
Abstract
TOMM40 ‘523 is associated with Alzheimer’s disease (AD), but APOE linkage disequilibrium confounds this association. In 170 APOE ɛ3 homozygotes, we evaluated relationships between short and very long TOMM40 alleles and longitudinal declines in three cognitive domains (attention, verbal memory, and executive function). We used factor analysis to create composite scores from 10 individual cognitive tests, and latent growth curve modeling adjusting for clinical status (normal, amnestic mild cognitive impairment, or AD) to summarize initial performance and change over three years. Relative to individuals with two very long TOMM40 alleles, APOE ɛ3 homozygotes with one or two short alleles showed lower baseline cognitive performance regardless of clinical status. The number of short or very long TOMM40 alleles was not associated with longitudinal cognitive changes. In APOE ɛ3 homozygotes from the University of Kansas Alzheimer’s Disease Center cohort, an association between TOMM40 ‘523 and cognition is consistent with the possibility that TOMM40 influences cognition independent of APOE.
INTRODUCTION
After Schellenberg et al. [1] and Pericak-Vance et al. [2] established connections between Alzheimer’s disease (AD) risk and chromosome 19, Corder et al. [3] and Strittmatter et al. [4] concluded variants in the APOE gene, located at 19q13.32, were responsible. Specifically, the APOE ɛ2, ɛ3, and ɛ4 alleles were shown to respectively confer reduced, neutral, and increased late-onset AD risk. Subsequent studies generated or supported hypotheses that might explain the impact of APOE on AD, but mechanistic consensus remains elusive.
In 2010, Roses et al. [5] reported an association between AD risk and variation in a gene adjacent to APOE, TOMM40. The variant of interest is a variable poly-T repeat in intron 6 that defines rs10524523 (‘523). Roses et al. used the ’523 poly-T repeat length to designate three general alleles: a “short” allele of <19 poly-T’s, a “long” allele of 20–29 poly-T’s, and a “very long” allele of >30 poly-T’s. TOMM40 alleles form haplotypes with APOE, and in Caucasians the TOMM40 long allele forms a haplotype with APOE ɛ4. Therefore similar to APOE ɛ4, a strong statistical association between TOMM40 long and increased AD risk clearly exists.
Because of the tight linkage disequilibrium that occurs between TOMM40 long and APOE ɛ4 alleles in Caucasian populations, based purely on association it is difficult to conclude the extent to which each individual gene contributes to AD risk. Many investigators favor the view that APOE is solely responsible. Others speculate that TOMM40, which encodes a protein critical to mitochondrial function, the translocase of the outer mitochondrial membrane 40 kD protein, might independently affect AD risk through effects on mitochondrial function [6].
Consistent with this latter view, recent studies report the TOMM40 short and very long alleles variably influence AD risk, cognitive test performance, and changes in brain volumes. However, findings are not uniform and suggest published relationships may be context and methods-dependent. For example, two studies report contrasting findings among individuals homozygous for both TOMM40 short and APOE ɛ3 [7, 8]. A longitudinal study by Yu et al. reported that among cognitively normal older adults, TOMM40 short homozygotes declined faster than other TOMM40 combinations, while a cross-sectional study by Laczo et al. reported that TOMM40 short homozygotes with amnestic mild cognitive impairment showed better cognitive performance and larger brain volumes.
In the present study, we tested whether TOMM40 ‘523 short and very long alleles were associated with cross sectional cognitive test performance and longitudinal stability in members of the University of Kansas Alzheimer’s Disease Center (KUADC) clinical cohort. We hypothesized that short and very long alleles would be differentially associated with cognitive performance and decline. Due to the highly mixed findings of previous studies, we did not have a basis for hypothesizing a direction of influence. Cognitively normal, amnestic mild cognitive impairment (aMCI), and AD participants were included in our analysis. To minimize the confounding effects of the APOE ɛ4-TOMM40 long linkage disequilibrium, we limited the analysis to APOE ɛ3 homozygotes.
METHODS
Study design
The Uniform Data Set (UDS) was created in 2005 to collect standard clinical data on subjects from the National Institute on Aging (NIA)-supported Alzheimer’s Disease Centers (ADCs). The UDS is administered to ADC clinical core participants on an approximately annual basis.
The KUADC collects longitudinal data on a clinical cohort that includes over 400 individuals. The cohort includes participants with cognitive impairment as well as normal cognition. Cognitively normal individuals are included at age 60 and older, while individuals with AD are included at any age. It qualifies as a sample of convenience, with members recruited from the community via online, radio, and television advertisements; representation at community events; referrals from community partners; and word of mouth. The present study is a three-year longitudinal analysis of a subset of data collected from this clinical cohort. The first time point represents baseline and each additional time point represents follow up visits at approximately one-year intervals.
Consent
Entry into the cohort requires written consent from each participant, as well as written consent from a study partner. The consent forms and process were approved by the University of Kansas Medical Center’s institutional review board and was conducted in accord with the Helsinki Declaration of 1975. The consent form and process specify that cognitive and genetic data obtained from each participant will be included in studies pertinent to AD and brain aging.
Participants and participant selection
Participants and their study partners/informants annually undergo a Clinical Dementia Rating (CDR) interview. The participants complete the UDS evaluation defined by the NIA ADC network, as well as additional cognitive tests (i.e., letter number sequencing, free and cued selective reminding test, Stroop test). At the time of cohort entry, APOE genotypes are determined, and TOMM40 ‘523 genotypes are identified through PCR with subsequent capillary gel electrophoresis. Poly-T lengths are used to classify each TOMM40 allele according to the short, long, and very long convention of Roses et al. [5, 9].
Initial and subsequent annual CDR and neuropsychological test scores are reviewed at a weekly consensus diagnostic conference that includes clinicians, a neuropsychologist, psychometricians, and other staff who participate in the evaluation process. Participants with a CDR score of 0 and no demonstrable cognitive deficits are classified as cognitively normal. Demonstrable cognitive deficits are defined as scores less than 1.5 standard deviations below the mean on two or more tests based on data from the National Alzheimer’s Coordinating Center. Participants with a CDR score of 0.5 but no loss of functional independence, as well as demonstrable cognitive test deficits, receive a designation of MCI. MCI participants with deficits in cognitive tests that focus on the memory domain qualify for aMCI. Participants with a CDR score of 0.5 or higher, loss of functional independence, cognitive test deficits, a primary etiology that is felt to include AD, and no likely explanation for their dementia other than AD are classified as having AD. Some KUADC clinical cohort participants do not fall into any of these diagnostic categories; for this study, only cognitively normal, aMCI, and AD participants were considered.
Participants categorized as having normal cognition, aMCI, or AD were included if they were assigned to those respective groups at each independent assessment for all evaluation waves or followed an expected progression from normal to aMCI, aMCI to AD, or normal to AD. We excluded cases in which a diagnosis of normal cognition was reported subsequent to a diagnosis of aMCI or AD, or cases in which individuals were diagnosed with another form of dementia as the primary etiology at another occasion. The designated primary etiology for all of the aMCI participants was “MCI due to AD,” and aMCI participants were included only if all further assignments were to aMCI or if their group assignment changed to AD. Finally, we excluded individuals who possessed an APOE ɛ4 or APOE ɛ2 allele.
Genotyping procedures
APOE genotyping was performed by the National Cell Repository for Alzheimer’s Disease (NCRAD), with independent verification of selected samples by the KUADC Mitochondrial Genomics and Metabolism Core using a previously described allelic discrimination assay [10]. TOMM40 ‘523 genotyping was performed by Polymorphic DNA Technologies (PDT), with independent verification of selected samples by the KUADC Mitochondrial Genomics and Metabolism Core using a previously reported procedure [10]. Poly-T length reproducibility between the PDT and KUADC measurements was uniformly within 1 T.
Cognitive tests
A standardized cognitive test battery was administered by a trained psychometrician, and included Logical Memory IA and IIA [11], selective reminding test [12], Digit Span forward and backward, letter number sequencing [13], Trail Making B [14], Category Fluency [15], the Digit Symbol Substitution Test [13], and the Stroop Test, Interference Condition [16]. For tests that changed from the UDS version 2.0 to the UDS version 3.0, we used the previously published “cross walk” scores that estimate the equivalent scores across the two versions of the test (i.e., Craft Immediate and Delayed Recall replaced Logical Memory IA and IIA, and number span forward and backward replaced digit span forward and backward) [17].
Covariates and statistical analysis
Age, sex, and years of education were treated as covariates. All cognitive tests were standardized using Z-scores relative to the mean of the cognitively normal group at baseline. Thus, scores can be interpreted as a difference from baseline in cognitively normal persons. We report findings from four waves of assessment, each one year apart.
We used Mplus to combine cognitive test scores into domain specific factor scores using Confirmatory Factor Analysis. Specific tests were organized by whether they primarily address attention, verbal memory, or executive function. Tests of attention included digits forward, digits backward, and letter-number sequencing. Tests of verbal memory included immediate and delayed logical memory, as well as the sum of three selective reminding test trials. Tests of executive function assessed set maintenance and shifting and included category fluency (sum of animal and vegetable categories), Stroop color word interference, trailmaking test B, and the digit symbol substitution test. We included the combined cognitive test scores as outcomes in subsequent models.
We estimated latent growth curve models to describe the intercept and change in performance in these three cognitive domains over four waves of data collection. Latent growth curve models are advantageous because they allow estimation of error variance to be included in the model. We specifically considered the effect of TOMM40 ‘523 status on the cognitive parameters. The full model estimated the association of TOMM40 with the intercept and slope of cognitive decline adjusting for covariates (age, sex, education, clinical status). We also tested an interaction term (clinical status×TOMM40 allele status) that would allow us to investigate whether TOMM40 allele status influenced cognitive performance differently in individuals with normal cognition compared to those with aMCI or AD. We estimated change in R2 between models with and without TOMM40 allele status as a measure of effect size.
Missing data were accounted for using a full information maximum likelihood algorithm. To evaluate model fit we used Root Mean Squared Error of Approximation (RMSEA), a measure of the discrepancy between predicted and observed model values. Values closer to 0 indicate better fit (preferred values are <0.09). We report a comparative fit index (CFI) that estimates the relative fit of a model compared to an alternative model, in which a CFI >0.90 indicates good fit. Typically, these multiple fit indices are considered together, as opposed to relying on any one indicator by itself. We used structural equation modeling to estimate all the pathways simultaneously, which allowed us to avoid multiple testing-induced inflation of type 1 error. Monte Carlo simulation estimated that we have >0.80 power to detect all significant estimates in the model.
RESULTS
We analyzed data from 173 KUADC clinical cohort participants. Because we excluded APOE ɛ4 carriers, we also filtered out all but three instances of the TOMM40 long allele. Among the TOMM40 alleles recorded, the various combinations of TOMM40 alleles were observed in the following frequencies: 29.2% short/short, <1% short/long, 47.6% short/very long, 0% long/long, 1.2% long/very long, and 21.4% very long/very long. We excluded the three individuals with a TOMM40 long allele from subsequent analyses for a final sample size of 170 at baseline. Due to attrition, sample sizes were smaller at each subsequent wave of follow up (Wave 2 = 167, Wave 3 = 147, Wave 4 = 104).
Table 1 shows baseline participant characteristics. A three-factor model of cognitive domains fit the data at all four waves (Wave 1 χ2 (df) = 131.60 (30), RMSEA = 0.089, CFI = 0.97; Wave 2 χ2 (df) = 98.47 (30), RMSEA = 0.074, CFI = 0.98; Wave 3 χ2 (df) = 81.34 (30), RMSEA = 0.071, CFI = 0.98; Wave 4 χ2 (df) = 74.20 (30), RMSEA = 0.081, CFI = 0.98). Table 2 shows the loadings of each cognitive test onto the factors at wave 1. Variables associated with baseline cognitive performance (intercept) and change across waves (slope) were estimated using latent growth curve modeling. Table 3 shows the results of these models, which are adjusted for age, sex, education, and clinical status.
Baseline participant characteristics
*Difference in number of males and females by clinical status was statistically significant p < 0.001.
Confirmatory factor analysis of cognitive domains at baseline
Variables associated with intercept and slope for attention, verbal memory, and executive function
For all three cognitive domains, having a higher number of short TOMM40 alleles was associated with lower baseline performance (intercept) regardless of clinical status. This effect was statistically significant for attention and executive function, and was a non-significant trend in the expected direction for verbal memory. Having a greater number of short alleles was not significantly associated with rates of cognitive decline over the longitudinal follow up. The pattern of declining performance by clinical status and TOMM40 length is represented visually in Fig. 1 in a common format for presenting the results of growth curve models. For simplicity of presentation, only homozygotes (short/short or very long/very long) are depicted in the figure. The lines represent the mean performance across waves by clinical status, TOMM40 genotype, adjusting for covariates in the model. The mean baseline cognitive scores split by TOMM40 allele combinations are depicted in Fig. 2. As planned, we tested an interaction effect (clinical status×TOMM40 allele status) that allowed us to investigate whether TOMM40 allele status influenced cognitive performance differently in individuals with normal cognition compared to those with aMCI or AD. We did not find this interaction effect to be statistically significant in any of the models (attention intercept β= –0.089, p = 0.365; slope β= 0.371, p = 0.219; verbal memory intercept β= –0.079, p = 0.274 slope β= 0.205, p = 0.264; executive function intercept β= –0.130, p = 0.111 slope β= 0.267, p = 0.133). Finding no significant differences by dementia status, we present the means according only to TOMM40 genotype in Fig. 2 for simplicity.
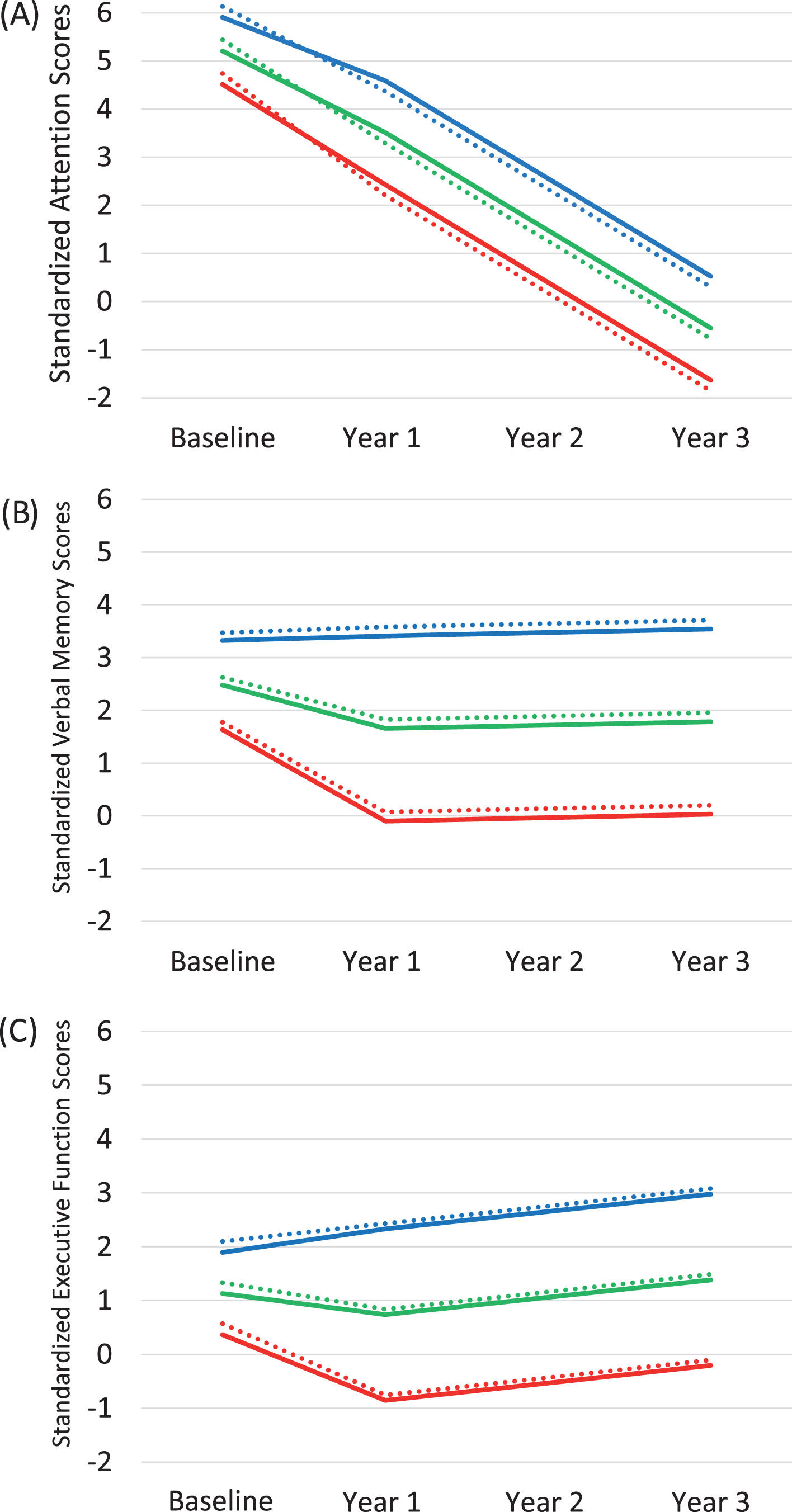
Longitudinal cognitive change by clinical status and TOMM40 length. A) Attention. B) Verbal memory. C) Executive function. To allow comparison across tests and participants, cognitive scores on the Y-axis are standardized to the baseline performance of the cognitively normal participants. Patterns of change for individuals with mean age and education representing each clinical status category and the short/short and very long/very long TOMM40 categories are illustrated. Solid lines represent short/short homozygotes, while dashed lines represent very long/very long homozygotes. Blue indicates cognitively normal subjects, green indicates aMCI subjects, and red indicates AD subjects. Though visually, the lines appear very close together, the difference at the intercept between the dotted lines and solid lines are statistically significantly different for (A) attention and (C) executive function at p < 0.05. The difference at the intercept between dotted lines and solid lines are not statistically significant for (B) verbal memory p = 0.09.
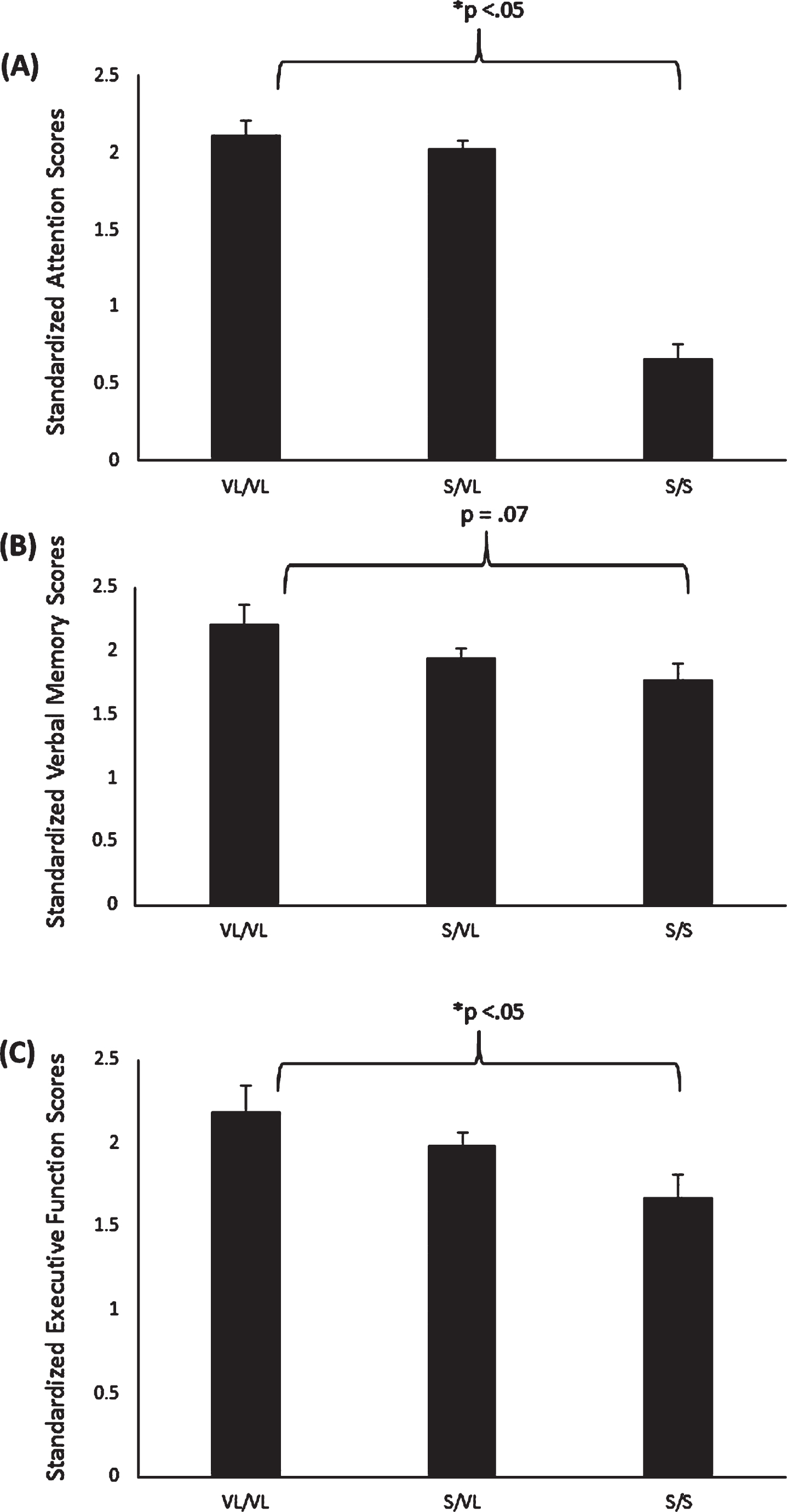
Mean standardized baseline cognitive scores by number of TOMM40 genotype. A) Mean baseline attention. B) Mean baseline executive function. C) Mean baseline verbal memory. Error bars indicate standard deviation.
Table 4 shows the fit indices for the growth models and the exact R2 values for each model. Across the four waves, the amount of variance explained by TOMM40 genotype in the model ranged from R2 = 0.000 to 0.008 for attention, R2 = 0.007 to 0.021 for verbal memory, and R2 = 0.003 to 0.011 for executive function. We tested the change in variance explained with and without the TOMM40 allele parameter to estimate the degree of variance explained only by TOMM40 alleles (indicated in the table by ΔR2). The proportion of variance explained was small, but consistently around 1–2%.
Growth curve fit indices and change in R2
DISCUSSION
Overall, KUADC clinical cohort participants with two TOMM40 short alleles showed lower baseline performance compared to those with two very long alleles, regardless of clinical status. TOMM40 allele status was not associated with the rate of longitudinal cognitive decline over a three-year follow-up period. Since individuals contributing data to this analysis were APOE ɛ3 homozygotes, the previously recognized linkage disequilibrium between TOMM40 long and APOE ɛ4 does not explain the observed relationship. Narrowly interpreted, these findings from APOE ɛ3 homozygotes suggest TOMM40 ‘523-defined alleles, to at least some extent, influence cognitive performance regardless of clinical status.
Three previous longitudinal studies also considered whether TOMM40 ‘523 genotypes affect cognition. These studies used different methodological approaches, and reached different conclusions [7, 19]. Our study and that of Yu et al. [7] evaluated older adults (mean age >70), and found TOMM40 short homozygotes had poorer cognitive function, and in the Yu et al. study, steeper rates of cognitive decline. Payton et al. [19] on the other hand, included a broader age range and concluded that TOMM40 short carriers had poorer baseline performance, but very long homozygotes had faster rates of decline. The Caselli et al. [18] study observed detrimental effects of TOMM40 very long over longitudinal follow up (i.e., a diminished test-retest effect) only in subjects younger than 60, and not in subjects over 60. These age differential results are consistent with the resource modulation hypothesis, which suggests the effects of genetic polymorphisms on cognition vary with advancing age, and change as cognitive performance declines from optimal levels [20]. In essence, this hypothesis proposes that genes may have a different impact on the relationship between age-related loss of brain resources and cognitive performance as neural reserve depletion occurs.
In contrast to the two longitudinal studies that focused on older adults, results of previous cross-sectional studies of TOMM40 ‘523 and cognitive performance mostly report TOMM40 short/short homozygotes have better cognitive outcomes than those with longer repeat lengths [8, 21–23]. It is important to note, however, that cognitive abilities observed at a single time point may not predict rate of change over time, and that factors which influence performance at a single time point may differ from those that affect longitudinal course.
Our study and the aforementioned studies also differ in terms of duration. We followed subjects over a three-year period, while the longest longitudinal study spanned 14 years. Cognitive tests and the cognitive domains examined also varied across studies. As some cognitive domains tend to remain more stable over time than others, differences in test batteries may influence the ability to detect and track cognitive decline-related changes. The test battery we used was designed to follow decline in persons with dementia, and is less useful for monitoring age-related cognitive change in normal individuals. Caselli et al. [18] used only a single verbal memory test, while the other three studies used composite scores to summarize tests from a wider array of cognitive domains. The two studies that focused on older adults, the Yu et al. study and the present study, both reported detrimental effects of short TOMM40 alleles on memory performance. Payton et al. [19] reported statistically significant findings only in vocabulary, which tends to be quite stable across time, even in older age. Future research would benefit from harmonization of the cognitive domains studied, and more consistent methods of combining multiple tests scores to enable better comparisons.
Each of the previous longitudinal studies used a different approach to the inclusion or exclusion of individuals with cognitive impairment. In the present study, we included individuals who remained cognitively normal, converted from cognitively normal to cognitively impaired, or who remained cognitively impaired. Yu et al. [7] included only participants that were not impaired at baseline, but who may have converted to impairment over the longitudinal follow up. These differences may affect the trajectory of cognitive declines observed and their relationship to TOMM40 ‘523. Caselli et al. [18] included only individuals who were cognitively normal at all time points, whereas dementia status was not clearly determined in Payton et al. [19]. These differences in study design may have important implications for interpretation of the effect of TOMM40 alleles on cognitive performance and decline. Few studies explicitly limit analyses to APOE ɛ3 homozygotes to differentiate the independent effects of TOMM40.
The KUADC clinical cohort is predominantly Caucasian. Because Caucasians show tight linkage disequilibrium between APOE ɛ4 and TOMM40 long alleles, excluding APOE ɛ4 carriers from the analysis also eliminated most TOMM40 long carriers and we excluded the remainder from subsequent analyses. Our results do not directly address the question of whether TOMM40 long alleles influence AD risk independently of APOE ɛ4. From a strict epidemiologic and statistical perspective, at least in Caucasians, the relative contributions of APOE ɛ4 and TOMM40 long alleles to AD risk remains confounded.
Epidemiologic studies of African-descended populations may help resolve this confounding, as some of these populations do not show tight linkage disequilibrium between the APOE ɛ4 and TOMM40 long alleles. Indeed, a second Yu et al. study [24] recently reported that in an African American cohort, slightly less than half of the identified APOE ɛ4 alleles presented within the context of an inferred APOE ɛ4-TOMM40 long haplotype. APOE ɛ3-TOMM40 long haplotypes were rarely observed. The apparent APOE-TOMM40 haplotype status had a remarkable effect on incident dementia. African Americans who had neither an APOE ɛ4 allele nor a TOMM40 long allele had an 8.4% risk of incident dementia. Those with 1 or 2 APOE ɛ4 alleles, but no TOMM40 long alleles had a 10.8% risk of incident dementia. Those with 1 or 2 APOE ɛ4 alleles and at least 1 TOMM40 long haplotype had a 17.7% risk of incident dementia. We note that these results fail to take into account the dose-response relationship with 1 versus 2 APOE ɛ4 alleles, thus gene dosage could provide a potential explanation for those findings.
In general, the associations between cognitive performance and TOMM40 short alleles that were seen in our study suggest the TOMM40 gene is relevant to cognition independent of the effect of APOE. We cannot conclude the extent to which TOMM40 contributes, although the impact of a TOMM40 short versus TOMM40 very long allele appears quite limited. When it comes to relationships between single genes and behavior, though, small effect sizes are generally expected. Although APOE has a larger effect size by comparison, genes with effect sizes that are readily detectable by linkage are rarer [25]. This is sometimes called “the fourth law of behavior genetics” which describes that complex human behaviors are associated with many genetic variants, each of which accounts for a very small percentage of variability in the behavior [26]. Allelic association can detect genes that account for as little as 1% of the variance in a trait and most single nucleotide polymorphisms (SNPs) function in concert with other genes within qualitative trait loci (QTLs) which may interact in complex ways. Furthermore, behavioral traits are difficult to measure with accuracy compared to more objectively measured outcomes [25, 27].
The mechanistic connection between APOE and AD remains unresolved, partly because multiple viable mechanistic explanations currently exist. Potential scenarios include effects on beta amyloid trafficking, oxidative stress, lipid homeostasis, neuron plasticity, and mitochondrial function [28]. It is also possible to propose mechanisms through which TOMM40 might influence AD risk [6]. As TOMM40 encodes a critical mitochondrial protein, a reasonable hypothesis would be that different ‘523 poly-T lengths ultimately affect mitochondrial function. While the effects of the different TOMM40 ‘523 alleles on mitochondrial function are currently unknown, it does appear that ‘523 poly-T length does influence TOMM40 transcription and, ultimately, protein levels [9, 29].
Our study has limitations. Studies of genetic markers without replication lack empirical rigor. Given the paucity of available comprehensive comparison datasets in which to complete a replication, we can only rely on our comparison of our results to others in the literature. Our longitudinal follow-up period of three years is relatively short, and certainly shorter than the longitudinal follow-up periods in the other reports. A longer duration might have allowed us to detect the steeper declines suggested by Yu et al. Our sample size was considerably smaller than that of Yu et al. [7], and thus we had less statistical power to detect the small effects that allelic contributions typically have on complex behaviors. Unique contributions of the present study include exclusion of APOE ɛ4 alleles to facilitate decoupling of the APOE ɛ4-TOMM40 ‘523 linkage disequilibrium, a range of clinical cognitive status (that included cognitively normal, aMCI, and AD subjects), and multiple cognitive tests summarized into distinct cognitive domains.
In the decades since relationships between AD and APOE isoforms were first recognized, genome wide association studies have implicated numerous specific genes with AD risk. The odds ratios for these genes are frequently quite small, although the impact of these discoveries is amplified by the insight they provide into the functional biology that determines AD. Partly because of genetic association, we now recognize roles for endosome function, immune function, cholesterol metabolism, and broader lipid metabolism [30]. Our findings from this study additionally support the increasingly accepted view that mitochondrial function also contributes to AD risk [31, 32].
Footnotes
ACKNOWLEDGMENTS
This work was supported by National Institute on Aging funding to the University of Kansas Alzheimer’s Disease Center (P30AG035982). The University of Kansas Alzheimer’s Disease Center Database and Statistics Management Core developed data tools that facilitated analyses performed in this study.