also been evidence that Aβ has a protective activity against certain viruses and may be overexpressed in patients with viral infections [35, 36].
Similarities between AD and infectious dementia
Some neurodegenerative diseases present strong infectious components in their pathogenesis, and therefore are commonly referred to as infectious dementia. One of these pathologies is Creutzfeldt-Jakob disease, which is a fatal brain disorder characterized by behavioral changes, memory loss and progressive dementia, among other symptoms [37, 38]. All these symptoms are very similar to the ones observed in AD patients. One of the causes of Creutzfeldt-Jakob disease is believed to be prion accumulation which can be acquired by exposure to brain tissue from an infected person [39]. The variant form of Creutzfeldt-Jakob disease (i.e., bovine spongiform encephalopathy) is a similar disease, but in this case the transmission is from cows to humans [40]. Other forms of infectious dementia include toxoplasmosis, syphilis, and human immunodeficiency virus (HIV) [41]. Interestingly, the symptomatology and pathology of general paresis in advanced stages of neurosyphilis is remarkably similar to AD [42]. Although AD is not traditionally considered as infectious, there has been increasing evidence that microorganisms may play a central role in the disease [43]. There is also recent evidence that Parkinson’s disease may have an infectious influence, as it has been shown to be associated to influenza virus [44].
THE ROLE OF BIOFILMS IN SYSTEMIC HEALTH AND DISEASE
Currently, it is known that biofilms are associated to a wide range of chronic diseases in the human body [45]. A biofilm refers to a community of different microorganisms (i.e., bacteria, virus, fungi) embedded in an extracellular matrix that confers resistance to antibiotic attack and facilitates nutrient exchange and adhesion to substrates [46, 47]. Biofilms are initiated by the irreversible adhesion of microorganisms onto a surface, and factors promoting this attachment include surface roughness, hydrophobicity and presence of bacterial appendages (i.e., fimbriae) [48 –50]. These biofilms are not only present on biological tissues, but also on surfaces of biomedical and prosthetic devices and are involved in the development of diseases such as tooth cavities, periodontal disease, urinary tract infections, and bacterial vaginosis [51 –54]. However, they are not only responsible for site-specific infection because single cells can detach from the biofilm mass and migrate to other sites and spread infection (reviewed in [55]).
In humans, biofilms can be found in different regions of the body, with the most important areas being the skin, mouth, gut, and urogenital tract [56]. Typically, the group of microorganisms that inhabit one of these regions is referred to as microbiota (e.g., gut microbiota). Recently, it has been demonstrated that the microbiota is an important factor in maintaining homeostasis in healthy patients, and that an imbalance in microbiota composition (i.e., increased concentrations of pathogenic bacteria) can lead to disease [57]. For the purpose of this review, focus will be placed on discussing the oral and gut microbiota, as they are currently believed to be the most implicated in the pathogenesis of AD.
Oral biofilms: An important reservoir of infections
The oral cavity is believed to contain between 500–700 bacterial species, although less than half of these species can be cultivated in the laboratory [58 –60]. In recent years, there has been increasing evidence that oral bacteria are associated to the onset and maintenance of a wide range of systemic diseases, including cardiovascular disease, pneumonia, atherosclerosis, and infective endocarditis [61]. For example, oral biofilm on the surface of infected removable dentures can generate a local inflammatory disease known as denture-related stomatitis [62, 63]; however, it may also act as a reservoir for pathogens involved in respiratory diseases. Recently, O’Donnell et al. isolated six respiratory pathogens from denture surfaces, including S. pneumoniae, Pseudomonas aeruginosa (P. aeruginosa), and Haemophilus influenzae B (H. influenzae B) [64]. Furthermore, a study by Iinuma et al. demonstrated that in elderly patients, increased microbial load and inflammation is associated to higher risks of pneumonia [65]. Also, oral cariogenic bacteria such as Streptococcus mutans (S. mutans) have been isolated from infected heart valves [66, 67], and a study by Nakano et al. demonstrated the presence of S. mutans in both heart halves and atheroma plaques [68]. Therefore, bacteria are able to propagate from the oral cavity to other tissues and disseminate infection. How does this process occur? It is believed that bacteria from the oral cavity can enter into the bloodstream during procedures that generate transitory bacteremia such as dental treatment, tooth extraction, and tooth brushing [69 –71]. The most relevant oral biofilms involved in dissemination are therefore the ones closest to the vascularized tissues surrounding the tooth, such as the periodontal biofilm.
Periodontal disease is a pathology that causes the inflammation and destruction of the tissues surrounding teeth, such as the periodontal ligament, alveolar bone, gingiva, and tooth cementum [72]. Periodontal disease is initiated and maintained by a specific biofilm on the surface of the tooth, which are mostly comprised of Gram-negative anaerobic strains such as Prevotella intermedia (P. intermedia), Porphyromonas gingivalis (P. gingivalis), Aggregatibacter actinomycetemcomitans (A. actinomycetemcomitans), Fusobacterium nucleatum (F. nucleatum), and Treponema denticola (T. denticola). If left uncontrolled it can lead to tooth loss and potentially progress toward total edentulism [73].
Although the association between periodontal disease bacteria and systemic diseases is well known, the mechanisms underlying this process are still not fully understood. Overall, it is now believed that periodontal disease induces an inflammatory response that is not confined to the local site of infection. Instead, microorganisms and pro-inflammatory mediators are able to enter the bloodstream and affect other tissues and organs [74, 75] (Fig. 1). This process is facilitated by the direct contact between the periodontal ligament and vascular components, as well as the large area of inflamed gingival tissue in periodontal disease patients (believed to be between 8–20 cm2) [76]. Increased levels of systemic C-reactive protein, IL-6, IFN-γ, TNF-α, and IL-1β have been found in patients suffering from periodontal disease [77 –79], which can lead to endothelial dysfunction and altered immune responses [80]. Ultimately, this combination of bacterial infection and immune modulation originating from periodontal tissues is able to create a systemic response that may lead to chronic inflammatory diseases such as rheumatoid arthritis [81] (Fig. 2). Therefore, the control of oral and dental infections is currently an important strategy against the prevention of a broad range of systemic conditions [82 –84].

Periodontal disease as a source of systemic infection and inflammation. A) Periodontal disease is a chronic, infectious disease caused by the accumulation of a specific biofilm on the surface of the tooth that causes inflammation of the soft tissues surrounding the tooth and bone resorption. B) Biofilm accumulation generates a chronic immune response in the periodontal tissues releasing inflammatory mediators such as IL-6, IFN-γ, TNF-α, and IL-1. Other mediators such as RANKL are involved in osteoclastic activation and bone resorption. Bacteria from the periodontal biofilm, as well as lipopolysaccharide (LPS) and outer membrane vesicles (OMVs), are able to migrate into the bloodstream and affect distant tissues and organs.
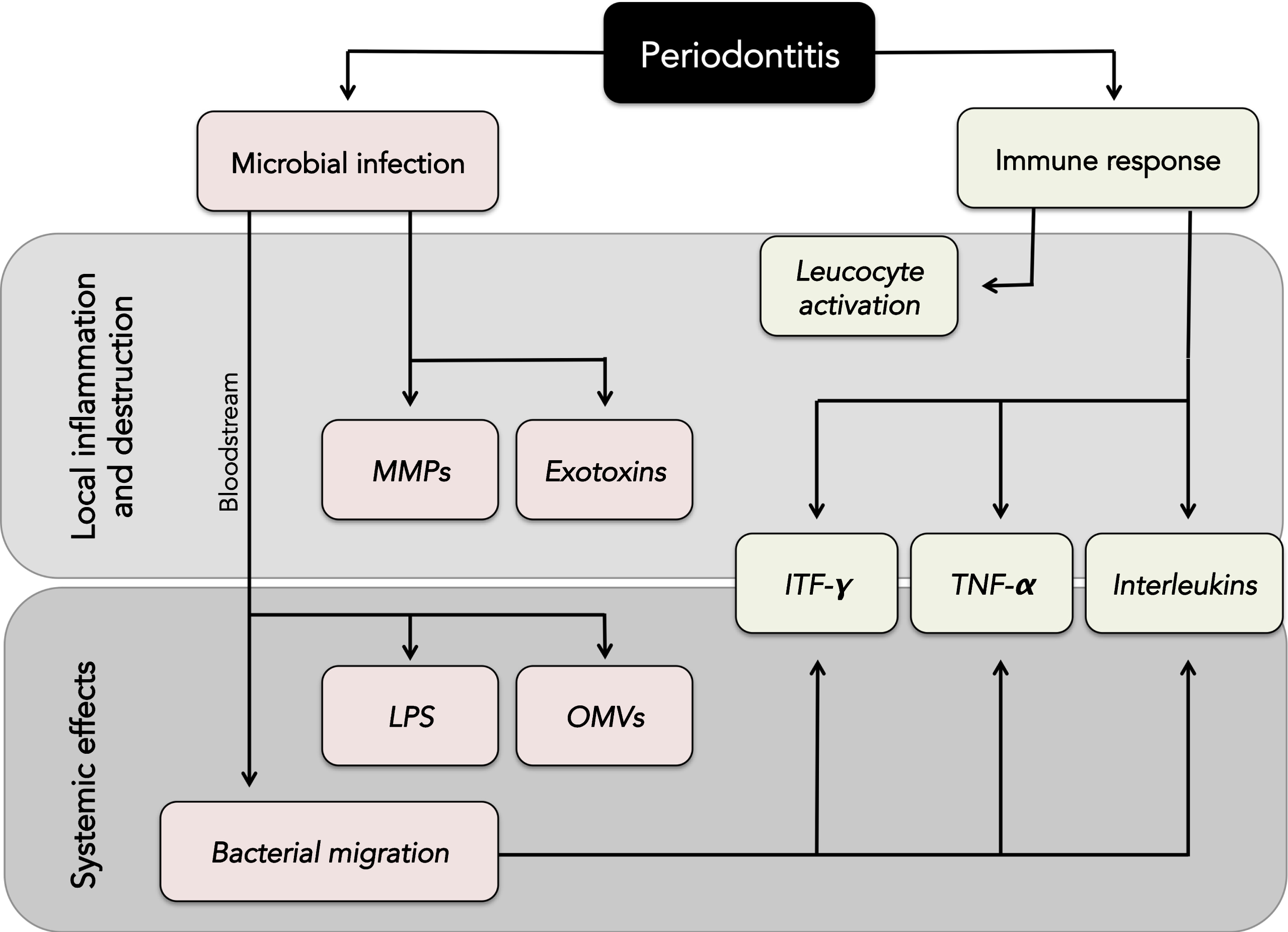
Overview of the local and systemic inflammatory effects of periodontal disease. MMPs, metalloproteinases; LPS, lipopolysaccharide; OMVs, outer membrane vesicles; IFN-γ, interferon gamma; TNF-α, tumor necrosis factor alpha.
Gut microbiota and chronic diseases
Trillions of microorganisms reside in the human gut where their composition varies due to nutritional factors, as well as chronic diseases such as inflammatory bowel syndrome and ulcerative colitis [85 –88]. They are of extreme importance for the host as they participate in nutrient digestion, influence gene expression, and regulate the mucosal immune system [89]. During early lifehood, the gut microbiota is determined by factors such as delivery method (vaginal delivery or caesarean) [90], cessation of breastfeeding [91], and introduction of complementary feeding [92]. Therefore, it is no surprise that alterations in the gut microbiota composition have been associated to several chronic diseases in humans, such as obesity, diabetes, and cancer (reviewed by [93]). Ley et al. observed that the ratios between Bacteroidetes and Firmicutes, two important taxa of bacteria, are significantly different between obese and lean humans and mice. The authors found that the relative proportions of Bacteroidetes are decreased in obese subjects when compared to lean controls [94, 95]. Furthermore, Ajslev et al. observed that early exposure to antibiotics during childhood (before 6 months of age) was associated with future obesity [96], which could be explained by the effect of antibiotic treatment on the establishment of gut microbiota at early stages of development. Also, the species Bilophila wadsworthia (B. wadsworthia) has been associated with bowel inflammation in rat models, which is probably a consequence of H2S production by this particular strain of bacteria [97]. Another important gut pathogen is Helicobacter pylori (H. pylori), which is a causative agent of stomach ulcer and gastric cancer. This is due to its capacity to locally harm mucosal epithelium via proteases and VacA cytotoxins [98], and also affect the immune response in the gastric tissue by production of TNF-α and IL-1β [99 –102].
EVIDENCE OF MICROBIAL INFECTION AS A FACTOR FOR AD DEVELOPMENT
Despite the abovementioned studies suggesting a link between microorganisms and a number of chronic diseases in humans, there is still no consensus regarding the potential effect that microbial infections may have on the initiation and progression of AD. Some early studies showed viral and bacterial strains, as well as parasites, in brains of AD patients; more recent experimental approaches, such as next generation sequencing of 16S ribosomal RNA, have demonstrated the presence of increased levels of bacteria in affected brain tissue [103]. Another study has also suggested the presence of poly-microbial infections in AD brain tissue [104]. Overall, these findings have strengthened the hypothesis that pathogens play an important role in the development of AD. This hypothesis is supported by research linking the presence of microorganisms with increased Aβ deposition, tau hyperphosphorylation, and glial cell inflammation and activation [105 –107]. Although the list of potential pathogens is growing, this review will focus on the strongest data regarding spirochetes, and oral and gut biofilms.
Spirochetes
One of the most relevant pathogens associated with the presence of AD are spirochetes, which are helical-shaped, highly motile Gram-negative bacteria involved in the pathogenesis of a wide range of human diseases [108, 109]. In a recent meta-analysis, Maheshwari and Eslick found a ten-fold increase of AD occurrence when spirochete infection was found in either the cerebral cortex or serum of patients [110]. Among these, species such as Treponema pallidum (T. pallidum), Borrelia burgdorferi (B. burgdorferi) and a number of dental spirochetes, which have shown the strongest associations to AD [43 , 112].
B. burgdorferi, the causative agent of Lyme disease, has been isolated from AD brains in several occasions [113, 114]. It is believed that B. burgdorferi may persist in tissues after chronic Lyme neuroborreliosis, and be associated with the onset of AD in a number of patients [115]. In vitro, Miklossy et al. found that upon exposure to B. burgdorferi, primary rat neurons developed morphological changes similar to the ones seen by Aβ deposition in AD, as well as increased levels of AβPP and hyperphosphorylated tau protein [116]. However, other groups have not been able to find evidence of this particular pathogen in brain tissue of AD patients, although these authors considered only B. burgdorferi in their research and ignored the predominant oral spirochetes [117, 118]. In an earlier study, Miklossy also found evidence of spirochetes in the brains of AD patients, which were not present in a group of paired healthy control samples [119]. Furthermore, Riviere et al. found strong evidence of six oral Treponema species (T. amylovorum, T. denticola, T. maltophilum, T. medium, T. pectinovorum, and T. socranskii) in brains of AD patients, supporting the idea that more than one species of spirochete may be involved in brain invasion. Analysis of the frontal cortex demonstrated oral Treponema invasion in 14 out of 16 AD brains, but only in 4 out of 18 non-AD patients [120]. Another study also showed that senile plaques from AD patients contained spirochete-specific DNA and that pure B. burgdorferi biofilms were able to produce both AβPP and Aβ, which suggests that senile plaques contain a combination of both host and bacterial produced Aβ [121].
T. pallidum, the spirochete responsible for syphilis and neurosyphilis, also has a well-known ability to colonize the brain and generate a disease known as general paresis, which is similar to AD in terms of symptoms and pathogenesis [122]. The similarities between AD and neurosyphilis have led to the assumption that both diseases may share a common bacterial background (reviewed in [42]). Although T. pallidum is sexually transmitted, it has been found in the brain of patients even at early stages of infection [123], and its accumulation over time leads to the development of general paresis, which shares many hallmarks of AD such as Aβ plaque deposition, NFT, progressive dementia and cortical atrophy [42, 124].
Oral bacteria and AD
There is a growing range of evidence strengthening the link between periodontal bacteria and AD. One of the most relevant species involved in periodontal disease is P. gingivalis [125]. Kamer et al. showed that AD patients present increased levels of TNF-α and antibodies against periodontal pathogens, including P. gingivalis, compared to normal controls [126]. In another recent study, Poole et al. demonstrated the presence of P. gingivalis derived lipopolysaccharide, an important bacterial endotoxin, in brain samples from AD patients [127]. This data is in line with previous work linking lipopolysaccharides with increased Aβ deposition [128, 129]. By comparing gene expression data of P. gingivalis to microarray datasets of the hippocampus of AD patients, Carter et al. demonstrated that upregulated genes in P. gingivalis-infected macrophages matched the ones found in AD brains [130]. They also suggested that P. gingivalis, as well as periodontitis, can influence AD either directly by inflammation, or indirectly by promoting other AD-related risk factors such as diabetes, atherosclerosis and hypertension [130]. Furthermore, Noble et al. utilized a logistic regression model to suggest that levels of P. gingivalis IgG are associated to cognitive decay in patients over 60 years of age [131]. This idea was explored further in an animal model demonstrating that ApoE– /– mice infected for 24 weeks with the oral pathogen P. gingivalis showed actual infections in the brain, and particularly in hippocampal neurons that are critical for memory formation [132]. This direct bacterial colonization in the brain is interesting since it suggests a direct effect of bacteria in the brain parenchyma, as opposed to a long-distance effect via inflammatory mediators. Alternatively, inflammatory factors such as LPS might also be involved by increasing the permeability of the blood-brain barrier (BBB) and thus facilitating bacterial penetration into the brain [133]. Under this setting, it is possible to consider that bacteria might induce local inflammation and release peptides that can act as porins.
In a subsequent study, it was reported that high serum IgG levels for Actinomyces naeslundii (A. naeslundii), another periodontal pathogen, was associated to increased risks for AD onset in patients over 65 years old [134]. Increased antibody levels for other important periodontal pathogens such as P. intermedia and F. nucleatum have also been found in patients with AD [135]. Interestingly, serum antibody levels for these bacteria were already elevated at baseline in the group that developed AD, even before cognitive impairment had occurred. This data suggests that periodontal disease and the presence of high serum antibody concentrations might occur prior to AD and may play a role as a causative factor. Another pathogen that has been associated to AD is E. faecalis, which is an oral pathogen implied in root canal infection and chronic periodontal disease [136], which has been shown to migrate into the brain and form abscesses [137]. Underly et al. found that E. faecalis infection increased expression of tau changes in primary rat cortical neurons, suggesting a potential link to AD pathogenesis [138]. The fact that AD seems to be associated to the presence of several periodontal bacteria instead of a unique species suggests that the bacterial factor may be non-specific, strengthening the idea that overall inflammation and infection play a part in the pathogenesis of AD.
Finally, it is important to consider the associations found between tooth loss and the development of AD. In a recent study, Batty et al. found that tooth loss was associated with increased risk of dementia in a group of 55–88 year old adults [139]. Similarly, in a 13-year long longitudinal study, Li et al. observed that tooth loss was associated to cognitive decline in elderly Chinese patients [140]. It is important to note that in both studies the number of teeth was self-reported by the patients and not confirmed by clinical examination. However, a study by Steward found no association between tooth loss and development of dementia, although they did find an association between dementia and gum inflammation [141]. It is necessary to emphasize that there are many potential causes for missing teeth (i.e., trauma, dental cavities) and therefore, the parameter of tooth loss may not be the optimal indicator for periodontal disease or the levels of periodontal bacteria in the oral cavity [142].
Gut microbiota
Among gut microorganisms, H. pylori appears to be one of the most relevant pathogens associated with AD. This is particularly worrying as it is estimated that 4.4 billion people are infected with H. pylori worldwide [143]. In a clinical study, Kountouras found that H. pylori was found in 88% of AD patients versus 47% of age-matched controls at baseline, and eradication of H. pylori improved cognitive parameters in the AD group [144]. In a recent publication, Felice demonstrated that an important H. pylori peptide modulates a large number of genes, some of which are directly related to AD [145]. The peptide, Hp(2-20), was found to alter the expression of 77 AD genes, 30 of which modulate inflammatory pathways. The specific mechanisms behind H. pylori pathogenesis in AD remains unknown, although it may be related to increased production of pro-inflammatory mediators such as TNF-α, INF-γ, and several interleukins [146], all of which have been shown to be important in AD pathogenesis [147].
Despite the apparent importance of H. pylori, there are other potential gut microbes believed to also play a role in AD. In a recent paper by Minter et al. mice that underwent long-term antibiotic treatment showed significant reduction in Aβ plaque deposition, which was correlated to a shift in the composition of the overall gut microbiota. This work is interesting as it demonstrates that gut microbiota can be altered to prevent the progression of AD pathogenesis [148]. Also, Liang et al. showed that treatment with probiotic Lactobacillus helveticus NS8 improved cognitive functions in rats subjected to stress [149]. Furthermore, Wang et al. demonstrated that treatment of rats with ampicillin for a month disrupted their gut microbiota and led to symptoms such as increased anxiety and spatial memory impairment, but further treatment with Lactobacillus fermentum NS9 reverted these effects [150]. Many other animal studies have also demonstrated an association between the administration of probiotic bacteria and improved cognitive function [151 –153]. A recent clinical study showed that Bifidobacterium longum 1714 improved visuospatial memory performance and reduced stress in 22 healthy patients [154], suggesting that specific strains of microorganisms have the ability of directly modulating brain function in humans. Furthermore, recent data has shown that strains such as E. coli and Salmonella enterica have the ability to secrete bacterial Aβ, which is used to promote biofilm stability as it is resistant to denaturing and degrading conditions (reviewed in [155 –158]); however, the effect of this bacterial-derived Aβ on the host is not yet understood.
Another topic of interest is the way in which gut microbiota may influence the brain by means of the gut-brain microbiota axis [159]. It has been shown that alterations of this microbiota can lead to changes in behavior, and are implied in the regulation of anxiety, pain, obesity, and autism [160, 161]. It is believed that the gut-brain microbiota axis is not influenced by direct bacterial infection of the brain, but via the homeostatic control of innate immunity [162]. Furthermore, disturbances in the gut epithelium barrier, or in other factors such as mucin production, may increase intestinal permeability and allow the passage of antigens and pathogens into the host (reviewed in [163]). This increased permeability has been denominated ‘leaky gut’, and it has been associated to multiple autoimmune diseases such as celiac disease, lupus erythematosus and diabetes type I [164, 165]. The residing microbiota is important in regulating the function of the epithelium layer and plays a key part in the maturation and maintenance of the intestinal barrier [166 –168]. Interestingly, a recent study by Braniste et al. demonstrated that the gut microbiota also has a direct impact on the permeability of the BBB because germ-free mice developed increased BBB permeability that was reduced once the gut microbiota was restored [169]. Increased permeability in both the intestinal barrier and BBB would provide easy passage of microorganisms and bacterial toxins not only into the bloodstream, but also into the brain and contribute to the pathogenesis of AD.
Other relevant microorganisms
Besides the abovementioned bacteria, there is also evidence suggesting that other microorganisms may be implied in the pathogenesis of AD. Recent work by Pisa et al. demonstrated the presence of several strains of fungi in brain tissue of 10 AD patients, whereas no fungal cells were found in the control group [170]. Also, Alonso et al. found evidence of increased serum levels of fungal polysaccharides and disseminated fungal infection in AD patients [171]. Interestingly, cases of infection by the fungus Cryptococcus neoformans have been clinically mistaken for AD, and in these cases dementia was reversed after proper antimycotic treatment [172, 173]. Furthermore, the fact that Aβ has shown to be antimicrobial against C. albicans supports the association between fungal infection and AD pathogenesis [33]. Also, it is important to mention that infection with HIV can lead to HIV-associated dementia, characterized by the infection of macrophages, microglia and astrocytes in the brain [174]. These infected cells shed viral proteins such as Tat and gp120, which in turn promote neuroinflammation by the production of inflammatory cytokines, as well as glutamate accumulation due to the down-regulation of the EAAT2 glutamate transporter in astrocytes [175 –177].
POTENTIAL LINK BETWEEN MICROBES AND Aβ: PORE FORMATION IN CELL MEMBRANES
In the microbe world, ion channels are critical for controlling cellular homeostasis and defense. Porins, for example, are outer membrane proteins found in many Gram-negative bacteria that allow the passage of small solutes important for various metabolic processes (<650 Da) [178]. Interestingly, like Aβ, porins present a secondary structure having predominantly β sheets that can form complex association with at least trimeric conformation [179 –182]. The oral Gram-negative anaerobe F. nucleatum is of particular interest since it not only causes periodontal disease but also can produce clinical infections in other body sites [183]. A recent study showed that people that developed cognitive impairment presented higher levels of serum antibodies for F. nucleatum [135]. However, it is unknown if the impairment was related to the presence of the bacteria in the brain. This oral bacterium is also interesting because it expresses a protein in the outer membrane, known as FomA, that functions as a non-specific porin in lipid bilayer membranes [184]. Furthermore, Zilm et al. described an upregulation of SSP 3802 (belonging to the Omp IP family of porins) in F. nucleatum as a response to a pH increase from 7.2 to 7.8, which is believed to be the pH of diseased periodontal sulcus [181]. Neisseria meningitidis expresses two major outer membrane porins, PorA and PorB, while Neisseria gonorrhoeae only expresses PorB, which is found to play multiple roles during infection [186, 187]. Novak and Cohen found that P. gingivalis are able to induce membrane depolarization of polymorphonuclear leukocytes following bacteria-cell contact, which is consistent with pore formation or activation [188]. Some studies have demonstrated that porins can translocate from bacteria to foreign membranes [189, 190], which suggests a potential mechanism for bacterial-induced cell death. Thus, it is possible that bacteria in the brain cause neuronal membrane leakage producing Aβ movement from/to the neuron, initiating a process of increased damage now by the toxic Aβ peptide.
Ion channel formation in microorganisms also serves a defensive role when released to the extracellular medium. Gramicidin, a classical ion channel forming antibiotic is secreted by Bacillus brevis and it corresponds to a hydrophobic pentadecapeptide, consisting of an alternating sequence of D- and L-amino acids forming a β-bonded helix in a hydrophobic environment (reviewed in [191]). This property leads to both ion channel formation and anti-microbial activity against other strains of bacteria [192, 193]. Similarly, nystatin derived from Streptomyces noursei is a polyene antifungal agent which in presence of ergosterol forms channels in the cell membrane disrupting the normal cellular environment and ultimately leading to cell death [194].
Furthermore, porin-like membrane proteins have been described in relevant strains of Treponema, as well as certain important periodontal pathogens. T. pallidum expresses TprC/D and TprI as outer membrane proteins, both of which have been identified to display porin-like activity [195]. Furthermore, T. denticola has been found to possess a 53 kDa surface antigen (Msp) on its outer sheath that demonstrated dual adhesin and porin activity [196, 197]. Outer membrane vesicles of Eikenella corrodens, another periodontal pathogen, displayed porin-like properties and induced cytotoxicity against mouse macrophages [198]. It is possible that the above-mentioned ion channel-forming peptides and the complex molecules normally found in the microbial environment are important in the development of AD. Similar to the Aβ peptides, these microbial-derived peptides might be causing membrane damage and ion dyshomeostasis leading to neuronal death. As discussed throughout this review, porin producing bacteria such as T. pallidum, T. denticola, and F. nucleatum are frequently observed in the brains of AD patients. Also, Aβ was reported to have antimicrobial activity against several bacteria [33], and therefore could be upregulated as a response to bacteria and bacterial toxins in the brain.
HOW COULD BACTERIAL OUTER MEMBRANE VESICLES CONTRIBUTE TO PORIN FORMATION?
Almost every cell (mammalian, bacterial, plant) has the ability to secrete extracellular vesicles, and until recently, it has been considered to be mainly a cellular waste system. However, it was found that extracellular vesicles can carry a variety of cargos such as DNA, RNA, proteins and lipids [199], and play key roles in cellular communication, even between kingdoms [200]. Bacteria shed periplasmic fluid containing extracellular vesicles or outer membrane vesicles (OMVs), which are 20 nm to 250 nm in size, in an active process during cell growth [201].
Over the last years, OMVs have become increasingly interesting in the study of both pathology and regeneration. Since OMVs are derived from the bacterial membrane, they also carry specific active membrane proteins. Several studies have shown the involvement of β-lactamase carrying OMVs in antibiotic resistance [202, 203] and also delivery of bacterial virulence factors by OMVs in bacteria such as E. coli [204], P. aeruginosa [205], and P. gingivalis [206]. Also, OMVs— especially ones derived from P. gingivalis— have been shown to modulate immune responses. One mechanism involves gingipain, an endopeptidase involved in nutrient collection. In the initial phase of P. gingivalis infection, gingipain activates the C1 complex, which creates a local inflammatory response improving nutrient supply and colonization. In later phases, with increasing presence in and around the biofilm, gingipain has the ability to inactivate complement factors (C3, C4, C5) leading to resistance to the complement system [207]. Furthermore, P. gingivalis [208], H. pylori [209], and P. aeruginosa [210] share an interesting trait: they are not only porin-forming bacteria, but they have been shown to produce outer membrane vesicles that enter mammalian cells via lipid rafts [205 , 212]. This is interesting since Malchiodi-Albeli et al. demonstrated that disruption of lipid rafts protects neurons from amyloid oligomer-induced toxicity [213], suggesting a similar entry route of both amyloid and porin-containing OMVs. Therefore, it is possible that bacterial OMVs could be a key factor regarding neuronal death, although further research is necessary in order to understand this potential association.
CONCLUSIONS
As discussed in this review, there is a broad amount of research demonstrating an association between AD and infectious processes such as periodontal disease [214]. This association is not limited to a specific type of microorganism, but rather to a wide range of bacteria, fungi, and viruses. This suggests that instead of being associated to a particular individual pathogen, the mechanisms behind the development of AD are more general and may be mostly derived from generalized microbial injury and its consequent chronic inflammatory response within the brain tissue. This microbial effect is probably a combination of both direct damage caused by the migration of microorganisms into the CNS due to increased BBB permeability in older patients [215], and an indirect effect mediated by the systemic release of bacterial toxins, OMVs and pro-inflammatory molecules into the bloodstream. For example, Treponema species can potentially migrate to the brain from the oral cavity (where around 60 species of Treponema have been identified, many of them invasive [216]), and cause local damage due to a combination of porin production and generalized neuroinflammation. In either case, the presence of microbial components in the brain may potentiate Aβ plaque deposition and generate neuronal dysfunction by the action of porin-like molecules that increase membrane permeability and calcium leakage.
From the above, it is necessary to begin redefining the current understanding of AD to include a microbial component (Fig. 3). This does not mean that the genetic background or other potential environmental factors should not be considered. However, several studies have provided evidence supporting the idea that infection might be related to AD pathogenesis, and further efforts should be placed on exploring this possibility [214]. Until recently, diseases such as gastric cancer were believed to be non-infectious, but nowadays they are known to be associated to pathogens. Thanks to this discovery, in the last decades antibacterial treatment against H. pylori has reduced the incidence of gastric cancer in the population [217]. In the same way, it is possible that novel therapeutic approaches to prevent and treat AD could include antibacterial management in the future. In any case, the importance of maintaining control of biofilm-mediated diseases must be strongly highlighted. In summary, the reviewed data suggests that the link between bacteria and AD resides in the capacity of the bacteria to act directly and indirectly on the brain, and future research should be encouraged in order to further explore the specific mechanisms behind the infectious component of AD.

Proposed link between bacterial infection and Aβ in AD pathogenesis. Bacteria and released components (such as LPS and OMVs) can enter brain tissue and generate a local immune response that initiates plaque deposition and tau hyperphosphorylation, as well as upregulate inflammatory mediators such as TNF-α and IL-1β that potentiate neuroinflammation. Furthermore, bacteria could directly damage neurons by the release and incorporation of porin-like proteins with the potential of causing unregulated calcium entry, synaptic toxicity and neuronal death. The pore-induced membrane damage could act synergistically with the Aβ induced pore formation to enhance neurotoxicity. Alternatively, Aβ could be released by neurons and act as an antibacterial agent because of its pore forming action.