
Abstract
Alzheimer’s disease (AD) is a neurodegenerative pathology, which is characterized by progressive and irreversible cognitive impairment. Most of the neuronal perturbations described in AD can be associated with soluble amyloid– β oligomers (SO-Aβ). There is a large amount of evidence demonstrating the neuroprotective effect of Nicotine neurotransmission in AD, mainly through nicotinic acetylcholine receptor (nAChR) activation and antiapoptotic PI3K/Akt/Bcl–2 pathway signaling. Using HPLC and GC/MS, we isolated and characterized two alkaloids obtained from C. scoparius, Lupanine (Lup), and 17– oxo-sparteine (17– ox), and examined their neuroprotective properties in a cellular model of SO-Aβ toxicity. Our results showed that Lup and 17– ox (both at 0.03μM) prevented SO-Aβ-induced toxicity in PC12 cells (Lup: 64±7%; 17– ox: 57±6%). Similar results were seen in hippocampal neurons where these alkaloids prevented SO-Aβ neurotoxicity (Lup: 57±2%; 17– ox: 52±3%) and increased the frequency of spontaneous calcium transients (Lup: 60±4%; 17– Ox: 40±3%), suggesting an enhancing effect on neural network activity and synaptic activity potentiation. All of the neuroprotective effects elicited by both alkaloids were completely blocked by α-bungarotoxin. Additionally, we observed that the presence of both Lup and 17– ox increased Akt phosphorylation levels (52±4% and 35±7%, respectively) in cells treated with SO-Aβ (3 h). Taken together, our results suggest that the activation of nAChR by Lup and 17– ox induces neuroprotection in different cellular models, and appears to be an interesting target for the development of new pharmacological tools and strategies against AD.
INTRODUCTION
Alzheimer’s disease (AD) is the most prevalent type of dementia, and it is characterized by the progression of symptoms like the loss of brain cognitive functions, especially memory and rationale behavior [1]. Strong experimental evidence supports the idea that soluble oligomers of amyloid-β (SO-Aβ) can have a major impact on the neuronal alterations described in AD development [2]. A classical experiment performed by Lesne et al. showed that brain injection of Aβ oligomers (56 KDa) in young rats negatively affected memory without the necessary presence of plaques [3], reinforcing the idea that these species could be the cause of synaptic activity alterations and neuronal death in vitro [4]. We and other groups have demonstrated that SO-Aβ can form a pore in the neuronal membrane, affecting its ionic homeostasis and inducing chronic synaptic failure, ATP depletion, and metabolic consequences leading to AD development [4–7]. Early in the 1980 s, the idea that the cognitive deficit characteristic of AD was related with a deficiency in cholinergic transmission, known as the cholinergic hypothesis [8]. One of the most used therapeutic approaches to treat AD is based on the strategy of enhancing the cholinergic tone in the CNS, for example through the use of galantamine and rivastigmine [9]. The idea that cholinergic neurotransmission is a key target for the development of new pharmacological tools that could enhance memory and learning functions, is still an interesting strategy to treat AD. This cholinergic hypothesis proposes that cholinergic system alterations contribute substantially to the etiology of AD, and has been updated based on new evidence that propose different actions related to the activation of nicotinic acetylcholine receptors (nAChR) in neurons as well as in glial cells [10–12]. Several studies show that there is an increase in cognitive functions when cholinergic systems are modulated [13]. For example, through the modulation of α7 and α4β2, the main and most studied nAChRs in the brain [14], several studies have demonstrated that α7 nAChR is involved in neuroprotection through the activation of the PI3K/Akt/Bcl–2 survival pathway [15–19]. Moreover, Sadigh-Eteghad et al. found that the α7 specific agonist PHA–543613 improves recognition memory in Aβ25–35 intraventricularly injected mice [20]. Accordingly, Callahan et al. reported that the partial α7 and α4β2 agonist Tropisetron improves memory in young and aged rats, and in aged Rhesus monkeys [21]. Classically, the most important compounds/metabolites that act on nAChRs come from natural sources (e.g., plants, fungus) and share the characteristic of being part of a wide family of molecules that have different and complex chemical structures: the alkaloids. The most common neuroactive natural compound used to activate these kind of receptors is the alkaloid Nicotine. However, other important alkaloids have been used such as epibatidine and cytisine, the latter being a very important pharmacological tool [22] and was found to have protective properties on excitotoxicity models induced by NMDA toxicity [23]. Recently, we isolated and characterized the neuroprotective properties of two quinolizidinic alkaloids obtained from Teline monspessulana (L) and described their activity against Aβ toxicity in cellular models [24].
MATERIALS AND METHODS
Reagents
All the reagents were P.A. or HPLC grade. Acetonitrile, hydrochloric acid, ethylacetate, and chloroform (Merck Chemicals, Darmstadt, Germany).The water used was from MilliQ system from Millipore (Milford, Mass, USA). The mobile phase was filtered through a Whatman membrane filter (47 mm diameter and 0.2μm pore size) while all the plant extracts were filtered through Acrodisc syringe filter (PTFE membrane with 0.22μm pore size).
Alkaloids obtaining and identification
Cytisus scoparius was collected in the city of Coronel (Biobío Region) during the months of June and July of 2014, and identified by Professor Roberto Rodríguez, Botanic Department, University of Concepción, Concepción, Chile. A voucher specimen was deposited in the Herbarium University of Concepción (CONC N° 152943). Subsequently, the dried aerial parts (leaves and stems) of the C. scoparius were isolated and prepared for the alkaloid extraction as follows: The air-dried aerial parts of the plant were cut into small pieces and were extracted with 99% methanol three times at room temperature. The combined MeOH extracts were concentrated under vacuum, acidified with 5% hydrochloric acid to pH 2–3, and then extracted with diethyl ether three times. The pH of aqueous fraction was alkalinized with 25% ammonium hydroxide to pH 9–10 and extracted exhaustively with chloroform o yield the basic CHCl3 extract.
The alkaloid extract was fractionated through a chromatographic column (CC, 100 cm×7 cm) that contain Merck 60 silica gel (0.032–0.063 mm) as stationary phase, and chloroform-methanol (CHCl3:MeOH) as mobile phase, with growing polarity (99:1 to 1:99). The next step was to check the fractions by thin layer chromatography (TLC), using TLC Silica Gel 60 F254 Plate (Merck Chemicals, Darmstadt, Germany) and Draggendorf reactive. Using a second column (CC, 30 cm×1.5 cm) with the same stationary phase and mobile phase described previously.
Preparative HPLC
With the purpose to isolate and purify compounds, the fractions were isolated through High Performance Liquid Chromatograph (HPLC) coupled to a Diode Array Detector (DAD). The HPLC system conformed by a binary pump YL9111 s coupled with a DAD YL9160 PDA (Younglin Instrument Co. Ltd., Anyang, Korea). The separation was carried out using as stationary phase Kromasil ® 60–5– SIL, 250×100 mm at room temperature. As eluent was used an isocratic mix of n-Hexane: Chloroform (80:20) at pH 9 using NH4OH 25% v/v. The flow rate was set at 5 mL/min for 20 min. The monitoring and detection of the alkaloids was performed by scanning at 254 and 280 nm simultaneously. YL-Clarity version 4.0.3.797 software was used for recording and processing chromatographic data.
GC/MS analysis
The GC/MS analysis of underivatized alkaloids was carried out on a gas chromatograph (Agilent 7890A) with a splitless injector (250°C) and a mass detector (Agilent 5975C). A HP–5MS capillary column (30 m×0.25 mm×0.25μm) and helium gas (constant flow 1.4 mL/min) were used for separation. The temperature program was: 5 min hold at 100°C, 100–275°C at 13°C/min and 32 min hold at 275°C. The ionization voltage was 70 eV. The detection range was m/z 50–550 Dalton (Da). The injector temperature was 250°C. The identification of the alkaloid components was accomplished by matching their mass spectra with those recorded in the NIST 05 (NIST /EPA/NIH MASS 2005 Spectral Library) and by comparing the obtained spectra with those reported in the literature. NIST 05 collects representative alkaloids of all the skeletons referenced. The structure of an alkaloid is assigned when the overlap is above 90% with the database. The skeleton of mostly of unidentified compounds was proposed according to the highest index of similarity to any of those listed in the database. The percentage of the compounds in the extracts was calculated in basis of the total area of the GC/MS peaks [25].
Primary neuronal hippocampal cultures
Pregnant C57BL/6 mice were treated in accordance with the regulations recommended by NIH and the Ethics Committee at the University of Concepcion. Mice were deeply anesthetized by CO2 inhalation before being sacrificed by cervical dislocation. Primary cultures of embryonic hippocampi (E18) were plated at 320,000 cells/ml on coverslips coated with poly-L-lysine (Trevigen, Gaithersburg, MD, USA) in minimal essential medium (MEM; Gibco, Grand Island, NY, USA) supplemented with 10% horse serum, 4 mg/ml DNAse, and 2 mM L-glutamine for 24 h. Culture medium was replaced after 24 h with MEM (Gibco, Grand Island, NY, USA), 2% horse serum, 2% fetal bovine serum and 0.5% N3 (BSA 1 mg/ml, putrescine 4 mg/ml, insulin 1.25 mg/ml, sodium selenite 1μg/ml, TH3 2μg/ml, progesterone 1.25μg/ml, corticosterone 4μg/ml). The cell cultures were maintained at 37°C with 5% CO2. Experiments were performed at 10–11 DIV in control and treated neurons.
PC–12 cells
PC12 cells from ATCC (Manassas, VA, USA) were cultured in DMEM with 5% fetal bovine serum, 100 U/ml penicillin, 100μg/ml streptomycin, and 2 mM L-glutamine. The cells were incubated under standard conditions (37°C, 5% CO2) and when 80% confluence was achieved, the cells were treated with 0.25% trypsin for 10 min, washed and resuspended in HyQ DMEM/High-Glucose (Hyclone, Logan, UT, USA) with 5% fetal bovine serum (Hyclone, Logan, UT, USA), 2 mM L-glutamine (Gibco, Grand Island, NY, USA), and 1% penicillin-streptomycin (Gibco, Grand Island, NY, USA). The cells were then plated at a concentration of 50,000 cells/well for experiments and used 24 h after plating under experimental conditions similar for neurons.
HEK293T cells
HEK293T cells (ATCC, Manassas, VA, USA) were cultured using standard methodologies and kept in a thermo-regulated incubator at 37°C and 5% CO2. To evaluate viability, cells were platted at a concentration of 30,000 cells/well in a 96–well plate in DMEM supplemented with 10 % fetal bovine serum and examined on a Novostar multiplate reader (BMG Labtech, Offenburg, Germany) after the cells reached 80% confluence.
Soluble oligomers of Aβ (SO- Aβ) preparation
Aβ1– 40 peptide (rPeptide, Bogart, GA, USA) was reconstituted in DMSO at a concentration of 2.3 mM. 2μL aliquots were dissolved in sterile distilled water to 80μM concentration. To generate the oligomeric forms, the solution was subjected to vertical stirring (500 rpm) using a magnetic agitator for 2 h at room temperature as previously described [7, 26]. The presence of SO-Aβ was routinely tested by western blot (not shown). This peptide solution was used at a concentration of 0.5μM in all experiments.
Cell viability assay
We used the in vitro MTT assay kit (Sigma-Aldrich, St. Louis, MO, USA) to evaluate changes in cell viability by measuring the ability of mitochondria to reduce 3–[4, 5- dimethylthiazol–2– yl] –2,5– diphenyl tetrazolium bromide (MTT salt) to formazan. The cells subjected to different experimental conditions were incubated for 30 min in MTT (1 mg/ml). The insoluble formazan was solubilized in 100μl of 2– propanol, and the absorbance was read (560 nm and 620 nm) in a NOVOstar multiplate reader (BMG Labtech, Offenburg, Germany).
Spontaneous Ca2 + transients
Hippocampal cultures were loaded with the non-ratiometric Ca2 + sensitive fluorescent probe Fluo–4 AM (Invitrogen, Carlsbad, CA, USA) for 20 min in PBS using standard incubation conditions. Subsequently, the cells were washed 20 min with PBS and finally washed 3 times with normal external solution and then mounted on an inverted microscope. Changes in fluorescence (ex 480 nm, em 520 nm, 200 ms exposure) were acquired every 1 s for 10 min using an iXon+EMCCD camera (Andor, Ireland) and the Imaging Workbench 6.0 software (Indec Biosystems, Santa Clara, CA, USA). Spontaneous Ca2 + transients were measured after a 24 h incubation with SO-Aβ (0.5μM), 17– ox and Lup.
Western blot
PC–12 protein lysates were subjected to SDS-PAGE and transferred to nitrocellulose membranes (250 mA, 100 min) that were subsequently blocked with 5% non-fat milk in TBS-Tween. The primary antibodies used were anti p-Akt/Akt (rabbit, 1:1000, SantaCruz Biotechnology, Dallas, TX, USA) and anti β-actin (mouse, 1:2000, Santa Cruz Biotechnology, Dallas, TX, USA). Anti-mouse-HRP (1:5000, Santa Cruz Biotechnology, Dallas, TX, USA) and anti-rabbit-HRP (1:5000, Santa Cruz Biotechnology, Dallas, TX, USA) were used as secondary antibodies. Immunoreactive bands were exposed using ClarityTM Western ECL substrate (Bio-Rad Laboratories, Inc., Hercules, CA, USA) and quantified using an Odyssey FC detection system (Li-Cor, Lincoln, NE, USA).
Statistical analysis
Data in graphics are expressed as mean±SEM. Statistical analysis was performed using one-way ANOVA. The following results were considered statistically significant: *p < 0.05, **p < 0.01, and ***p < 0.001 versus control;
RESULTS
Chemical characterization of the purified alkaloids 17– oxo-sparteine and Lupanine
The specimens of Cytisus scoparius were obtained from a southern region in Chile (see materials and methods). The alkaloids were initially obtained from a crude extract of C. scoparius. 360 fractions (Fr) were obtained at the beginning, and fractions number 4 (Fr–4) and number 13 (Fr–13) were positive for the presence of alkaloids (not shown). Both fractions were purified independently of each one by HPLC (see Materials and Methods). From Fr–4 Lupanine (Lup, inset Fig. 1A) was identified by GC/MS showing a retention time of 16.638 min and 98.68% purity (Fig. 1A), with a molecular mass of 248 Da (Fig. 1B). On the other hand, from Fr–13, we identified 17–oxo-sparteine (17–ox) with a retention time of 16.560 min and 93.33% purity (Fig. 2A), while its molecular mass was 248 Da (Fig. 2B). Taken together, and correlating the experimental data with our compound library, we confirmed that the structures and the chemical nature of the molecules isolated were Lupanine (inset Fig. 1A), and 17–oxo-sparteine (inset Fig. 2A).

Chemical characterization of Lupanine (Lup). A) Chromatogram obtained by GC/MS and chemical structure of lupanine. B) Mass spectrum of Lup.
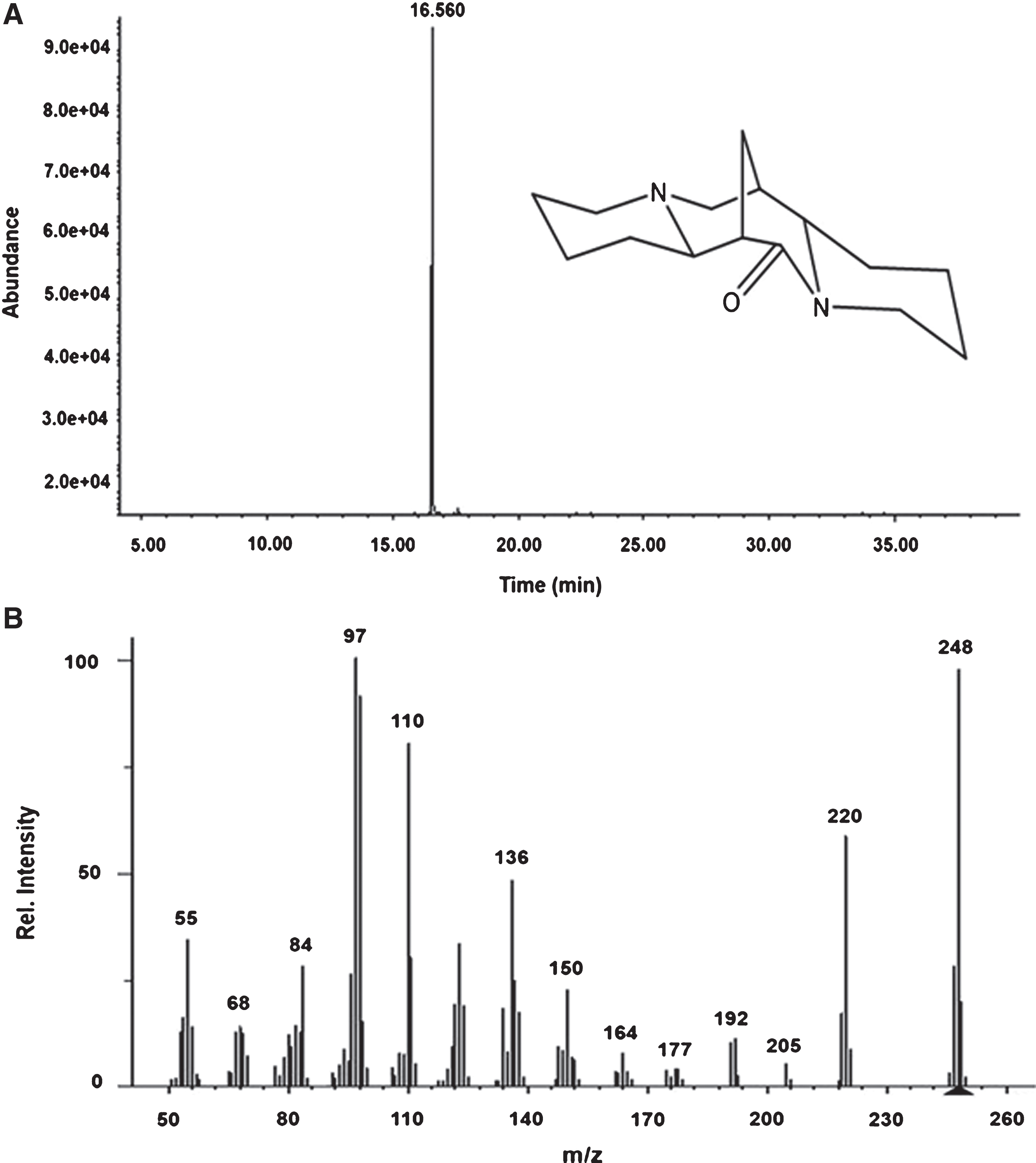
Chemical characterization of 17–oxo-spartein (17–ox). A) Chromatogram obtained by GC/MS and chemical structure of 17–oxo–spartein. B) Mass spectrum of 17–ox.
These two compounds were identified based on their spectral data (GC/MS). The mass spectrum of compound Lupanine (C15H24N2O,Mw = 248 g/mol) showed a molecular ion [M]+ at m/z 248 (46 %). A base at m/z 247 (32%) corresponding of [M - H]+, another peak at m/z 219 (11.47 %) indicating the loss of 28 units of mass [M- CO]+. The predominant ions at m/z 150 (36%), 149 (51%), 136 (100%), 134 (20%), 110 (19%), 98 (20%), 84 (10.65%), 55 (35%), and m/z 41 (9.83%) are characteristic of lupanine. The mass spectrum of compound 17–oxo-sparteine (C15H24N2O, Mw = 248 g/mol) showed a molecular ion [M]+ at m/z 248 (97%). A base at m/z 247 (28%) corresponding of [M - H]+, another peak at m/z 220 (58%) indicating the loss of 28 units of mass [M- CO]+. The predominant ions at m/z 98 (91%), 110 (51%), 97 (100%), 136 (48%), and m/z 55 (34%) are characteristic of 17–oxo-sparteine [27].
We then proceeded to examine both of these alkaloids in order to characterize their biological effects on cellular and neuronal models of toxicity induced by SO-Aβ), focusing on their neuroprotective properties through modulation of nicotinic receptors.
Chronic SO-Aβ toxicity and preventive effects induced by 17– ox and Lup
In order to discard intrinsic toxicity of the alkaloids, 17– ox and Lup were tested at a wide range of concentrations (0.0003– 30μM) and neither alkaloid displayed toxicity at any of the concentrations studied (data not shown). After that, PC–12 cells that express nicotinic receptors [28, 29] were used to evaluate the protective effects of 17– ox and Lup. Treatment of the cells with SO-Aβ (0.5μM) during 24 h reduced cell viability by about 48±5%, while co-incubation with 17– ox (Fig. 3A) or Lup (Fig. 3B), at different concentrations (0.0003–30μM), was able to prevent SO-Aβ toxicity; FCCP (10μM) was used as a positive control of the technique for toxicity. While the observed toxicity induced by chronic exposure to SO-Aβ has been described by our laboratory and others, the protective effects of these two alkaloids have not been reported previously, and is interesting to observe that the maximal effects of 17– ox (109±6%) and Lup (116±7%) were obtained in the sub-micromolar concentration range, suggesting a high potency of these two alkaloids to induce protection against SO-Aβ toxicity.
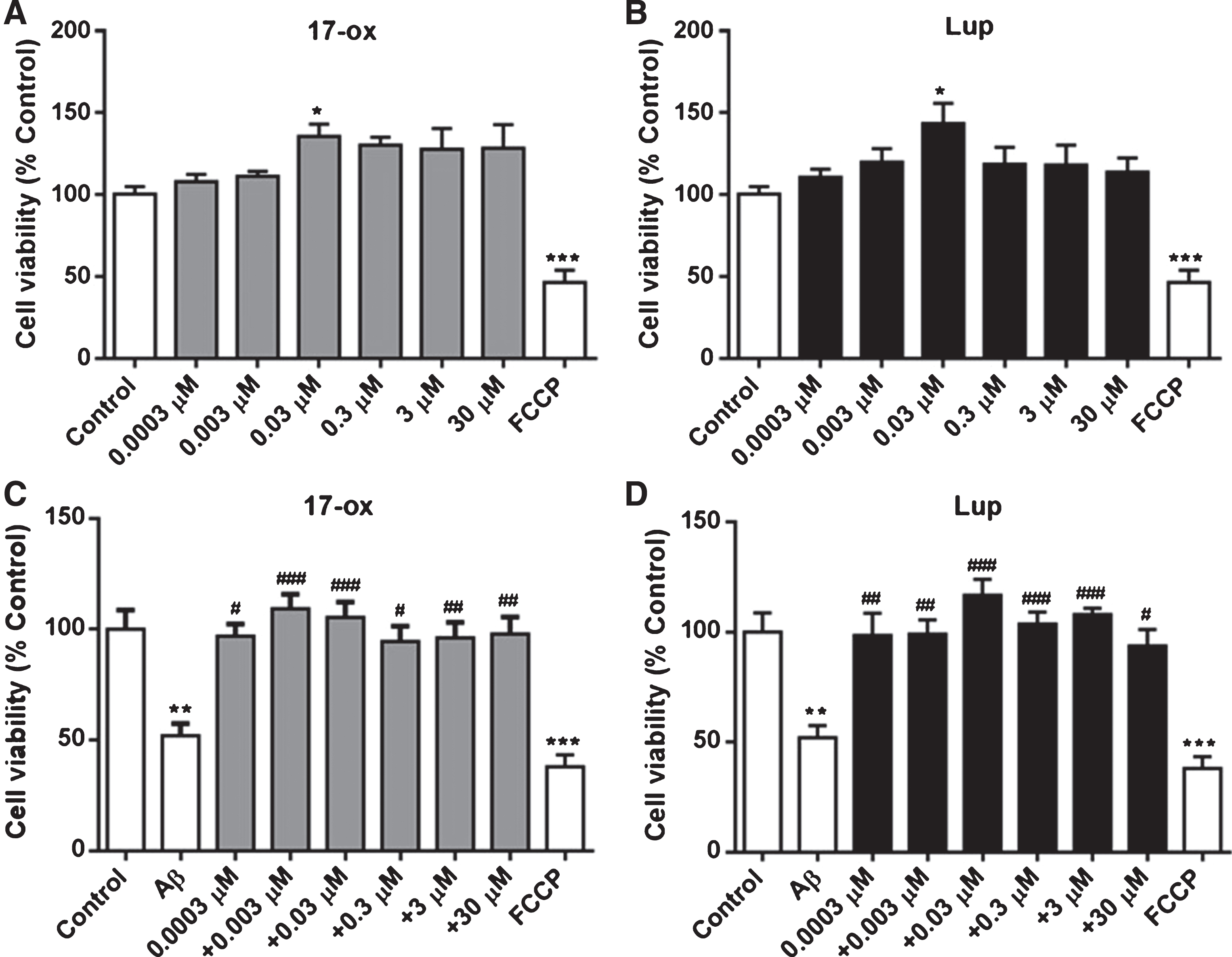
Effects of 17– ox and Lup on viability of PC–12 cells treated chronically with SO-Aβ. A) Effects of 17–ox (0.3–30μM) on cell viability using MTT techniques. B) Effects of Lup (0.03–30μM) on cell viability using MTT techniques. C) Effects of the toxicity induced by SO-Aβ (0.5μM, 24 h) and the effects of 17– ox (0.3–30μM) on cell viability. D) Effects of Lup (0.03–30μM) under similar conditions to C. FCCP (10μM) was used as a control for toxicity. The values are represented as percentage of control without treatment (n = 3; N = 9). **p < 0.01 and ***p < 0.001 versus control; #p < 0.05, # #p < 0.01, and # # #p < 0.001 versus Aβ.
17– ox and Lup prevented Ca2 + signal decay and synaptic failure induced by SO-Aβ in mice hippocampal neurons
Synaptic neurotransmission, which involve vesicle fusion and neurotransmitter release, is an exocytotic process that is modulated by transient cytosolic Ca2 + increase in the neuron. These transient increments reflect the spontaneous activity of the neuronal network and have a correlation with synaptic activity. Using microfluorimetry techniques, we determined that the normal frequency of cytosolic Ca2 + transients is about 0.06 Hz in hippocampal neuronal cell cultures (Fig. 4A) [4, 30]. Using the same experimental conditions (10 DIV, see Material and Methods), we observed that alkaloids alone were able to increase the transients over the control conditions (164±4% and 140±3% for Lup and 17– ox, respectively). While that the SO-Aβ (0.5μM, 24 h) produced a drastic decay (white bars) in the amplitude (Fig. 4B) and frequency (Fig. 4C) of Ca2 + transients, leading in example only the 63±2% of the control frequency. These data suggest that there are detrimental effects on the neuronal network synaptic activity (Fig. 4A, second trace). On the other hand, the presence of 17– ox (0.03μM) and Lup (0.03μM) prevented the decay and restored the amplitude and frequency almost to the control values (Fig. 4B and 4C, respectively). These data indicate that the presence of these alkaloids can help to maintain and promote the neuronal communication network in hippocampal neurons.

Changes in cytosolic Ca2 + transients induced by chronic treatment with SO-Aβ in the presence of 17– ox and Lup in hippocampal neurons. A) Original recordings of spontaneous Ca2 + signals measured with Fluo4– AM in hippocampal neurons in control conditions, treated for 24 h with SO-Aβ (0.5μM) alone, co-incubated with 17– ox (0.03μM) and Lup (0.03μM). B) Quantification of amplitude (F/Fo represented as % of control) for conditions in A. C) Quantification of frequency (events/min) for conditions in A represented as percentage of control (N = 4; n = 40). *p < 0.05 and ***p < 0.001 versus control; # # #p < 0.001 versus Aβ.
17– ox and Lup neuroprotective properties are mediated by nicotinic α7 receptors
The effect of quinolizidinic alkaloid derivates on nicotinic receptors have been widely studied [24, 31–34]. Therefore, in order to determine if the effect of these two alkaloids involve nicotinic receptors, we used HEK cells as a negative control since no nicotinic receptors have been reported in this cell line [35]. We incubated HEK cells with SO-Aβ (0.5μM, 24 h) and observed a decrease in cell viability by 28±3% compared to control conditions, while 17– ox and Lup (0.03 and 0.3μM, respectively) had no effect on cell viability (LUP alone at 0.03μM: 98±4%; at 0.3μM: 99±6%; 17– ox at 0.03μM: 99±7%; at 0.3μM: 98±4%, Fig. 5). Hence, these results reinforce the idea that neuroprotection and maintenance of neuronal network activity could be mediated by nicotinic receptors.

17– ox and Lup did not prevent the toxicity of SO-Aβ in HEK293 cells. Effects of 17– ox and Lup (0.03–0.3μM) on cell viability using MTT techniques to evaluate the toxicity induced by SO-Aβ (0.5μM, 24 h) in a cell model that does not express nAChR. FCCP (10μM) was used as a control for toxicity. The values represent the percentage of control without treatment (n = 3; N = 18). *p < 0.05, **p < 0.01, and ***p < 0.001 versus control.
In parallel, PC–12 cells were treated under similar conditions as shown in Fig. 5, but with the additional presence of a specific α7 nicotinic receptor blocker, α-bungarotoxin (α-BTX, 3μM, Fig. 6). Under these experimental conditions, 17– ox and Lup (0.03 and 0.3μM) do not alter viability (LUP at 0.03μM: 98±4% : at 0.3μM: 99±6%; 17– ox at 0.03μM: 99±7%; at 0.3μM: 98±4%) and additionally, were unable to induce cell protection as observed in Fig. 3, suggesting that the presence of nicotinic α7 receptors is important to exert their neuroprotective actions. Furthermore, it is possible that the alkaloids can modulate the activity of this receptor and the signaling pathway associated with α7. Nevertheless, we cannot discard the modulation of other subtypes of the nicotinic receptor such as α4b2, or α3b4. Note that FCCP was not in the presence of bungarotoxin.
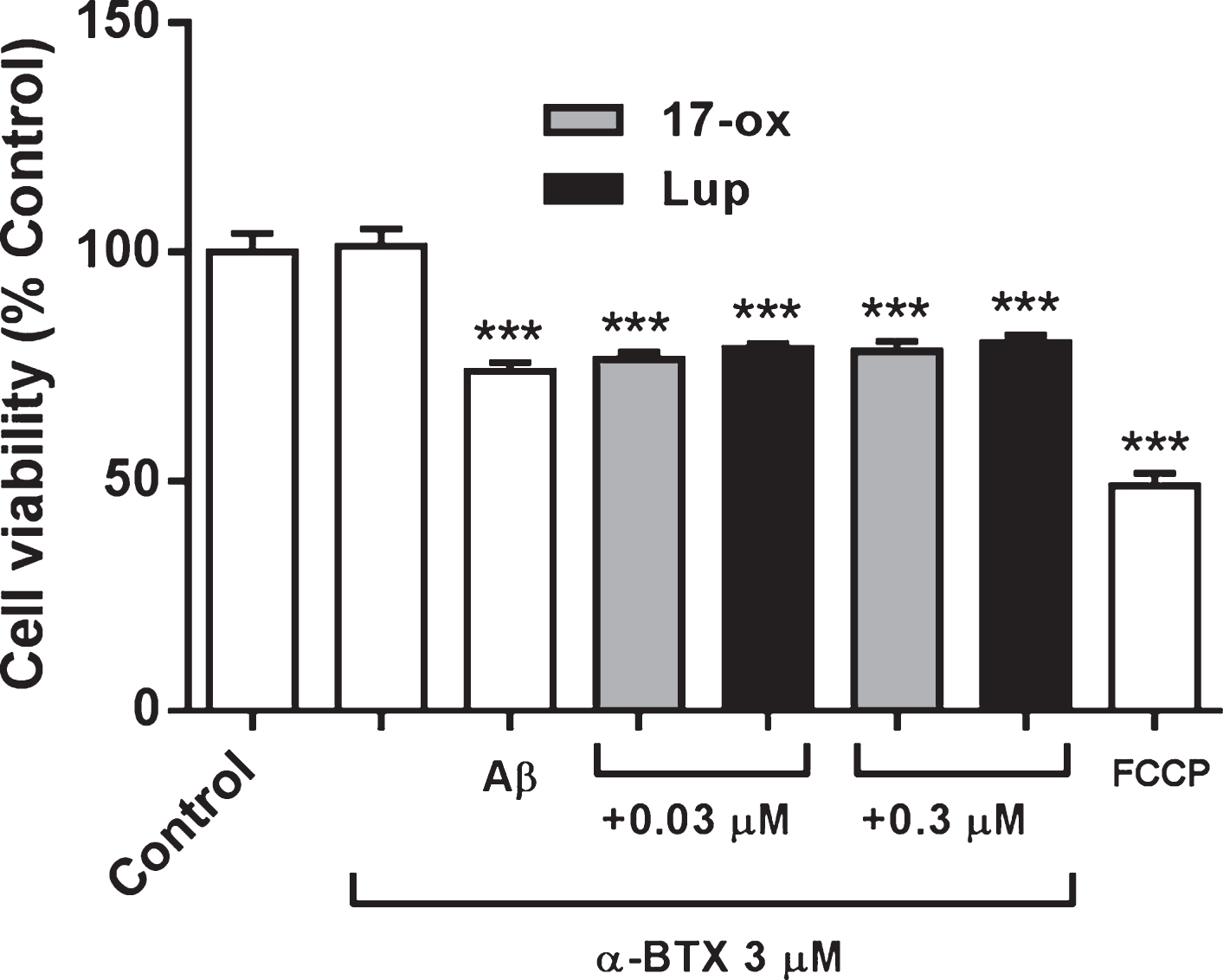
Effects of α-bungarotoxin, a selective α7 nAChR blocker, on the effects of 17– ox and Lup on SO-Aβ toxicity in PC–12 cells. PC–12 cells were treated with similar condition for Fig. 4, plus the co-incubation with α– BTX (3μM). FCCP (10μM) was used as a control for toxicity. The values represent the percentage of control without treatment (n = 3; N = 18). ***p < 0.001 versus control.
Commonly, the neuroprotective properties associated with α7 nAChR have been attributed to the activation of survivor pathways, mainly to phosphorylation of Akt (p-Akt) and an increase in the p-Akt/Akt ratio [15, 37], which is classically associated to antiapoptotic effects [38–40]. To evaluate if our alkaloids exert modulation of this pathway, we incubated PC12 cells with SO-Aβ (0.5μM) alone or co-incubated with 17– ox and Lup (0.03μM) for 1 h. Subsequently, western blot was done to detect the levels of p-Akt and Akt (Fig. 7A, upper panel), and the data showed that 17– ox had a non significant increment in p-Akt/Akt (27±15%), while Lup induced an increase of 58±12% at 1 h (Fig. 7A, lower panel). After 3 h of treatments, on the other hand, both alkaloids were able to increase the p-Akt/Akt ratio (35±7% and 52±4%, respectively; Fig. 7B).
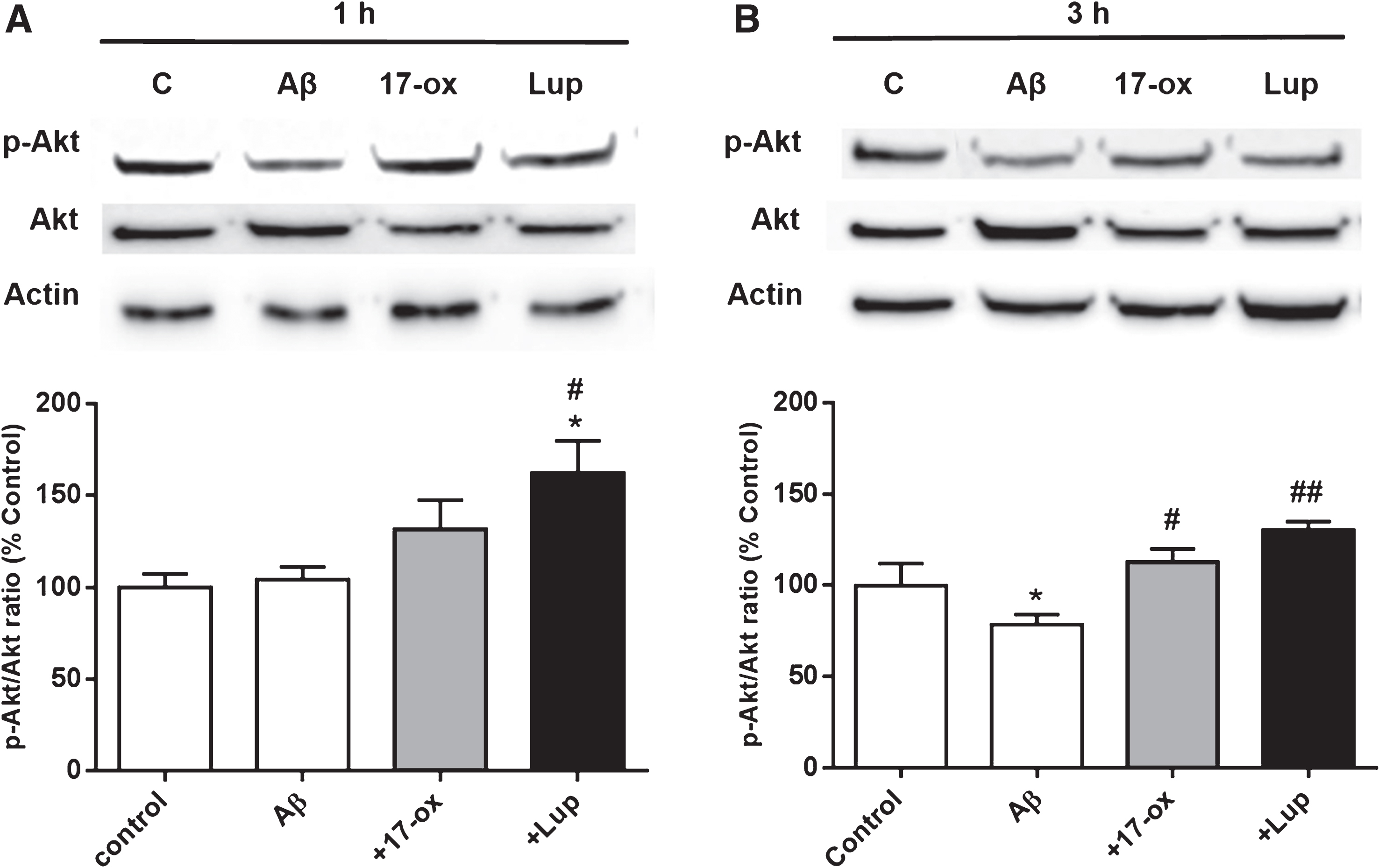
Activation of the Akt pathway by 17– ox- and Lup in PC–12 cells treated with SO-Aβ. A) Western blot of p-Akt and Akt levels using specific antibodies (upper panel) in PC–12 cells treated for 1 h with SO-Aβ (0.5μM) alone, or co-incubated with 17– ox (0.03μM) or Lup (0.03μM). The quantifications of band intensities are represented as percentage of control conditions (lower graph). B) Western blot using similar conditions shown in A, but after 3 h of treatment. The graphs represent the ratio p-Akt/Akt (n = 3). *p < 0.05 versus control;
Taken together, it appears that an early activation of the Akt pathway is important to induce and maintain neuronal functioning against SO-Aβ toxicity. All the more interesting is that those effects were observed at very low alkaloid concentrations, suggesting a high potency (0.03μM), and also the involvement of nicotinic α7 activation. In order to compare the actions of our isolated alkaloids with Nicotine on cell survival and reinforce the idea that α7 is involved in the effect of these alkaloids, we co-incubated PC–12 cells with SO-Aβ (0.5μM, 24 h) alone and co-incubated with 17– ox (0.03μM), Lup (0.03μM), or Nicotine (10μM). Subsequently, cell viability was measured using the MTT assay. According with previous results, 17– ox induced a neuroprotection showing recovery of the SO-Aβ damage to near control values (97±1%), while co-incubation of 17– ox plus Nicotine potentiated this effect with a viability value of 116±2% (Fig. 8A). Nicotine was also able to potentiate (127±3%) the effects of Lup on cell viability (105±2%) when they were co-incubated together (Fig. 8B). Figure 8C confirms that the observed effects were mediated by Nicotinic receptors since the presence of α-BTX blocked all of the protective effects, indicating that the neuroprotective effects of 17– ox and Lup are modulated by the nicotinic receptor, possibly by α7 (Fig. 8C).
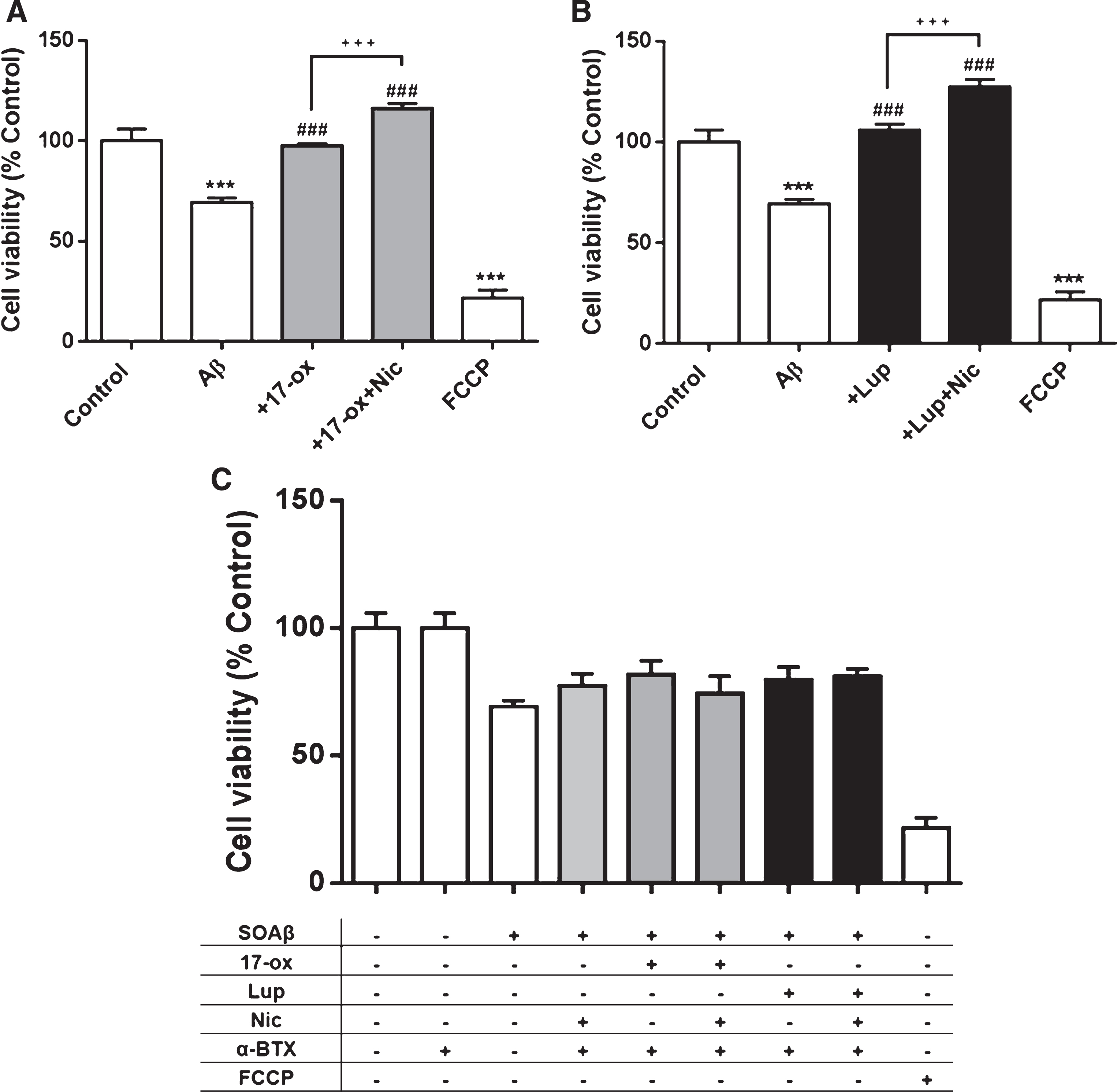
Comparison of neuroprotector effects of alkaloids that modulate α7 nAChR (Nicotine, 17– ox, and Lup) in PC–12 cells treated with SO-Aβ. A) Cell viability of PC–12 cells treated with SO-βA (0.5μM, 24 h) and co-incubated with 17– ox (0.03μM) alone and in the presence of Nicotine (10μM). B) Cell viability of PC–12 cells treated with SO-Aβ (0.5μM, 24 h) and co-incubated with Lup (0.03μM) alone and in the presence of Nicotine (10μM) (n = 3; N = 9). C) α-BTX (3μM, 24 h) blocks the neuroprotector effect of alkaloids that modulate α7 nAChR (Nicotine, 17– ox, and Lup) in PC–12 cells treated with SO-Aβ. PC–12 cells treated under similar conditions as A and B, but in the presence of α-BTX (3μM, 24 h). FCCP (10μM) was used as a control for toxicity (n = 3; N = 12). ***p < 0.001 versus control;
DISCUSSION
The relevance of this work is related with the mechanism of action of these two alkaloids purified from Cytisus scoparius that have shown beneficial effects on cell viability, which has not been previously studied, and additionally these protective effects are mediated by nicotinic receptors. Both compounds at submicromolar concentrations prevented amyloid toxicity induced by SO-Aβ. Additionally, 17– ox and Lup activate the AKT cascade, an important survival pathway. More interesting, these alkaloids contributed to maintenance of synaptic activity and neuronal network connectivity in hippocampal neurons. This study is the first to provide data showing a potential neuroprotective effect of these two alkaloids on neurons exposed to SO-Aβ.
The main evidence related to pharmacological effects of compounds obtained from Cytisus are linked to cytisine from seeds that have been used to treat smoking [41], antidepressive alterations [42], and neural protection against glutamatergic exitotoxicity [23]. However, no other molecules or alkaloids obtained from this family (Fabaceae) have been reported to be neuroprotective or have biological properties. Although Lupanine has been shown to have some physiological effects on glucose homeostasis and insulin release [43, 44], other oxo-substituted sparteins, like 17– oxospartein, do not have reported biological activity. Therefore, this study provides the first evidence of a pharmacological application for these alkaloids and their therapeutic use; however, remain pending to characterize or elucidate the precise concentration-response dependence for these alkaloids in future works.
In our hands, these quinolizidinic alkaloids had a high potency compared to Nicotine that had an EC50 in the micromolar range for the different Nicotinic receptor isoforms [45]; in particular for α7 nAChR the EC50 was close to 13 micromolar [45]. This is especially important since Nicotine has shown strong neuroprotective properties [46], however, the addictive properties has been the reason for its disuse. It is interesting to think that molecules with a high potency used early in AD development could be more effective than actual drugs [47]. Hence, 17– ox and Lup are very interesting molecules to develop new pharmacological entities with application for AD. More studies will be needed to further characterize them pharmacologically, with special attention on the use of mouse AD models. Our results indicate potential actions on α7 nAChR since co-incubation of both alkaloids with Nicotine did not show different effects to those with Nicotine alone, suggesting that all these molecules use the same target, this are reinforced with the observation related to the potentiation of nicotine effects with alkaloids, that shown to be more potent that alkaloids alone (Fig. 8). Moreover, the use of α-BTX, a specific α7 nAChR blocker, completely abolished the effects of all nicotinic agonists. Although α-BTX selectivity has been demonstrated with immunoprecipitation experiments using monoclonal antibodies [48], the same data show that the toxin did not block and was unable to bind to α4β2. In general terms, α4β2 and α7 have been associated to the modulation of synaptic neurotransmission and neuroprotective properties mediated by Nicotine [49, 50]. Therefore, the fact that α-BTX blocked all the effects of 17– ox and Lup leads us to think that their neuroprotective effects are mediated mainly by α7 nAChR.
On the other hand, the activation of α7 nAChR have been shown to activate Akt thereby regulating cellular apoptosis [18]. In our hands, the phosphorylation of Akt was significant and fast (1 and 3 h) according to other studies [51] and correlated with the chronic effects observed at 24 h [24]. It has been postulated that Nicotine, through nAChR, could alter cellular cycle promoting hyperplasia in lung cancer [52, 53] by the PI3K/AKT pathway [51]; however, with the experimental approach used in this study, it was not possible to discard these effects in cell lines and require more studies, but is important to comment that the activation of cholinergic system could be involved in a beneficial effects by itself, as has been demonstrates on neuronal and non-neuronal tissues, in this way two alkaloids shown to promote a more healthy conditions (Figs. 3 and 4) in the cells we it were apply alone, reinforcing this idea and the observation related with the neuroprotective effects and the preservation of synaptic connectivity is what fails in several AD models [24, 54].
In this context, the evidence related with the preservation of Ca2 + signals as an indicator of synaptic activity is relevant, since our group has described that Aβ peptide induces an acute excitotoxicity by massive release of neurotransmitters [5] that with time leads to vesicle depletion and synaptic failure [7, 26]; all these events mediated by Ca2 + dyshomeostasis [54, 55]. In our hands, chronic treatment of neurons with 17– ox and Lup prevented these Ca2 + alterations suggesting that modulation of nAChR could possibly regulate synaptic function and prevent neuronal death. Additionally, these observed effects could be useful to prevent the glutamate toxicity in the synaptic spaces, and then the action mechanism of these molecules could be useful against the glutamate excitotoxicity and its Ca2 + overload, but require being study deeply.
In all the experiments, Lup had more potency than 17– ox, suggesting that the position of oxygen could perhaps be important for biological activity, as has been demonstrated by spartein that affects mainly mAChR, while Lup that is different only in the oxygen in carbon 2, has shown to be active on nAChR [56].
In summary, these alkaloids isolated from Cytisus scoparius modulate the nAChR, activate the Akt pathway, and preserve synaptic activity in hippocampal neurons, and represent important types of molecules that open a new chemical space to develop new pharmacological tools to treat Aβ toxicity and AD.
Footnotes
ACKNOWLEDGMENTS
This work was funded by FONDECYT 1161078 (JF). JGG is MSc fellow of Postgraduate program of U. de Concepcion. TSG and PAG are PhD students of CONICYT (21160392 and 201161295, respectively). We thank Laurie Aguayo for edition of this manuscript, Ixia Cid for technical assistance, and CMA-BIOBIO for their technical support.