
Abstract
Certain proteases are involved in Alzheimer’s disease (AD) and their erroneous control may contribute to the pathology onset and progression. In this study we evaluated the cerebral expression of eight proteases, involved in both AβPP processing and extracellular matrix remodeling. Among these proteases, ADAM10, ADAMTS1, Cathepsin D, and Meprin β show a significantly higher mRNAs expression in sporadic AD subjects versus controls, while ADAMTS1, Cathepsin D, and Meprin β show an increment also at the protein level. These data indicate that transcriptional events affecting brain proteases are activated in AD patients, suggesting a link between proteolysis and AD.
INTRODUCTION
The proteolytic processing of amyloid-β protein precursor (AβPP) and the formation of amyloid-β peptides (Aβ) by β- and γ-secretases are considered pivotal events in the genesis of Alzheimer’s disease (AD) [1]. The processing of AβPP is crucial in AD either considering the “amyloid hypothesis”, which states that soluble Aβ oligomers are the cause of neurodegeneration, or the “cell-cycle hypothesis”, which considers AβPP (and its proteolytic fragments) as signaling molecules that force post-mitotic neurons to re-enter into the cell-cycle [2
–5]. However, besides secretases, other proteases can cleave AβPP with a yet unclear role in AD [6]. Specific endoproteases are often upregulated in AD and are directly involved in the processing of AβPP or in Aβ clearance [7
–11]. In general, matrix metalloproteases, metallo-endopeptidases, or acidic proteases are important mediators in brain development and homeostasis; however, their erroneous control may cause a remodeling of the extracellular matrix (ECM) contributing to neurodegeneration and neuroinflammation [12, 13]. γ-secretase, besides AβPP, may cleave different substrates with a relevant role in neuronal differentiation and migration and in brain homeostasis [14, 15]. In this sense, recent data suggest a link between specific proteases that cleave AβPP and neurodegeneration: ADAM proteases, mainly ADAM10 and ADAM17, cause the α-secretase cleavage of AβPP as well as of cell surface receptors involved in neuronal differentiation and migration, such as Deleted in Colorectal Cancer and Notch [15]. In AD, α-secretase cleavage of AβPP is considered a neuroprotective event, since its product is not amyloidogenic. ADAMTS proteases share with ADAM multiple conserved domains, and elevated protein levels are described in AD, Down’s syndrome (as AβPP also ADAMTS1 is located on chromosome 21), and Pick’s disease [16]. Matrix metalloprotease 9 (MMP9) positively regulates synaptogenesis and plasticity and is progressively upregulated, during the acquisition of the neural traits, in neuroblastoma cells [17
–19] together with an increased amyloidogenic processing of AβPP and Aβ secretion [20]. MMP9 activity is likely increased in AD brains [8, 13], in which was found in pyramidal neurons and near amyloid plaques, showing α-secretase-like activity over Aβ40 [21, 22]. Whether this is a protective condition (improved degradation of Aβ) or not (ECM degradation and inflammation) has to be clarified. Meprin α (MEP1A) and β (MEP1B) are Zn2 +-dependent proteases of the astacin family, known to be expressed in different tumors (mainly Meprin α) [23, 24] and, only recently, have been involved in AβPP’s proteolytic processing at N-terminal side (essentially Meprin β), generating aggregation-prone, N-terminally truncated Aβ peptides: in particular, compared to Aβ40, N-terminally truncated Aβ2 - 40 variant shows increased aggregation propensity and acts as a seed for Aβ40 aggregation [11
, 25–28]. Interestingly, ADAM10 can be activated by Meprin β which, in turn, is shed from the cell surface by ADAM10 [29, 30]. Cathepsins, in particular Cathepsin D (CTSD) and L (CTSL), are intracellular acidic proteases which cleave AβPP and can be overexpressed and secreted in pathological conditions related to cancer (as mitogenic and pro-metastatic molecules [31
–33]) and to neurodegeneration. In fact, Cathepsin D, which accumulates within AD neurons, cleaves at the N-terminal side the first aspartate residue of amyloid β [34
–36]. On the other side, under physiological conditions, a subset of AβPP undergoes alternative processing and the intermediate products are rapidly degraded by Cathepsin L [37].
There are few anecdotal studies that compare the expression levels of mRNA or protein of these proteases in human brain. In this study we analyzed in human frontal cortex from AD and non-AD subjects the expression of these eight proteases involved in neurodegenerative processes, ECM remodeling, and AβPP processing (amyloid formation/degradation), searching for a possible contribution of transcriptional events that may justify an increment/decrease of selected protease in AD versus non-AD brain.
MATERIALS AND METHODS
Human brain samples
Frontal cortices were obtained (see Supplementary Material) at autopsy from clinically and neuropathologically verified cases with the diagnosis of “definite” sporadic AD (SAD; total n = 16, average age 76.44±13.25 years old), according to the CERAD criteria, and aged non-demented subjects (total n = 16, average age 76.06±10.50 years old), in which AD has been excluded by clinical evaluation, autopsy examination, and immunohistochemical analysis (Table 1). At least three pieces of grey matter from frontal cortices were excised under sterile conditions and collected as a unique sample [20].
Demographic and clinical brain information of the analyzed patients
+ to +++ symbols indicate the crescent amount of amyloid plaques. A.A., amyloid angiopathy; B&B, Braak and Braak classification stage; N.A., not available; Neg., negative. ApoE alleles genotypes (ɛ2, ɛ3, and ɛ4) are also provided.
Apolipoprotein E (ApoE) genotyping
ApoE genotyping from cortical section was carried out according to previously described methods [38].
RNA extraction and cDNA synthesis
RNAs were extracted from human frontal cortices using PureLink RNA Mini Kit (Life Technologies, USA), and were reverse-transcribed using High-Capacity cDNA Reverse Transcription kit (Applied Biosystems, USA). The cDNA samples were tested for gDNA contamination: the overall level was≤0.05%.
Real-time PCR (RT-PCR)
Genes (included EIF4A2 and GAPDH, used as control housekeeping genes), accession numbers, probes localization of FAM-MGB Taqman assays (Applied Biosystems, USA) are given in Supplementary Table 1. RT-PCR was performed using ABI PRISM 7900HT platform. Briefly, each sample was run in triplicate in each experimental set under standard amplification conditions up to 45 amplification cycles [40]. Data collection was set at the annealing/extension step. For the RT-PCR analysis, see Supplementary Material.
Western blots (WB)
The preparation of soluble brain fractions from brain homogenates was carried out as previously described [41, 42]. Details about electrophoresis and WB techniques are reported in Supplementary Material. Data are normalized versus the highly stable protein γ-tubulin measured in each sample and are represented as fold of expression versus controls.
Statistical analysis
Data are reported as mean value±standard error of the mean, and statistical significance was examined using the Student’s t-test, with the threshold set at p < 0.05 (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001). All the analyses were performed using GraphPad Prism (v.7; GraphPad Software, USA).
RESULTS
Proteases mRNAs expression in human frontal cortices
In order to evaluate mRNA expression levels of these proteases in frontal cortex samples from age-matched SAD (n = 16) and from non-demented (n = 16) subjects, we performed a comparative analysis by RT-PCR; for each gene analyzed, RQ values, normalized by the two housekeeping genes EIF4A2 and GAPDH, are reported as fold of expression levels versus control group.
Data show that, among α-secretases, ADAM10 mRNA expression is significantly (p < 0.01) higher in SAD (2.25±0.33) compared to control subjects (1.00±0.19) (Fig. 1A), while, ADAM17 mRNA levels are similar between groups (controls = 1.00±0.21 versus SAD = 1.04±0.12) (Fig. 1B).
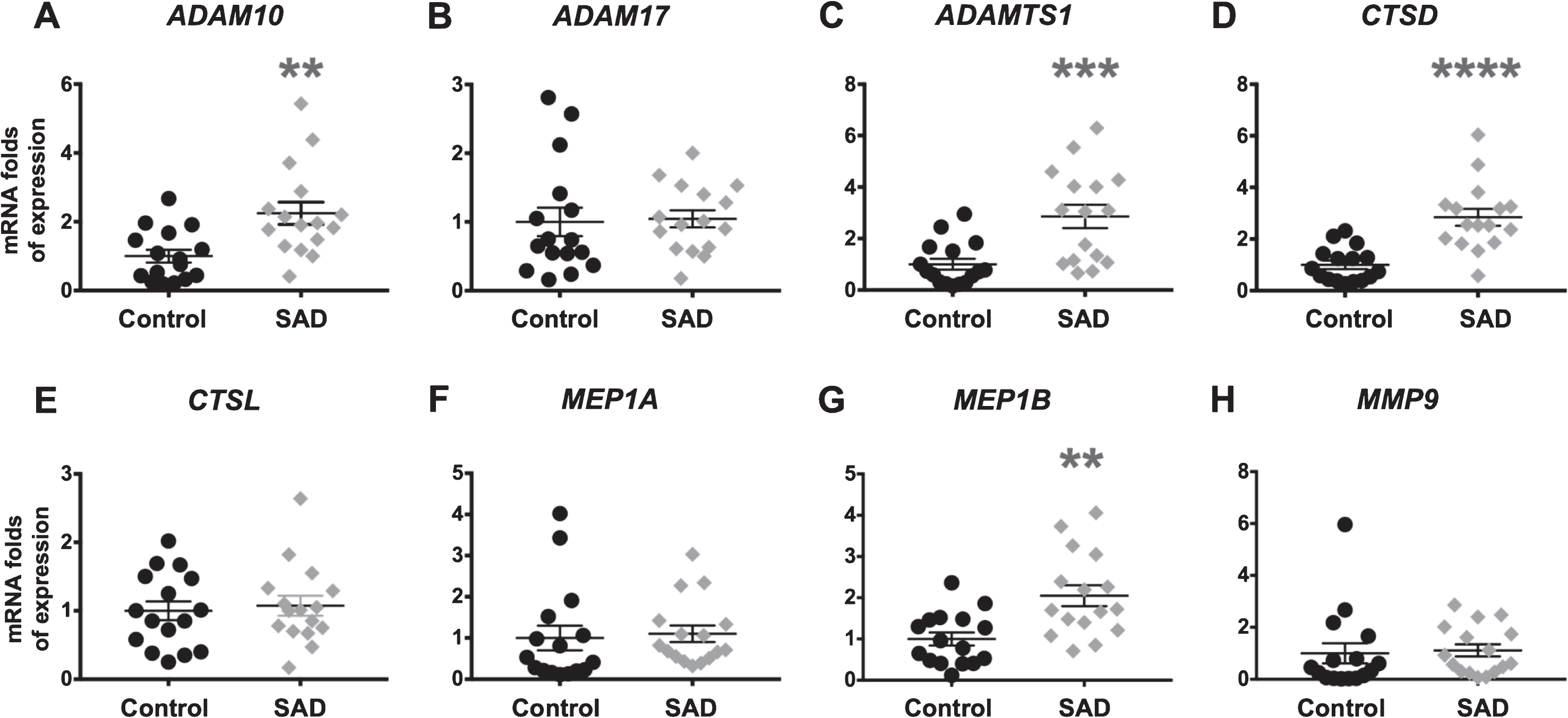
Proteases mRNA content in frontal cortex of age-matched non-demented controls and SAD patients: ADAM10 (A), ADAM17 (B), ADAMTS1 (C), CTSD (D), CTSL (E), MEP1A (F), MEP1B (G), and MMP9 (H). Statistical analysis revealed that, among the proteases analyzed, only the levels of ADAM10, ADAMTS1, CTSD, and MEP1B are significantly higher (2.25-, 2.86-, 2.84-, and 2.05-fold, respectively) in frontal cortex of SAD patients than in control non-demented subjects. Each symbol represents the mean value of a single patient. On the y axis average RQ values are reported as fold of expression respect to controls.
Moreover, there are significant differences between groups for mRNA expression of ADAMTS1 (p < 0.001), CTSD (p < 0.0001), and MEP1B (p < 0.01). In detail, ADAMTS1 is 2.86±0.45 in SAD and 1.00±0.21 in controls (Fig. 1C); CTSD is 2.84±0.33 in SAD and 1.00±0.17 in controls (Fig. 1D); and MEP1B is 2.05±0.25 in SAD and 1.00±0.16 in controls (Fig. 1G).
CTSL, MEP1A, and MMP9 mRNA expression levels did not statistically differ between SAD and control samples (Fig. 1E, F, H): CTSL is 1.00±0.14 in controls and 1.07±0.15 in SAD; MEP1A is 1.00±0.30 in controls and 1.10±0.20 in SAD; and MMP9 is 1.00±0.39 in controls and 1.11±0.24 in SAD.
pro-ADAM10, ADAM10, ADAMTS1, pro-Cathepsin D, Cathepsin D, and Meprin β protein expression in human frontal cortices
In order to ascertain a correlation between differential mRNAs expression and protein level, we analyzed by WB the same cortical samples previously analyzed by RT-PCR looking at the expression of those proteases with a significant differential mRNA expression, hypothesizing a transcriptional mechanism: ADAM10, ADAMTS1, Cathepsin D, and Meprin β.
Cathepsin D undergoes proteolytic cleavage and activation, producing two chains (light and heavy) that are reassembled in the active isoform by disulfide bonds: the anti-Cathepsin D antibody used in this study is able to detect both pro-Cathepsin D and the heavy chain of the active form, upon cleavage of disulfide bridges in denaturant conditions [43, 44]. ADAM10 as well is activated upon cleavage, among the others also by Meprin β [29]. On the basis of these observations, we evaluated also the pro-peptide expression of ADAM10 and Cathepsin D.
As reported in representative WB (Fig. 2A), we detected a relative higher presence of Meprin β, ADAMTS1, and pro-Cathepsin D/Cathepsin D (heavy chain) in SAD versus controls: in detail (Fig. 2B), pro-ADAM10 and ADAM10 expression level are similar between controls and SAD patients (controls = 1.00±0.10 versus SAD = 0.96±0.09 and controls = 1.00±0.15 versus SAD = 1.01±0.14, respectively); ADAMTS1 expression level is 4.34±0.35-fold higher (p < 0.0001) in SAD patients versus controls (1.00±0.11); pro-Cathepsin D and Cathepsin D (heavy chain) are expressed 3.11±0.45-fold (p < 0.001) and 2.84±0.29-fold higher (p < 0.0001) in SAD patients versus non-demented subjects (1.00±0.17 and 1.00±0.11), respectively. Meprin β protein level is significantly 1.82±0.21-fold higher (p < 0.01) in SAD subjects in comparison to non-demented subjects (1.00±0.08). In addition, the average ratios of pro-peptide/active form of ADAM10 and Cathepsin D are similar between control and SAD subjects: 2.34±0.34 versus 2.37±0.43 and 0.79±0.10 versus 0.91±0.15, respectively; however, in contrast to pro-Cathepsin D, which, in both groups, is expressed at similar levels than the active isoform (control = 1.00±0.17 versus 1.35±0.15 and SAD = 1.00±0.14 versus 1.23±0.13), pro-ADAM10 content is more (p < 0.01) represented than mature ADAM10 in both groups (controls = 1.00±0.10 versus 0.55±0.08 and SAD = 1.00±0.14 versus 0.58±0.08).
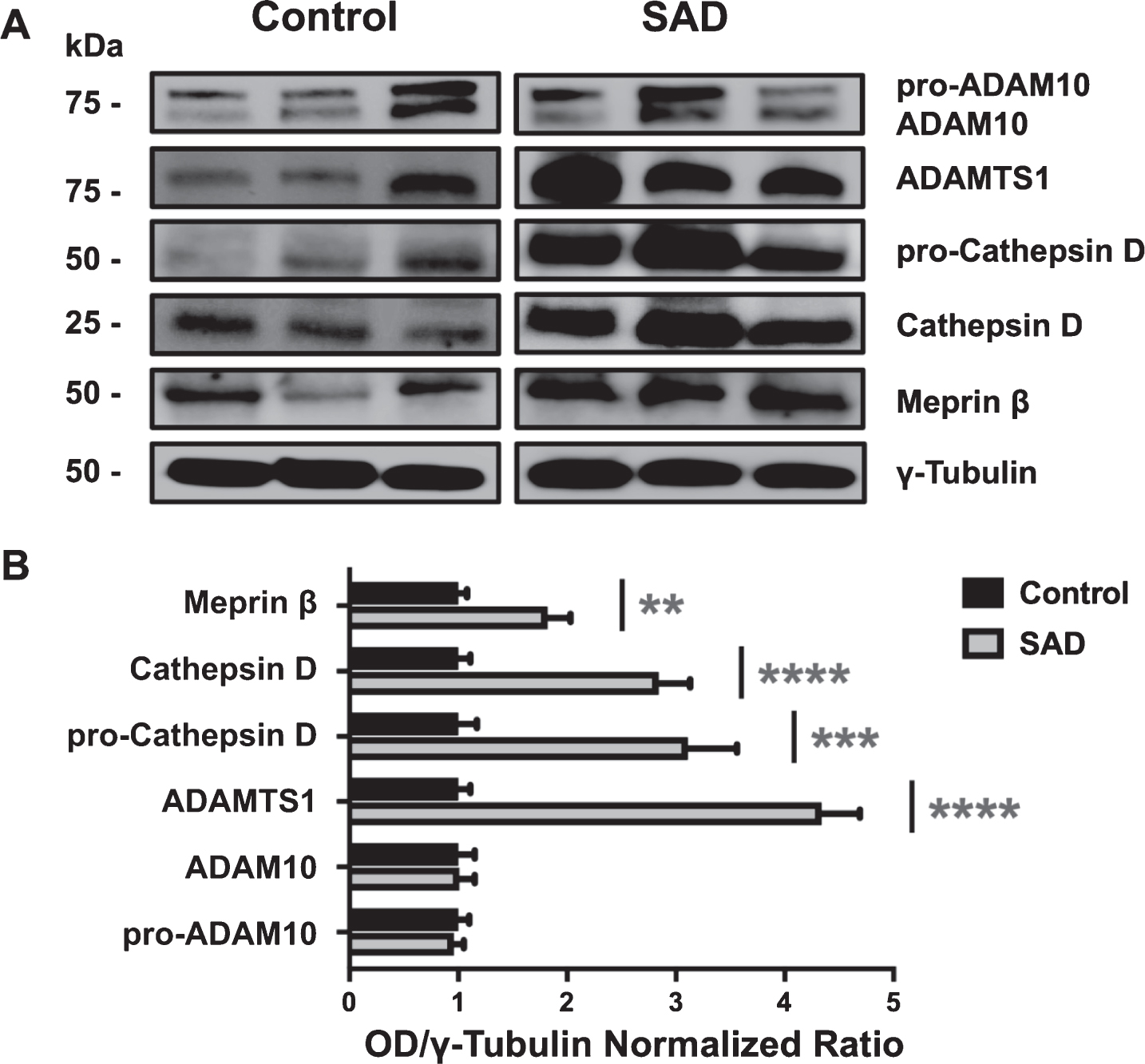
A) Representative western blot of frontal cortex protease levels of pro-ADAM10, ADAM10, ADAMTS1, pro-Cathepsin D, Cathepsin D, and Meprin β of three patients for each group. B) Densitometric analysis reveals significant higher levels of proteases analyzed. ADAMTS1 is 4.34-fold higher (p < 0.0001), pro-Cathepsin D and Cathepsin D are 3.11-fold (p < 0.001) and 2.84-fold higher (p < 0.0001), and Meprin β is 1.82-fold higher (p < 0.01) in SAD patients in comparison to non-demented subjects. Pro-ADAM10 and ADAM10 expression is not statistically significant among the groups.
Conclusions
During the last years, the interplay between ECM and proteases has been recognized as an important issue in brain plasticity, learning and memory, and even in neurodegenerative diseases, such as in AD [12]. The erroneous control of specific proteases and their abnormal activation, may cause the remodeling of the ECM, contributing to brain atrophy, neuroinflammation and decreased dendritic arborization [12, 13] and, in parallel, to the aberrant processing of proteins involved in AD, mainly AβPP [25, 34].
In this work, we measured the expression levels, in human cerebral cortices, of specific proteases known to be involved in the processing of AβPP and in ECM remodeling, looking for a possible correlation between mRNA and protein expression. We detail the increment, at both mRNA and protein level, for ADAMTS1, Cathepsin D, and Meprin β proteases in SAD patients compared to non-demented controls, while ADAM10 is increased only at mRNA level.
Previous works have reported, although in anecdotal manner, the elevated level of some of these proteases in AD brains: ADAM10 (in the hippocampus) and Meprin β mRNAs levels were detected about two-fold higher in AD than in controls, and ADAMTS1 protein showed higher levels in cerebral cortex of AD patients in comparison to non-demented subjects [16 , 45].
Altogether these data clearly indicate that transcriptional (and post-transcriptional) events could positively modulate the levels of these proteases during AD. An interesting case is that of Cathepsin D and Meprin β, where the expression fold of mRNA correlates to that of the protein, suggesting a linear relationship. This feature may have effects on ECM remodeling with consequences on brain plasticity and memory and/or on the aberrant AβPP cleavage which occurs in AD. In fact, Meprin β overexpression may have a direct role in the alternative β-secretase cleavage of AβPP, producing more prone-to-aggregate and more cytotoxic Aβ fragments [25]. Furthermore, in normal conditions, ADAM10 is able to shed Meprin β from cell membrane, limiting its activity on AβPP [11]; upon Meprin β overexpression (and with ADAM10 at normal level) this restraint may be inadequate, raising the levels of BACE-like cleavages induced by Meprin β on AβPP and enhancing the levels of toxic Aβ N-terminally truncated isoforms [11 , 46]. In light of the “cell-cycle hypothesis”, it is interesting also to consider the possible mitogenic activity of pro-Cathepsin D [47, 48]. As far as the possible role of ApoE is concerned, our results are insufficient, and a definite answer would likely require a larger sample size. The same situation reflects the evaluation of a possible “gender effect”; however, analyzing the control group, it seems that there is no clear-cut “gender effect” on proteases mRNA and protein expression.
Obviously, our data represent a sort of “snapshot” taken in AD post mortem brain from subjects in the final stages of the disease. The picture here described suggests that transcriptional events may upregulate selected proteases in course of AD. At present, we cannot ascertain whether these features are cause or consequences of AD; although they suggest a link between aberrant AβPP processing and abnormal ECM remodeling induced by selected proteases. The latter event, in turn, may represent either a contributor to neurodegeneration and inflammation or, on the other side, an extreme neuronal attempt to survive.
Footnotes
ACKNOWLEDGMENTS
This work was supported by the grant “Early diagnosis in Alzheimer’s disease to optimize pharmacological therapy and clinical assistance”- National Priority and National Priority Objectives 2013 - Molise Region to C.R.
The Human Brain and Spinal Fluid Resource Center, VA West Los Angeles Healthcare Center, Los Angeles, CA, USA is sponsored by National Institutes of Health, National Multiple Sclerosis Society and the US Department of Veterans Affairs.