
Abstract
Background:
Both advanced glycation end products (AGEs) N-(1-carboxymethyl)-L-lysine (CML) and pentosidine were found in the brain from Alzheimer’s disease (AD) patients and were associated with the neuropathological hallmarks of AD. In AD patients, the circulating level of both AGEs remains unknown. Moreover, their levels in peripheral extracellular vesicles (EVs) and their association with AD remain to be determined. Finally, it is not known if neuronal cells can release AGEs via EVs and propagate AGEs.
Objective:
To determine the levels of circulating CML and pentosidine during the progression of AD. Moreover, their levels in circulating EVs were determined and their association with the clinical cognitive scores were analyzed. Finally, we have studied the possibility that neuronal cells eliminate and transfer these AGEs through EVs.
Methods:
CML and pentosidine levels were measured in serum and in circulating EVs. Released-EVs from SK-N-SH neuronal cells were isolated and CML levels were also determined.
Results:
The levels of CML in albumin-free serum proteins were higher in the early stage of AD while the levels of pentosidine remained unchanged. In contrast, the levels of CML in the EVs were lower in the moderate stage of AD. Interestingly, the levels of CML in serum were negatively correlated with the clinical cognitive scores MMSE and MoCA. For the first time, we were able to demonstrate that CML was present in EVs released from neuronal cells in culture.
Conclusion:
Peripheral and circulating EVs levels of CML can differentiate early to moderate AD. In the brain, neuronal CML can propagate from cells-to-cells via EVs.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is the most common neurodegenerative disease. It is estimated that more than 50 million people lived with dementia worldwide, and its prevalence rises to 152 million in 2050, in the absence of prevention strategies and effective treatments [1]. The amyloid-β (Aβ)-containing plaques and the phosphorylated tau-containing neurofibrillary tangles represent both neuropathological hallmarks of AD.
It is now well recognized that AD dementia is preceded by a long asymptomatic (preclinical) phase followed by a transitional stage with mild cognitive impairment (MCI) [2]. The transitional stage offers an important opportunity for prevention and therapeutic interventions. Currently, the probable AD diagnosis is based on a combination of cognitive tests, neuroimaging and the p-tau/Aβ42 ratio in cerebrospinal fluid (CSF). However, the specificity and sensitivity of these criteria are relatively limited because the amyloid plaques and neurofibrillary tangles are necessary but not sufficient for the development of AD. In addition, the present diagnosis represents a late stage associated with an irreversible brain damage. Thus, the identification and development of specific biomarkers to detect the disease as early as possible represents a great challenge [3].
Many reports have demonstrated the implication of advanced glycation end products (AGEs) in the pathophysiology of AD. AGEs are implicated in diabetic, metabolic syndromes, in aging and cardiovascular complications, which are also well-known risk factors for AD development [4 –8]. Accordingly, in AD brain, AGEs were found in pyramidal neurons of human hippocampus with an accumulation in endosomes and lysosomes [9, 10]. The interaction between AGEs and their receptors RAGE induces the generation of intracellular oxidative stress, release of pro-inflammatory cytokines as IL6 and TNF-alpha, and increases Aβ production and tau hyperphosphorylation [11]. In an animal model, an intracerebroventricular injection of glycated Aβ caused memory deficit in rats [12].
AGEs are a heterogeneous class of molecules formed nonenzymatically through a series of Maillard reactions, between glucose, or other reducing sugars, and free amino groups such as lysine and arginine residues of proteins [13]. Alternatively, these compounds can be formed via the interaction between arginine, lysine, cysteine, or tryptophan residues of different proteins and dicarbonyl compounds such as methylglyoxal or glyoxal [14]. The reaction of methylglyoxal with arginine leads to the formation of cyclic imidazolone adducts [15] and other related structural isomers. Its reaction with lysine residues generates the N-(1-carboxyethyl)-L-lysine (CEL) and N-(1-carboxymethyl)-L-lysine (CML) adducts and the lysine dimer 1,3-di(N-lysino)-4-methyl-imidazolium (MOLD) [16]. Pentosidine, another AGE, is mainly formed in reactions of pentoses with lysine and arginine. Both CML and pentosidine are generated by an oxidation-dependent mechanism.
In human brain, CML was found in the cytoplasm of neurons with higher expression in hippocampus than in frontal and temporal cortex. In AD, CML and pentosidine are co-localized with microglia and positive GFAP (glial fibrillary acidic protein) astrocytes [17] with the expression of CML being inversely correlated with the Braak staging [18, 19]. CML was identified in the neuronal perikarya in neuritic plaques but not in the core of amyloid plaques, while pentosidine was present in the core of senile plaque and in the fiber-like structure within the neuropil [20]. Neurons which display diffuse cytosolic AGEs and particularly CML immunoreactivity also contain hyperphosphorylated tau, suggesting a link between AGEs accumulation with the formation of early neurofibrillary tangles [21]. With age, CML was found to be increased in the four regions of the brain, the cerebral cortex, midbrain, striatum, and hippocampus [22]. In AD, the accumulation of CML and pentosidine in the CA1 region of the hippocampus is higher than in controls [23 –25]. Altogether, these specific AGEs formed under oxidative conditions [26, 27] seem to be a major contributor to the initiation and development of age-related AD. Taken together, AGEs and particularly CML and pentosidine could be considered as important factors in the development of AD.
Previous studies have evaluated the levels of pentosidine and CML in the serum and CSF from AD patients. However, the results in CSF were controversial and the quantification of pentosidine and CML levels in the serum was performed in total proteins with albumin [28 –31]. Albumin, the most abundant circulating protein in serum, may be exposed to glycoxidation which may lead to the formation of various types of AGEs. Thus, we hypothesize that the quantification of total AGEs in serum containing albumin would not be related to AD pathology.
Extracellular vesicles (EVs) such as exosomes play a crucial role in the secretion and transport of multilevel information, including signaling factors, misfolding proteins, nucleic acid, and regulatory molecules. EVs are small vesicles secreted by all cell types including neurons, astrocytes, and microglia [32 –34]. Several studies have suggested the implication of EVs in the propagation of pathogenic proteins in AD. Other studies have demonstrated the presence of AD hallmarks (Aβ peptide and phosphorylated tau protein) in peripheral EVs in AD patients [35 –38]. However, the presence of AGEs in peripheral EVs and their association with AD remain to be determined. Moreover, the propagation of AGEs and particularly CML and pentosidine, in the brain by neuronal cells-derived EVs remains to be demonstrated.
The aim of present study was designed to compare the circulating levels of pentosidine and CML between controls, MCI, and AD patients during the progression of the pathology. Then, the association between these peripheral levels of AGEs with two clinical scores, the Mini-Mental State Examination (MMSE) and the Montreal Cognitive Assessment (MoCA) were analyzed. Furthermore, we have explored the possibility of the release of CML by neuronal cells in culture through EVs.
MATERIALS AND METHODS
Study design and participants
Serum samples from control subjects, MCI, and AD patients were generously given by the Memory Clinic of the Sherbrooke. The main characteristics of the population enrolled in this study are summarized in Table 1. The healthy control subjects were defined according to the SENIEUR protocol [39]. Selection of MCI subjects was based on cognitive test scores and Pertersen criteria [40]. The AD patients were selected depending on criteria developed by the National Institute of Neurological and Communicative Disorders and Stroke and the Alzheimer’s Disease and Related Disorders Association (NINCDS-ADRDA) and the fourth edition of the Diagnostic and Statistical Manual of Mental Disorders (DSM-IV) published by the American Psychiatric Association in 1984 [41]. The study protocol was approved by the Ethics Committee of the University of Sherbrooke and informed written consent was obtained from all participants and/or their representatives (protocol #2010-21/Fülöp). All selected subjects, except for the late AD patients, underwent clinical and neuropsychological evaluation included the MoCA and MMSE [42, 43]. Blood was obtained after overnight fasting in heparin-containing vacuum tubes. Blood samples obtained from selected subjects were centrifuged at 3000 rpm for 10 min at 4°C. Blood glucose, albumin, total cholesterol, HDL cholesterol, LDL cholesterol, and triglyceride were measured. Serum were collected and stored at –80°C until analysis.
Characteristics of patients and controls participants
Values are mean±standard error of the mean (SEM). AD, Alzheimer’s disease; ES, early stage of Alzheimer’s disease; MS, Moderate-stage of Alzheimer’s disease; LS, late-stage of Alzheimer’s disease; MMSE, Mini-Mental State Examination; MoCA, Montreal Cognitive Assessment; BMI, body mass index; HDL, high density lipoprotein; LDL, low density lipoprotein; ND, not detected. MMSE and MOCA scores are not detected in LS patients because of their incapacity to accomplish the tests. Statistical analysis was performed using the one-way ANOVA followed by LSD test with a p < 0.05, b p < 0.01, c p < 0.001 versus controls subjects.
Isolation and characterization of extracellular vesicles from serum samples
Serum EVs were isolated by precipitation method using the Total Exosome Isolation reagent (Invitrogen™ by Life Technologies Inc., Carlsbad, CA, USA) [44] with minor modifications. For this, serum samples from all groups were clarified by centrifugation at 10,000×g for 20 min to remove debris. 1/5 volume of exosome isolation reagent was added to supernatant and samples were refrigerated at 4°C for 30 min. Then, after centrifugation at 10,000×g for 5 min, EVs pellets were re-suspended in PBS and purified by a series of filtrations and precipitations. Final EVs preparations were re-suspended in PBS, divided in aliquots and conserved at –80°C for further analysis. EVs proteins were extracted using Radio Immuno Precipitation Assay (RIPA) buffer (50 mM Tris buffer, pH 8, 150 mM sodium chloride, 0.1% sodium dodecyl sulfate, 1% Igepal, 1% sodium deoxycholate, 5 mM EDTA, 1% protease and phosphatase inhibitor cocktail). The total protein concentration was measured using bicinchoninic acid (BCA) assay (Pierce™ BCA Protein Assay Kit, ThermoFisher Scientific, Inc).
Transmission electron microscopy (TEM)
Transmission electron microscopy was used for morphological examination of isolated EVs. EVs preparations were suspended in 2% paraformaldehyde. Ten microliters of mixture were loaded on Formvar-carbon coated grid for 5 min. Grids were negatively stained using 2% uranyl acetate solution for 1 min. The excess fluid was gently removed and grids were observed using the HITACHI 7100 transmission electron microscope (75 kV) at 15000X–40000X magnification.
Nanoparticle tracking analysis (NTA)
Sizes and concentrations of EVs preparations were analyzed using a Nanosight NS300 instrument and the Nanosight NTA 3.2 Analytical Software (Malvern Instruments Company, Nanosight, and Malvern, United Kingdom). Diluted EVs suspensions were illuminated by laser light. The light scattered by the particles was captured using a digital camera. The motion of each particle is tracked from frame to frame by the Nanosight NTA 3.2 Analytical Software.
Immunoblot analysis
Twenty micrograms of total EVs proteins were loaded in 10% polyacrylamide gel for electrophoresis. After the separation of proteins according their molecular weight, gel was transferred onto PVDF membranes using a Trans-Blot Turbo System (Bio-Rad). Membranes were blocked for 60 min in TBS (Tris-buffered saline) containing 0.1% Tween 20 with 5% skim milk and incubated with primary antibodies: TSG101(tumor susceptibility gene 101) (MyBiosource; 1/2500) or calnexin (Santa Cruz; 1/500) overnight at 4°C. Then, membranes were washed with TBS-Tween 0.1% and incubated with the secondary antibody HRP-conjugated anti rabbit (Cell Signaling Technology; 1/5000) and anti-mouse (Cell Signaling Technology; 1/1000) for 60 min. Detection was realized with ECL substrate (Bio-Rad Laboratories, Inc., Hercules, CA, USA) and bands were analyzed using the luminescent imaging system FluorChem.
Cell culture, isolation, and characterization of extracellular vesicles in culture media
Human neuroblastoma cell line SK-N-SH from ATCC (Manassas, VA, USA) was used to isolate EVs. Cells were cultivated in Eagle’s minimal essential medium (EMEM) supplemented with 10% (v/v) Fetal Bovine Serum (FBS), 1% sodium pyruvate (1 mM), and antibiotics (100 U/ml penicillin and 100 μg/ml streptomycin) in 37°C with 5% CO2. Cells were grown to 80% of confluence, washed with PBS and incubated in FBS free media for 24 h. The day after, cell culture supernatant was collected for EVs isolation and cellular total protein was extracted with RIPA buffer containing a cocktail of protease and phosphatase inhibitors. Total proteins concentrations were measured using BCA assay.
Dead cells and cell debris were removed from medium by successive centrifugations (300 g for 10 min, 2000 g for 10 min, and 10000 g for 30 min). The final supernatant was concentrated and precipitated with the Total Exosome Isolation reagent (Invitrogen™ by Life Technologies Inc., Carlsbad, CA, USA) to collect the pellet that contains EVs. The EVs pellet was resuspended in PBS and characterized with TEM, NTA and western-blot analysis as described above. Total proteins from SK-N-SH cells and EVs cells were extracted with RIPA buffer containing a cocktail of protease inhibitors and were measured using BCA assay.
Enzyme-linked immunosorbent assay (ELISA)
Levels of pentosidine and CML in serum were determined by competitive ELISA provided by MyBioSource, Inc., according to the manufacturer’s instructions. The detection range of pentosidine and CML kits was 6.2 to 400 ng/ml and 15.6 to 1000 ng/ml, respectively. To detect and measure CML levels in EVs from all study groups and from SK-N-SH cells, sandwich ELISA provided by Cusabio Biothech Co., Ltd., was used according to the manufacturer’s instructions. Detection range of CML kit was 62.5–4000 pg/ml.
Statistical analysis
Data were expressed as mean±SEM. Statistical analyses were performed with the using the SPSS program using one-way ANOVA analysis followed by LSD test and Student t-test. The statistical correlation was determined by a linear regression analysis using the Pearson coefficient of correlation. p value less than 0.05 was considered statistically significant.
RESULTS
Main characteristics of patients and control participants
The main general characteristics of the five groups of participants are reported in the Table 1. MCI and AD participants are older than controls. Mean and standard error of MMSE and MoCA scores were significantly lower in MCI and AD groups compared to control subjects.
Although blood glucose levels were in the physiological range, they were higher in MCI and in moderate AD participants compared to controls. To note, albumin levels in severe AD participants were lower than in controls but remain in the physiological range of circulating levels. The BMI value, the HDL, LDL, TG, and total cholesterol levels were not different between groups.
Determination of pentosidine and CML levels in serum
Serum proteins are divided in two groups, high and low abundance circulating proteins, with albumin being among the most abundant protein. This circulating protein may be exposed to increased glycoxidation stress leading to the formation of various types of AGEs. Therefore, we have measured and compared pentosidine and CML levels in the total serum and in the low abundance circulating proteins without the albumin fraction (S-Ab).
CML levels in the total serum proteins were not different among five groups (Fig. 1A). However, CML levels in the S-Ab fraction were significantly higher in AD group (92.6±9.94 ng/mg protein) and specifically in early AD group (101.6±19.19 ng/mg protein) as compared to the control group (56.95±6.55 ng/mg protein) (Fig. 1B). Pentosidine levels in the total serum proteins and in S-Ab fraction were not different between the five groups (Fig. 2).

Serum CML levels in controls, MCI and different groups of AD patients. A) CML levels in total serum proteins. B) CML levels expressed in total serum proteins without albumin (S-Ab). Difference between groups were analyzed with one-way ANOVA followed by the LSD post-hoc test. Values are mean±SEM with a p < 0.05 versus Controls subjects.
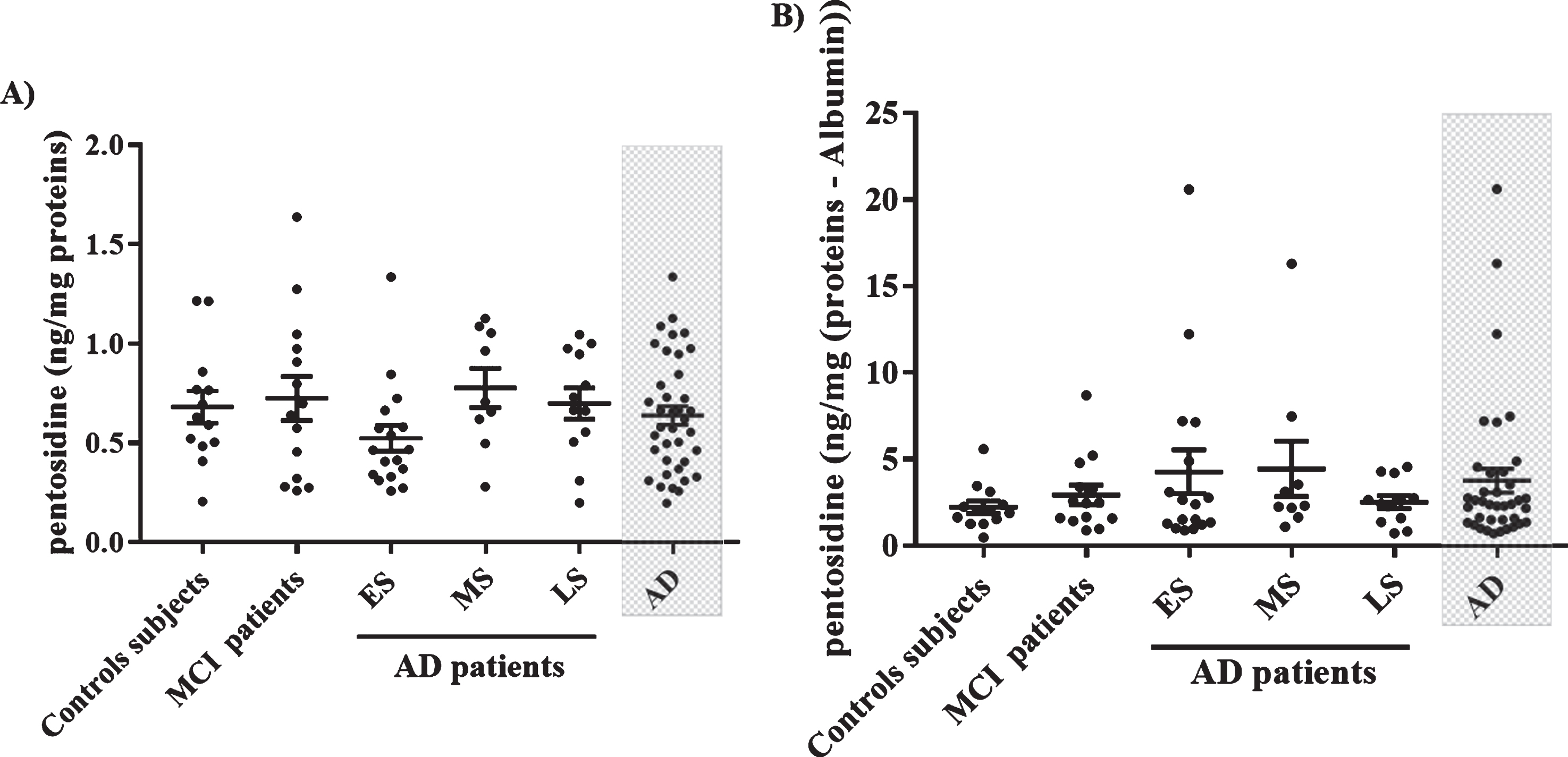
Serum pentosidine levels in control subjects, MCI, and AD patients. A) Pentosidine levels in total serum proteins. B) Pentosidine levels expressed in total serum proteins without albumin (S-Ab). Data groups were compared with analysis of variance one-way ANOVA followed by LSD post hoc test. Values are mean±SEM.
Interestingly, CML levels in S-Ab were negatively correlated with the MMSE and MoCA scores (Fig. 3A, B) while pentosidine levels were not (Fig. 3D, E). Pentosidine levels in S-Ab were age-dependent (Fig. 3F) but not CML (Fig. 3C).

Statistical correlation between CML and pentosidine levels expressed in total serum proteins without albumin (S-Ab) and cognitive scores (MMSE and MoCA) and age (years) obtained with the Pearson correlation.
CML levels in EVs from total serum
Circulating EVs were isolated from control subjects, MCI, and AD patients. The characterization of EVs was based on the analysis of the morphology by TEM, their sizes distribution by the Nanosight system NTA and the presence of exosomal markers TSG101 by western blot (Fig. 4). TEM images showed that vesicles were surrounded by a lipid layer with a cup-shaped morphology and a size being between 80 to 100 nm diameter (Fig. 4A). The NTA analysis showed that the density of the most abundant particles has a diameter ranging between 50 and 150 nm (Fig. 4B). Our EVs fraction contained protein TSG101 while calnexin, a negative marker of EVs was absent. SK-N-SH neuronal cells lysate was used as a positive control for the calnexin expression (Fig. 4C).

Characterization of EVs isolated from the serum. A) The TEM image depicts the spherical morphology of the isolated particles, bar = 100 nm. B) Determination of concentration and size of isolated EVs in the study population by NTA. C) Representative blot of extracellular vesicles TSG 101 and calnexin in control subjects, MCI and AD patients and in SK-N-SH cells lysate.
Our results showed that circulating EVs contained CML. Moreover, CML levels in EVs were lower in moderate AD as compared to control subjects with 38.52±5.69 pg/mg proteins and 67.31±8.99 pg/mg proteins, respectively (Fig. 5). To note, levels of CML were 1000-fold higher in serum as compared to EVs levels.

CML levels in EVs from control subjects (n = 8), MCI (n = 7), and AD patients (ES, n = 8; MS, n = 7; LS, n = 8). Data groups were compared with analysis of variance one-way ANOVA followed by LSD post hoc test. Values are mean±standard error of the mean. a p < 0.05 versus controls subjects.
Isolation and characterization of neuronal derived-EVs and CML detection
In order to demonstrate that neuronal cells are able to release CML in EVs, we isolated these vesicles from the human SK-N-SH neuronal cells culture supernatant. As in serum-derived EVs, TEM images revealed that these vesicles are surrounded with a lipid layer producing a cup-shaped morphology (Fig. 6A). The expression of EVs marker was also confirmed by the presence of TSG101 and the absence of calnexin (Fig. 6B). The size distribution of these extracellular vesicles was similar to those from serum (Fig. 6C). Interestingly, our results demonstrated that neuronal cells can release CML in EVs with CML levels in vesicles being ∼2.5 fold lower than in cell lysates (Fig. 6D).
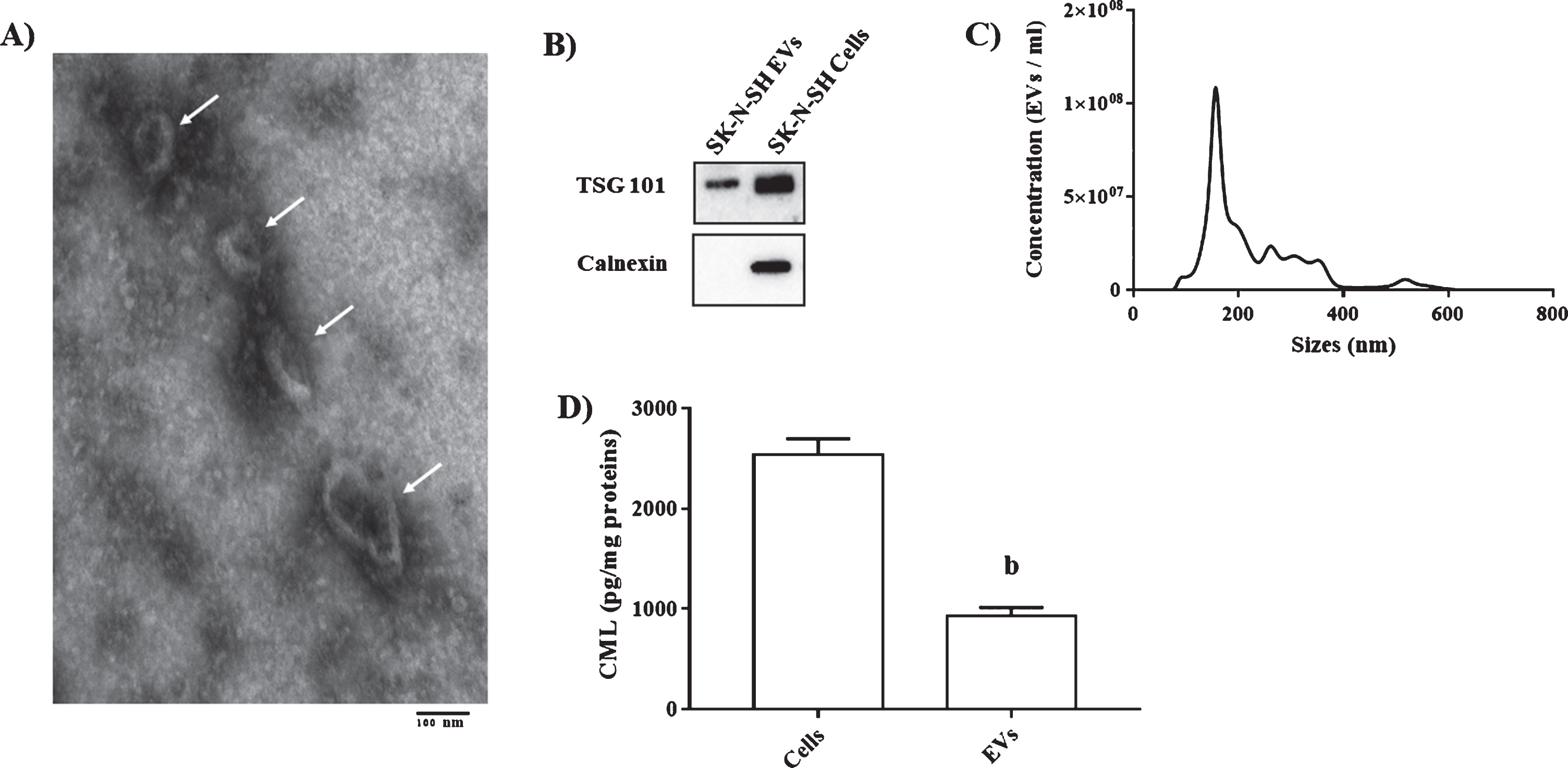
Characterization of cells EVs and CML determination in cells lysate and EVs. A) The TEM image showed the spherical morphology of the isolated particles, bar = 100 nm. B) Representative blot of TSG 101 and Calnexin in EVs from SK-N-SH and cells lysate. C) Concentration and size of isolated EVs by NTA. D) CML levels in EVs from SK-N-SH and cells lysate. Comparison between different groups were performed with Student t-test. b p < 0.01.
DISCUSSION
To date, the absence of validated peripheral blood-based biomarkers of AD limit the early detection, the evaluation of the progression of the disease, and the development of efficient pharmacotherapy. The actual challenge is to link peripheral biomarkers to brain pathology and cognitive decline. In the present study, we showed that serum levels of CML in S-Ab were higher in AD patients and particularly in early AD. We found that peripheral CML levels were negatively correlated with both cognitive scores MMSE and MoCA. For the first time, we identified that CML are released in circulating EVs. Moreover, we demonstrated that CML is also present in neuronal cells-derived EVs. In contrast to CML, circulating levels of pentosidine were not different between controls and AD patients and were thus not related to the decline of cognitive scores.
There is an abundant literature suggesting that AGEs play a pivotal role in the development of AD. For instance, in AD brain, Aβ peptide and tau proteins could be the target of glycation leading to their resistance toward hydrolysis by proteases. Aβ-AGEs were found both in vitro and in vivo [12] and the nucleation-dependent polymerization of Aβ was significantly accelerated by crosslinking with AGEs [45]. The intrahippocampal injection of AGEs, obtained from BSA-glucose, induced tau hyperphosphorylation, memory deterioration, decline of synaptic proteins, and impairment of long-term potentiation (LTP) through RAGE-mediated glycogen synthase kinase-3 (GSK3β) activity [46].
AGEs are heterogeneous class of molecules. The challenge is to determine if some specific AGEs could propagate from cell-to cell and if specific circulating AGEs is elevated in MCI or during the progression of the disease. Among different AGEs, we have focused on CML and pentosidine because both were detected in AD hallmarks [23–25 , 47]. For instance, CML was shown to be intense in neurofibrillary tangles [23] likely because tau is a lysine-rich protein. CML was also abundant and elevated in CSF from AD patients, while pentosidine levels were decreased or remains unchanged [29, 30]. Finally, CML represents the dominant AGEs in human [48]. Our data showed that circulating CML levels do not differentiate MCI to control patients, but it was increased in AD patients. Our results contrast with the previous study from Bar et al. [29], likely due to the number of AD patients (15 versus 35 in our study) and to the composition of the AD group. In the previous study [29], AD group included mainly moderate and severe AD and the average MMSE scores was 16.3±6.4, while in the present study the elevation of CML was observed only in early AD with the MMSE score being between 23±0.9. Hence, our results showed that peripheral levels of CML could differentiate AD from controls and particularly early AD but their levels in circulating EVs was lower specifically in moderate AD.
In total serum proteins from AD patients, pentosidine levels remain controversial. They were not different between controls and AD or increased in converted MCI subjects to AD patients and remain elevated in AD [28 , 31]. Our results in S-Ab fraction confirm that pentosidine levels were not different between control, MCI, and AD groups.
It is well-known that high circulating levels of AGEs represents an overall change of physiological functions with an increase of their formation during various chronic diseases such as inflammation, diabetes, metabolic syndrome, insulin resistance, endothelial dysfunction, cardiovascular complication [4 , 49–51], or aging [52 –54] and is not specifically related to AD. However, these chronic diseases represent the risk factor for the development of AD. As these chronic diseases were not included in our study, our results indicate that the elevation of peripheral CML in early AD and in AD group could be specifically associated to AD which was reinforced with the negative correlation with the cognitive scores.
The presence of peripheral AGEs depends on several factors, including the metabolic rate, the redox state, the small fraction of dietary absorbed AGEs, and the turnover rate of the targeted proteins or cells excretion. The abundance of CML could be explained by the multiple pathways leading to its formation such as the reaction between glucose, other sugars or glyoxal with amino groups of lysine, or by-product of lipids peroxidation and oxidized proteins while pentosidine (protein cross-link) could be formed by the reaction of ribose, glucose, fructose, or ascorbate with lysine and arginine [55]. The elevation of CML in early AD observed in this study could be related to the alteration of the antioxidant/oxidant homeostasis and to oxidative damage recently observed in the same group of patients, with an early decrease of the total antioxidant capacity in MCI and an increase of the protein carbonyls observed in early AD [56]. Similarly, several previous studies have also demonstrated that oxidative-induced damages increased early in AD [57 –63]. In addition, CML is also a RAGE receptor ligand. The interaction of CML with RAGE causes the generation of intracellular oxidative stress, release of proinflammatory factors as IL6 and TNF alpha and increase Aβ production and tau hyperphosphorylation [64, 65]. Hence, the increase of circulating CML could the consequence of the modification of the redox environment which could, in turn, induce oxidative stress.
Albumin is the major circulating proteins and most of AGEs are composed of glycated albumin protein. Therefore, we have determined the levels of CML and pentosidine in the total serum proteins and compared with their levels in the low abundance circulating proteins without the albumin fraction. Interestingly, our data showed that they are higher in S-Ab. Moreover, when CML levels were determined in the low abundance proteins in serum, they are elevated in early AD and are negatively associated to cognitive decline. Hence, the level of CML in S-Ab is a better representation of the cognitive status than the total circulating CML.
In AD brain, Aβ peptides and tau protein can be glycated and formed CML. This complex Aβ-CML or Tau-CML can also efflux through the blood-brain barrier (BBB) to the circulation [66]. When AD progress, brain glycated proteins and peptides accumulate and form aggregates that cannot cross the BBB. This property can explain the elevation of circulating CML levels only in early AD and not with the progression of the disease. However, the explanation of the variability of CML and pentosidine levels in blood and brain, need further investigations.
Recently, it has been suggested that EVs released by brain cells are able to cross the BBB and can be detected in blood [67]. As we have demonstrated that the EVs-derived human neuronal cell-line SK-N-SH contained CML, thus we cannot exclude the possibility that a fraction of the CML-containing EVs in the circulation were from the brain. These results are of great interest because more and more studies suggest that the EVs are responsible of the propagation of pathological proteins such as phosphorylated tau, Aβ peptide, or α-synuclein oligomer in neurodegenerative diseases. They can also play a role in communication across the BBB between the brain and periphery [37 , 68–70]. AGEs formation promotes aggregation and intracellular accumulation of proteins. We hypothesize that with AD progression there is an accumulation of glycated proteins and peptides in the brain which form aggregates that cannot to be released by EVs. Therefore, decreased EVs CML in MS of AD can be explained by accumulation of CML-proteins in the brain.
These results strengthen the notion that peripheral EVs might serve as relevant material for developing biomarkers for the central nervous system diseases.
Conclusion
In conclusion, the present study demonstrated that the association between the serum/S-Ab CML levels and cognitive scores. It can be a useful diagnostic markers for early stage of AD. In the brain, some CML can propagate from cells to cells by EVs and further studies are required to better understand the roles of specific AGEs in EVs.