
Abstract
There is a great need for diagnostic biomarkers of impending dementia. Metabolite markers in blood have been investigated in several studies, but inconclusive findings encourage further investigation, particularly in the pre-diagnostic phase. In the present study, the serum metabolomes of 110 dementia or pre-diagnostic dementia individuals and 201 healthy individuals matched for age, gender, and education were analyzed by nuclear magnetic resonance spectroscopy in combination with multivariate data analysis. 58 metabolites were quantified in each of the 311 samples. Individuals with dementia were discriminated from controls using a panel of seven metabolites, while the pre-diagnostic dementia subjects were distinguished from controls using a separate set of seven metabolites, where threonine was a common significant metabolite in both panels. Metabolite and pathway alterations specific for dementia and pre-diagnostic dementia were identified, in particular a disturbed threonine catabolism at the pre-diagnostic stage that extends to several threonine-linked pathways at the dementia stage.
INTRODUCTION
Alzheimer’s disease (AD) and vascular dementia (VaD) are the two most common forms of dementia, and although extensively studied, the disease mechanisms behind these disorders remain elusive. The assembly of amyloid-β (Aβ) peptide plaques and/or neurofibrillary tangles of the tau-protein within the brain is associated with AD [1], while a vascular-induced pathophysiology is implicated in VaD [2]. For AD, the Aβ1–42 peptide, the total tau protein, and the phosphorylated tau protein are three generally accepted cerebrospinal fluid biomarkers with high diagnostic accuracy [3 –5]. There is still a need for additional biomarkers of AD and dementia, especially biomarkers predictive of disease prior to any clinical symptoms, preferably obtained from an easily accessible peripheral body fluid that requires less invasive sample collection (e.g., [6, 7]).
Blood-based metabolite markers would be an attractive diagnostic option for dementia, and numerous metabolomics studies of blood [8 –14] have indeed identified putative markers of dementia and AD. Several extensive blood-based studies have been published [15 –20], detecting lipid and metabolite markers of dementia (e.g., ceramides, phosphatidylcholines, sphingomyelins, branched-chain amino acids (BCAA), and glutamine), as well as differences in the lipoprotein lipid profiles between dementia and control groups. Although potential biomarkers were identified, the limited overlap between studies requires further validation. At present there is no clear and indisputable blood metabolite marker or metabolite panel identified for dementia or pre-dementia. The fairly modest disease-specific changes of the metabolome detected, and the limited overlap and even contradictions between studies, highlight the importance of careful experimental designs, analysis methods, and rigorous statistical/bioinformatic analysis [17]. Many of the studies referred to above were carried out using mass spectrometry (MS)-based methods, which has the advantage of providing excellent sensitivity. An alternative, less sensitive but very robust quantitative approach is NMR-based metabolomics/metabonomics [21 –23], a method that can potentially detect subtle differences between patients and controls. In our previous studies of dementia, we successfully applied MS for the analysis of saliva and serum [24] and NMR for the analysis of saliva [25], and NMR was used in the present work for the analysis of serum samples using the same study design and an expanded set of samples.
In the present study, the serum metabolomes of 110 individuals with dementia or subjects who within the following five years after sample collection received a diagnosis of dementia, as well as 201 age, gender, and education-matched healthy controls, were quantitatively analyzed by NMR spectroscopy. We compared the metabolite signatures of these groups by using multivariate statistical analysis and identified putative metabolic markers for both the dementia and pre-diagnostic dementia groups, as well as a limited set of affected metabolic pathways.
MATERIALS AND METHODS
Participants
Serum samples were obtained from the Betula bio-samples collection, which is part of the prospective longitudinal Betula study on aging, memory, and dementia [26, 27]. Since its start in 1988, the Betula study has monitored the health and cognitive development in 4,500 participants through comprehensive health and neuropsychological assessments, as well as collected biological samples from the participants. Every fifth year a new assessment has been performed, and in total six test waves (T1–T6) have been carried out over a period of 25 years. The AD and VaD subjects were diagnosed according to the DSM-IV criteria [28] as previously described [29, 30]. The serum samples analyzed in the current study were obtained at test wave T4 (between September 2003 and April 2005), at which time point also saliva was collected, facilitating parallel metabolomics studies using saliva and serum from the same patients [24, 25]. In total 1,142 individuals had biofluids available at T4, where 53 were diagnosed with either AD or VaD, and 57 showed no signs of dementia at T4 but developed AD or VaD until T5; these individuals were all included in the present study. Although analysis of serum samples from other time-points would have improved the power of our present study the original design that included samples from T4 only was retained. Control subjects were selected from T4 and if they met the following criteria: 1) no dementia at the T5 diagnostic evaluation, 2) available results from health and cognitive assessments at T4, 3) available biofluids at T4, and 4) available DNA or buffy coat from any test occasion. To each dementia case the goal was to identify and select two controls, matched for age, gender, and education, as well as vital status (alive/deceased). These subjects were subsequently divided into two substudies based on the onset of the dementia disorder. Substudy 1 (SS1) included 53 dementia patients (40 AD, 13 VaD) and 91 control subjects from T4. Substudy 2 (SS2) aimed at identifying pre-diagnostic metabolic markers of dementia at T4 and included samples collected from 57 healthy subjects (at T4) who were diagnosed with dementia (28 AD, 29 VaD) five years later (at T5) and from 110 control subjects who were healthy at both T4 and T5. The difference in the AD:VaD participant ratio for the prevalent cases in SS1 (ratio ∼3 : 1) compared to the incident cases in SS2 (∼1 : 1) may be a reflection of the two-fold lower participation frequency among prevalent VaD patients (18%) compared to prevalent AD patients (35%), which in part should be dependent on the higher mortality rate among VaD patients (57% versus 36% for AD patients). Table 1 summarizes the group and participant characteristics, including clinical information and the primary medication.
Group characteristics of substudies 1 and 2, showing the number of included subjects, disease status, age and gender distribution, education, MMSE, BMI, other disorders, and the number of prescribed drugs
Age, education, MMSE, and BMI are presented as mean±SD; MMSE, Mini-Mental State Examination; BMI, body mass index; na, not available; Hypertensive, diabetic or smoker within the last five years. A list of drugs for each group of medication follows: 1Adalat, Atacand, Atenolol, Captopril, Cardizem, Cozaar, Diovan, Emconcor, Felodipin, Inderal, Isoptin, Logimax, Norvasc, Plendil, Propranolol, Renitec, Seloken, Triatec; 2Esidrex, Furix, Lasix, Linatil, Salures, Spironolakton; 3Plavix, Trombyl; 4Behepan, Betolvex, Betolvidon, Folacin, Pyridoxin, TrioBe; 5Lipitor, Lopid, Questran, Simvastatin, Zocord; 6Daonil, Glipizid, Glucophage, Insulatard, Insulin, Metformin, Mixtard; 7Aricept, Ebixa, Exelon, Reminyl.
The present NMR study is a follow-up of our previous MS-based investigation on serum and saliva [24] and NMR-based study on saliva [25], both using the same study design and largely identical sets of samples. The Betula study has been carried out in accordance with the Declaration of Helsinki (WMA, 1964) with ethical approval from the Regional Ethical Review Board in Umeå, Sweden. Written informed consent was obtained from all subjects.
Sample collection and preparation
The sample collection took place within one week of the health and neuropsychological assessments. Serum samples from non-fasting subjects were obtained from venous blood and stored at –80°C until NMR analysis. NMR samples were prepared as described in the Supplementary material, and the resulting 200 μL samples were transferred to 3 mm (600 MHz) NMR tubes (Bruker Biospin) for subsequent NMR analysis.
NMR spectroscopy, data processing, and spectral analysis
NMR experiments were carried out at 25°C on a Bruker 600 MHz AVANCE III spectrometer equipped with a 5 mm triple-resonance z-gradient cryo-probe (CPTCI 1H-31P/13C/2D Z-GRD) and a SampleXpress sample changer. TOPSPIN, version 3.2, was used for spectrometer control and data processing (Bruker Biospin). NMR data acquisition and processing were carried out as described in the Supplementary Material.
Metabolite identification and quantification was carried out with the Chenomx NMR Suite software package, version 8.01 (Chenomx Inc., Edmonton, Canada). This program uses a line-fitting procedure in which NMR resonances of a recorded spectrum are mathematically modeled using a database containing reference spectra of 332 metabolites. Metabolite identities were confirmed by comparing our results with previously reported assignments, in particular those from the human metabolome data bank [31 –33].
Statistical analysis
Multivariate statistical data analysis and receiver operating characteristic (ROC) analysis was carried out using SIMCA 13–15 (Umetrics, Umeå, Sweden). In the simultaneous statistical analysis of multiple variables, datasets were mean-centered and unit variance scaled and then subjected to principal component analysis (PCA) to obtain an overview of the data and to orthogonal projections to latent structures discriminant analysis (OPLS-DA) [34, 35] to identify features that separate the groups and, in this context, to identify metabolic markers of dementia. The OPLS-DA models for SS1 and SS2 were all calculated with 1 + 1 + 0 components to avoid over-fitting of data and cross-validated by leaving matched groups out. R2X and R2Y are the fractions of the explained variation for the X and Y component, respectively. Q2 indicates the goodness of a prediction and describes the fraction of the total variation predicted by a component, as estimated by cross-validation. Group separations were evaluated through CV-ANOVA [36] (expressed as p-values for the models), and important metabolites were identified through the first weight vector (w*[1]). The cut-off values of w*[1] were determined empirically by iterative model building to obtain the most significant model. Univariate statistical analysis was carried out by Student’s t-tests, and pathway analysis was carried out with the MetaboAnalyst [37] and HumanCyc [38] web servers.
RESULTS
NMR spectra of 311 filtered samples from the subjects in the dementia substudy (SS1) and in the substudy of pre-diagnostic dementia (SS2) were recorded, and 58 metabolites were identified and quantified in each sample using the Chenomx software. Group and clinical parameters of the dementia, pre-diagnostic dementia, and matched control groups of SS1 and SS2 are summarized in (Table 1). A complete list of the quantified metabolites, including their average concentrations and standard deviations, as well as fold changes, statistical significance, and weight values for each group comparison, is provided in Supplementary Table 1.
Although the multivariate statistical data analysis by PCA revealed no significant outliers or trends, initial OPLS-DA analysis of the data from the pre-diagnostic dementia study (SS2) identified one clear outlier among the samples from the VaD patients. This sample turned out to have an approximately sixty times higher concentration of dimethyl sulfone than the average of the other samples, and this patient and their matched controls were therefore excluded from the statistical analysis.
In SS1, the initial OPLS-DA modeling (using all metabolites) identified the following seven metabolites as particularly important (|w*[1]| >0.2) for the separation between the dementia and control groups: threonine, creatine, alanine, acetate, aspartate, acetoacetate, and N,N-dimethylglycine (Fig. 1A). The final OPLS-DA model was calculated using these metabolites, which resulted in a statistically significant model (R2X = 0.40, R2Y = 0.13, Q2 = 0.067, p = 0.047), see the cross-validated score plot in (Fig. 1B). However, the ROC analysis using these seven metabolites showed a fairly poor discriminatory ability with an AUC of 0.61 (Fig. 1C). The univariate statistical analysis did not recognize the altered metabolite concentrations as significant. To confirm the relevance of the selected metabolites with an alternative method, the more elaborate OPLS effect projection (OPLS-EP) analysis was performed [39]. This method was specially designed for pairwise comparisons on a multivariate basis and for detecting subtle disease-associated variations that may otherwise be hidden among variations of other sources such as age, gender and sample storage time. The OPLS-EP method may provide some advantages over OPLS-DA, but it is less frequently used and therefore the OPLS-EP results are presented in the Supplement. The discriminatory metabolites identified in the OPLS-DA of SS1 were all present in the OPLS-EP analysis (Supplementary Figure 1). The dementia group did however include 15 patients with anti-AD treatment (Table 1), and to ensure that their medication did not notably affect the outcome of the OPLS-DA, an additional modeling was carried out where these patients were removed, resulting in a slightly improved statistically significant model (R2X = 0.42, R2Y = 0.16, Q2 = 0.078, p = 0.038).
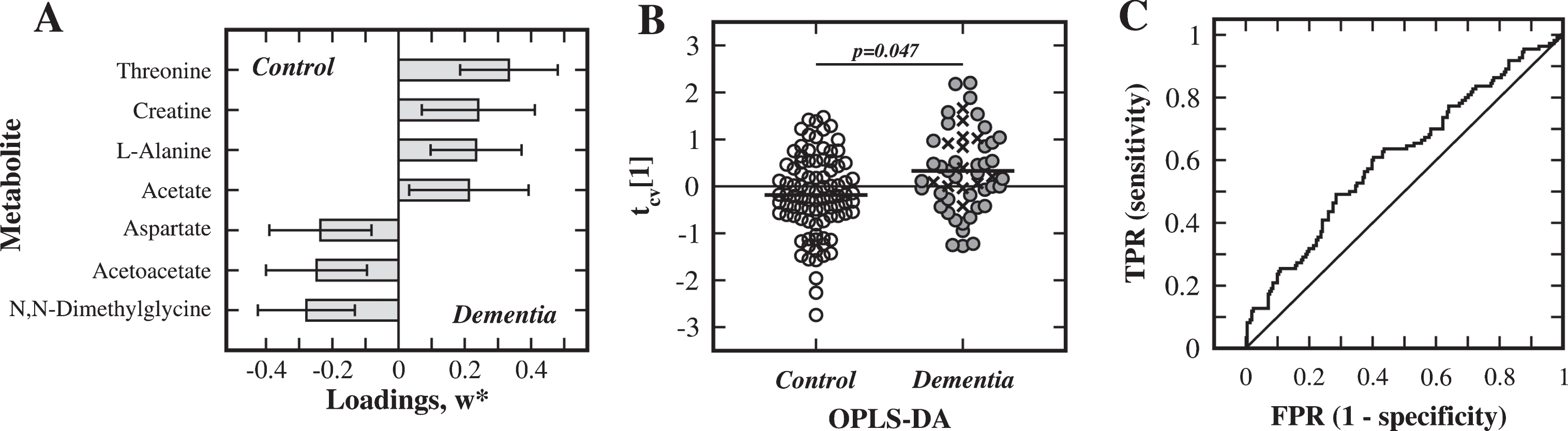
Analysis of substudy 1. Multivariate statistical comparison of serum from dementia patients and matched control subjects in SS1. A) Metabolites, with weight values |w*| >0.2, important for separation between the dementia and control groups. B) Cross-validated OPLS-DA score plot of the first component, tcv[1] (AD: filled circles; VaD: crosses; control subjects: open circles). C) ROC analysis for discrimination between groups using the discriminatory metabolites (TPR: true positive rate, FPR: false positive rate).
For the comparison of pre-diagnostic dementia patients with controls (SS2), the initial OPLS-DA analysis identified the following panel of seven metabolites as particularly important (|w*[1]| >0.2) for discriminating between the groups: threonine, trimethylamine, isobutyrate, propylene glycol, glutamine, 2-hydroxybutyrate, and tyrosine (Fig. 2A). Using this set of metabolites, a statistically significant OPLS-DA model (R2X = 0.53, R2Y = 0.12, Q2 = 0.068, p = 0.023) was obtained, see the cross-validated score plot in (Fig. 2B). The accompanying ROC analysis resulted in an AUC of 0.71, which supports the use of these seven metabolites for distinguishing between the groups (Fig. 2C). In the univariate analysis, only the concentration of threonine was significantly changed (p = 0.0006) (Fig. 3). In this case an OPLS-EP analysis was also carried out, confirming the relevance of five of the identified discriminatory metabolites (Supplementary Figure 2).
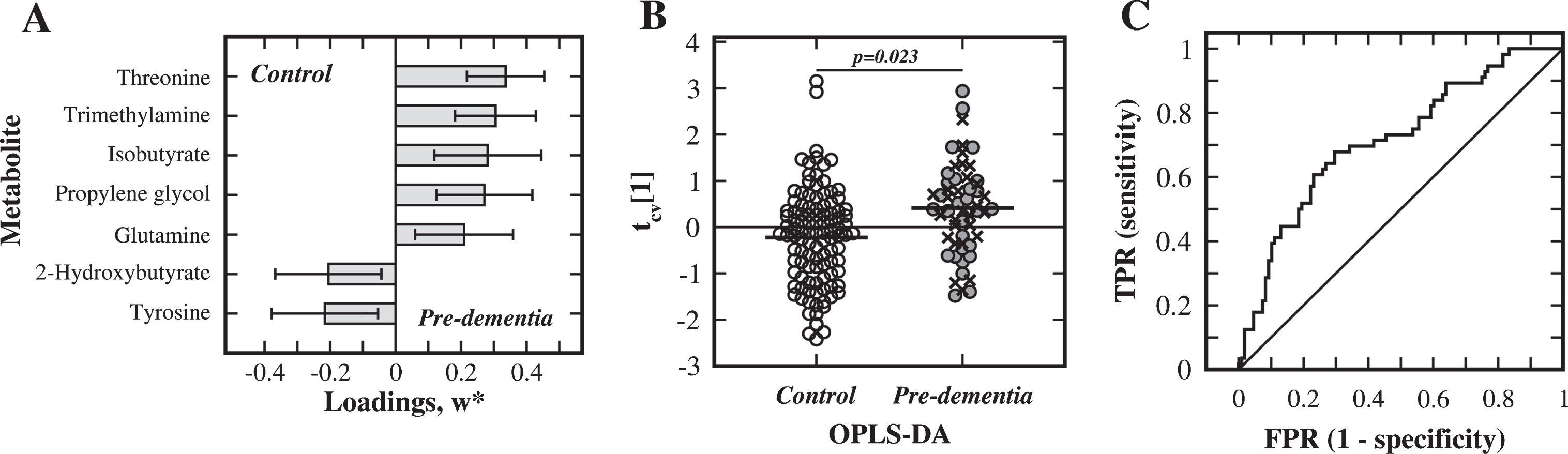
Analysis of substudy 2. Multivariate statistical comparison of serum from the pre-diagnostic dementia (Pre-dementia) patients and matched control subjects in SS2. A) Metabolites, with weight values |w*| >0.2, important for separation between the pre-diagnostic dementia and control groups. B) Cross-validated OPLS-DA score plot of the first component, tcv[1] (Pre-AD: filled circles; Pre-VaD: crosses; control subjects: open circles). C) ROC analysis for the discrimination between groups, using the discriminatory metabolites (TPR: true positive rate, FPR: false positive rate).

Threonine concentration in substudy 1 and 2. Bar chart showing the concentrations (mean and standard deviation) of threonine in SS1 and SS2, where shaded bars indicate dementia or pre-diagnostic dementia patients, and open bars represent matched control subjects. The statistical significance of the difference in SS2 is indicated.
The fold changes and |w*|-values for the selected metabolites of the dementia substudy (SS1) and the pre-diagnostic dementia substudy (SS2) are listed in (Table 2). Additional multivariate modeling of SS1 using the seven metabolites of SS2, modeling of SS2 using the seven metabolites of SS1, and modeling of either SS1 or SS2 using all thirteen discriminatory metabolites failed to give any significant models. The AD and VaD patient groups (identified with different symbols in Figs. 1B and 2B) were initially treated separately, but since the two groups showed no statistically significant separation and statistically behaved nearly identical in comparisons with controls (data not shown), they were consequently treated as one group. Furthermore, no statistically significant differences due to gender or age at onset or education level were detected, which was expected due to the design of the study (data not shown).
Serum metabolites important for the separation of dementia subjects from matched controls (substudy 1, SS1) and for the separation of pre-diagnostic dementia subjects from matched controls (substudy 2, SS2) based on the multivariate statistical OPLS-DA models. Increased and decreased concentrations of the metabolites are indicated with an upward or downward pointing arrow, respectively, together with their corresponding fold change and absolute weight value |w *| from the OPLS-DA model. Bold arrows indicate metabolites with a p-value below 0.05 in the univariate analysis. A selection of affected pathways and/or associated physiological functions or origins for each metabolite is listed
BCAA, branched chain amino acid; GABA, γ-aminobutyric acid; CVD, cardiovascular disease.
DISCUSSION
The present NMR-based study searched for metabolic markers of dementia and consisted of two carefully designed substudies. In the first substudy (SS1), samples from dementia patients were compared with matched healthy controls, and in the second substudy (SS2) samples from subjects who developed dementia within 5 years after sampling were compared with matched healthy controls (Table 1).
Discriminatory metabolites
In SS1 the multivariate statistical analysis of the NMR data separated the dementia and control groups in a fairly weak but significant model using a panel of seven metabolites (Fig. 1). In SS2 a different panel of seven metabolites was identified, resulting in a statistically significant model discriminating between the pre-diagnostic dementia and control groups (Fig. 2). The thirteen discriminatory metabolites (seven metabolites from SS1 and seven from SS2, with threonine in common) are listed in (Table 2), together with a brief overview of their roles in various metabolic pathways. The results in both SS1 and SS2 are in line with recent metabolomics studies of serum and plasma, which also display a modest sensitivity and congruence. The different compositions of the two panels of discriminatory metabolites are interesting and might to some extent be explained by the hypothetical biomarker model of AD in which the magnitude of biomarkers should change with disease stage or time in a sigmoidal fashion [40]. At an early pre-clinical/pre-diagnostic stage, markers for dementia and AD might therefore be below the detection limit, but it is possible that the early neurodegenerative process might affect other metabolic pathways leading to detectable differences for different markers. This speculative explanation of an indirect effect is potentially interesting and needs to be addressed in future studies. The group separation in SS2 was somewhat better than in SS1, which might reflect the larger variation and heterogeneity typically observed in later stage dementia and in older subjects. The fairly modest AUC:s observed in SS1 and SS2 (Figs. 1C and 2C) may reflect limited pathophysiological differences between the metabolomes of these groups, but also a heterogeneity within the groups caused by for example co-morbidities at old age, nutritional intake, gut microbiota and medication. Furthermore, no separation of the AD and VaD patient groups was observed within the dementia and pre-diagnostic dementia groups, indicating a similar metabolite profile of these dementia disorders, which is in agreement with our previous studies [24, 25] and likely a manifestation of the overlapping clinical-pathological similarities [41].
In the interpretation of our results a few sources of errors and limitations need to be mentioned. Since the samples were collected between September 2003 and April 2005, the results may be affected by the storage time as well as a seasonal effect. There may also be dietary effects on the analysis results since serum was obtained from non-fasting individuals; the type of nutrition is further known to be an important factor for modifying the risk for dementia [42]. Medication and other clinical parameters (Table 1), as well as lifestyle changes induced by cognitive decline may further influence the outcome. In order to validate the identified discriminatory metabolites these findings need to be replicated in an independent material.
In the discussion below, the biological significance of the identified metabolites will be assessed, and links to pathways known to be affected in dementia will be identified. It is however important to perform a careful interpretation because serum is a peripheral body fluid not directly in contact with the neurodegenerative processes of the brain. Furthermore, it is at this time not possible to determine if any of these metabolic changes are causing dementia or if they are an effect of the syndrome.
Identifying a putative metabolic pathway towards dementia
A central outcome of this study is the identification of a putative pathway for the metabolic changes during the dementia disease process. An initial disturbance in threonine metabolism was observed in pre-diagnostic dementia patients, which subsequently evolved into a broader effect on threonine metabolism in dementia patients (Table 2 and Fig. 4). Most notable was the elevated concentrations of the essential amino acid threonine found in the pre-diagnostic dementia (SS2, fc = 1.11) and the dementia (SS1, fc = 1.10) patients when compared to control subjects (Fig 3). This observation was particularly clear in SS2, where the concentration difference was significantly (p = 0.0006) higher in pre-diagnostic dementia patients. An elevated level of threonine was previously identified in AD brain extracts [43], but in a recent serum study among AD patients no significant change was observed [44].

Schematic illustration of the metabolic pathways putatively affected in substudy 1. The pathways between discriminatory metabolites are indicated with grey arrows and intermediate metabolites with circles. The up- and downward pointing wide arrows indicate increased and decreased metabolite concentrations, respectively, in dementia patients (grey), in pre-diagnostic dementia patients (hollow), or in both (dark grey).
Threonine is primarily catabolized by the enzyme threonine dehydratase into 2-ketobutyric acid, which in turn can be converted into 2-hydroxybutyric acid through a reaction catalyzed by lactate dehydrogenase or alpha-hydroxybutyrate dehydrogenase. In the pre-diagnostic dementia substudy (SS2) a decreased concentration of 2-hydroxybutyic acid was in fact observed (fc = 0.94; Fig. 4), which is in agreement with a recent study of plasma from AD patients and control subjects [16]. 2-ketobutyric acid can also be converted into propionyl-CoA, an important metabolite in BCAA metabolism. Although we did not observe any significant changes in the concentration levels of BCAAs (valine, leucine, and isoleucine), there are several reports implicating BCAA metabolism in AD patients [16 , 19]. Both of these routes for catabolism of threonine can consequently be linked to disturbances in metabolic pathways in pre-dementia and dementia patients.
For the dementia patients, a more extensive effect on threonine metabolism was detected, where the significantly altered metabolites threonine (fc = 1.10), aspartate (fc = 0.91), creatine (fc = 1.19), and N,N-dimethylglycine (fc = 0.86) are all part of the glycine, serine, and threonine metabolic pathway (Fig. 4). The same pathway was implicated in a recent metabolomics study of AD brain extracts, where similar alterations of the threonine and aspartate levels were observed [43]. This does not, however, rule out that threonine can be connected to alterations in other pathways, such as BCAA metabolism. The reduced level of N,N-dimethylglycine observed here is in agreement with a previous study where patients with AD or mild cognitive impairment were compared to healthy controls [11].
Aspartate is furthermore directly linked to alanine in the alanine, aspartate, and glutamate metabolism pathway (Fig. 4), where aspartate has a decreased (fc = 0.91) and alanine has a slightly elevated (fc = 1.06) level in dementia patients. A study of human AD brain extracts reported similar alterations of these metabolite levels and identified the same pathway as being the most affected [43]. In a recent study using serum, a significant decrease in the aspartate concentration was also observed for subjects with AD or mild cognitive impairment when compared to healthy control subjects; however, no significant change was observed for alanine [13]. A second study on serum also found a reduced concentration of aspartate in AD patients [45].
Alanine is also closely linked to acetic acid within the taurine and hypotaurine pathway (Fig. 4), a route previously implicated in our metabolomics study of saliva from dementia patients and healthy control subjects [25], and in agreement with that study the present study on serum showed an increased concentration of acetic acid in dementia patients (fc = 1.08). Dysregulation of the same pathway was previously suggested based on the alteration of metabolite levels in brain extracts from AD patients and healthy control subjects, among them an elevated alanine concentration [46]. However, the altered level of acetic acid might also be related to its involvement in other pathways such as pyruvate and glycolysis/gluconeogenesis metabolism. Interestingly, the pathways discussed above are all linked together as illustrated in (Fig. 4), where threonine and threonine metabolism are of particular interest.
Several additional discriminatory metabolites were identified that are not directly linked to threonine metabolism. Glutamine showed a slightly increased concentration in pre-diagnostic dementia subjects (fc = 1.04), which agrees with studies of blood and brain extracts from dementia and AD patients [18, 43], but disagrees with a study of serum from AD patients [45]. Glutamine participates in numerous physiological functions, such as protein biosynthesis and energy production, and it plays a major role in the synthesis of the neurotransmitters glutamate, aspartate, and γ-aminobutyric acid (GABA). It is consequently involved in the glutamine-glutamate/GABA cycle [47] and thereby indirectly affects the GABA shunt, a pathway known to be affected in early stages of AD [48].
Tyrosine had a slightly reduced serum concentration in pre-diagnostic dementia patients when compared to controls (fc = 0.94), which previously has been observed in serum [13, 45] and plasma [16] from AD patients. An altered level of tyrosine was not unexpected because tyrosine has an important role for neurotransmission as a precursor for the neurotransmitters dopamine, epinephrine, and norepinephrine [49].
Trimethylamine, which is primarily produced by gut microbiota, showed an increased level in the pre-diagnostic dementia patients (fc = 1.26) compared to healthy individuals. This metabolite is usually associated with cardiovascular disease [50], a disorder known to share risk factors, mechanisms, and pathophysiology with dementia [51].
Two metabolites with increased levels in pre-diagnostic dementia patients were isobutyrate (fc = 1.10) and propylene glycol (fc = 1.11). Neither of the two has previously been implicated as a marker for dementia, and no specific metabolic pathway important in dementia could be linked to the two metabolites. Further investigations are thus necessary to confirm these findings.
In the dementia substudy (SS1), the level of acetoacetic acid was decreased (fc = 0.82) in dementia patients. This might indicate a disturbance in ketone body synthesis and degradation that might be related to deteriorated glucose metabolism of the brain in dementia patients.
Comparison with our previous NMR- and MS-based studies
A subset of the serum samples was previously characterized using an MS-based method, resulting in discrimination between the dementia and control groups and the identification of 3,4-dihydroxybutanoic acid, docosapentaenoic acid, and uric acid as discriminatory metabolites [24]. None of these serum metabolites were possible to detect with the NMR-methods used in the present study, which prevented confirmation of previous results.
Saliva from a subset of the subjects was also studied by both NMR- and MS-based methods [24, 25], but only the NMR study was able to discriminate between groups. Among the discriminatory saliva metabolites, which also showed a weak pre-diagnostic capability, only acetic acid was found in the present serum NMR-study, and in both cases an elevated level was found in the dementia group. Interestingly, even though only one overlapping metabolite was identified, there were two common metabolic pathways implicated in both the saliva and serum NMR-studies – the taurine and hypotaurine pathway and the GABA shunt.
Concluding remarks
In the present NMR-based comparison of the serum metabolite profiles of dementia, pre-diagnostic dementia, and healthy control subjects, several discriminatory metabolites were identified, where threonine was found to be of particular importance. In the pre-diagnostic dementia substudy, a disturbance in threonine catabolism was detected, which in the dementia substudy was expanded to include several metabolic pathways that directly or indirectly involve threonine (threonine catabolism; the glycine, serine, and threonine pathway; the alanine, aspartate, and glutamate pathway; and the taurine and hypotaurine pathway) (Fig. 4). Furthermore, the GABA shunt and the neurotransmitter synthesis pathway appear to be important at the pre-diagnostic dementia stage, and ketone body synthesis appears to be important at the dementia stage. Several of these metabolites and pathways have been implicated in previous studies of dementia and AD, and therefore these studies and the present study support each other. Replication of the present findings in a separate independent cohort is however necessary for validation. Finally, the present study provides a valuable contribution to the search for metabolic markers for dementia, as well as clues to the mechanisms behind the syndrome.
Footnotes
ACKNOWLEDGMENTS
We are indebted to the subjects who contributed to this study. We are grateful to the Erling-Persson Family Foundation for funding. The Betula data collection was supported by a grant from the Knut and Alice Wallenberg Foundation to Prof. Lars Nyberg. Dr. Anders Öhman was supported by grants from the Erling-Persson Family Foundation, the Alzheimer Foundation in Sweden, Insamlingsstiftelsen at Umeå University, and the Foundation for Clinical Neuroscience at Umeå University Hospital. Dr. João Figueira is grateful for a post-doctoral fellowship from the Kempe Foundation. We would like to thank Prof. Lars-Göran Nilsson (Aging Research Center, Karolinska Institutet, Sweden) for managing the Betula project and for generous support. The Kempe Foundation and the Knut and Alice Wallenberg foundation are acknowledged for their support of the NMR infrastructure at Umeå University (NMR4life).