
Abstract
The association of Apolipoprotein E (APOE) with late-onset Alzheimer’s disease (LOAD) and cognitive endophenotypes of aging has been widely investigated. There is increasing interest in evaluating the association of other LOAD risk loci with cognitive performance and decline. The results of these studies have been inconsistent and inconclusive. We conducted a systematic review of studies investigating the association of non-APOE LOAD risk loci with cognitive performance in older adults. Studies published from January 2009 to April 2018 were identified through a PubMed database search using keywords and by scanning reference lists. Studies were included if they were either cross-sectional or longitudinal in design, included at least one genome-wide significant LOAD risk loci or a genetic risk score, and had one objective measure of cognition. Quality assessment of the studies was conducted using the quality of genetic studies (Q-Genie) tool. Of 2,466 studies reviewed, 49 met inclusion criteria. Fifteen percent of the associations between non-APOE LOAD risk loci and cognition were significant. However, these associations were not replicated across studies, and the majority were rendered non-significant when adjusting for multiple testing. One-third of the studies included genetic risk scores, and these were typically significant only when APOE was included. The findings of this systematic review do not support a consistent association between individual non-APOE LOAD risk and cognitive performance or decline. However, evidence suggests that aggregate LOAD genetic risk exerts deleterious effects on decline in episodic memory and global cognition.
INTRODUCTION
Cognitive performance generally declines with age, however, the patterns are characterized by 1) differences across cognitive domains and 2) substantial individual variation in level and trajectory [1, 2]. Performance on measures of episodic memory, executive function, reasoning, and processing speed may begin to decline in early adulthood whereas gradual improvement in some verbal and knowledge abilities may continue to the sixth or seventh decade of life [3]. Variation in individual trajectories reflects life-long differences in demographic, lifestyle, medical, environmental, neurobiological, and genetic factors [4].
Cognitive decline is a multifactorial process that is likely promoted by the gradual accumulation of neuropathology associated with various chronic conditions of aging [5–7] and in particular late-onset Alzheimer’s disease (LOAD) [8]. The accumulation of amyloid-β (Aβ) and neurofibrillary tangles (NFT) begins decades prior to the onset of the clinical symptoms of LOAD [9–12]. In dementia-free individuals a higher burden of LOAD pathology is on average associated with reduced cognitive performance and faster rates of cognitive decline [13–15]. As such, age-related cognitive decline may be mediated by the co-occurrence of Aβ, NFT, and other neuropathologies [16–18].
Genetic factors play an important role in the development of LOAD, accounting for 53% of the total phenotypic variance [19]. The Apolipoprotein E (APOE) epsilon (*ɛ4) allele was the first common genetic variant associated with LOAD [20], with recent genome-wide association studies (GWAS) identifying a further 42 loci associated with LOAD (Supplementary Table 1). GWAS performed separately by four LOAD genetic consortia initially identified 11 loci (ABCA7, BIN1, CD2AP, CD33, CLU, CR1, EPHA1, MS4A4A, MS4A4E, MS4A6A, and PICALM) [21–25]. A further 12 loci (HLA-DRB5, PTK2B, SORL1, SLC24A4-RIN3, INPP5D, MEF2 C, NME8, ZCWPW1, CELF1, FERMT2, and CASS4) were identified in a meta-analysis by the International Genomics of Alzheimer’s Project (IGAP) [26]. A meta-analysis of IGAP and a proxy GWAS case-control study of self-reported family history of parental Alzheimer’s dementia in 114 564 (14 482 proxy-cases & 100 082 proxy-controls) individuals from the UK Biobank identified a further 4 loci (HBEGF, ECHDC3, SCIMP, and SPPL2A) [27].
A trio of recent GWAS have identified a further 16 loci. A second meta-analysis of IGAP with an expanded UK Biobank dataset (n = 314 278) identified three loci (ADAM10, KAT8, and ACE) [28]. A meta-analysis of UK Biobank proxy case-control status (n = 376,113), the personality genomics consortium Alzheimer’s disease working group of the Psychiatric Genomics Consortium (PGC-ALZ, n = 17,477), IGAP (n = 54,162), and the Alzheimer’s Disease Sequencing Project (ADSP, n = 7,506) identified 8 loci (ADAMTS4, HESX1, CLNK, CNTAP2, APH1B, ABI3, ALPK2, and ACO74212.3) [29]. Finally, an expanded IGAP analysis (n = 94,437) identified five loci (OARD1, TREM2, IQCK, WWOX, and ADAMTS1) [30]. TREM2 and ABI3, however, were identified as AD associated loci in an earlier rare variant analysis [31].
There is increasing interest in evaluating the role of LOAD genetic risk variants with cognitive decline. First, the shared cognitive and neuroanatomical characteristics of normal cognitive aging and the early stages of LOAD may be mediated by shared genetic mechanisms. The presence of individual LOAD-associated risk loci may lead to diminished overall cognitive function, in the absence of cognitive impairment or dementia, mediated by the gradual accumulation of LOAD pathology [13, 14]. Second, cognitive decline prior to dementia represents an important endophenotype for LOAD. Cognitive domain-specific variance reflects localized regional brain structures/networks and the connectivity of those networks. Therefore, the differential association of individual loci with specific cognitive domains may reflect associations with particular neuroanatomical structures that influence LOAD onset and progression.
Initial support for the association of LOAD risk loci with cognitive performance was obtained from studies assessing the association of APOE with cognition, where the APOE*ɛ4 allele was associated with specific deleterious effects on episodic memory, executive functioning, perceptual speed, and global cognitive ability [32, 33]. Further studies examining the association of other LOAD risk loci with cognitive function have been inconsistent and inconclusive. The aim of this systematic review is to evaluate the evidence of the association of non-APOE LOAD risk loci with cognitive performance and decline, within the context of both cognitive aging and a LOAD cognitive endophenotype. We provide a narrative synthesis rather than focusing on the relatively few studies that would be amenable to meta-analysis due to the heterogeneity in methodologies between studies.
METHODS
Registration of protocol and reporting
The protocol for the review was registered with the International Prospective Register of Systematic Reviews (PROSPERO CRD42017075685) [34] and the review is reported in accordance with the PRISMA checklist (see Supplementary Material).
Search strategy
A PubMed database search (see Supplementary Material) included papers published between January 2009 (the publication year of the first GWAS to identify non-APOE genome-wide significant SNPs for LOAD) and April 2018 (inclusive). Articles were restricted to human studies published in English. Reference lists of all articles selected for data extraction were screened for additional articles.
Inclusion and exclusion criteria
Studies were included in the review if they met the following inclusion criteria: 1) included genetic data from non-APOE genome-wide significant risk loci for LOAD (ABCA7, BIN1, CD2AP, CD33, CLU, CR1, EPHA1, MS4A4A, MS4A4E, MS4A6A, PICALM, HLA-DRB, PTK2B, SORL1, SLC24A4, RIN3, INPP5D, MEF2 C, NME8, ZCWPW1, CELF1, FERMT2, CASS4, HBEGF, ECHDC3, SPPL2A, and SCIMP) or a LOAD genetic risk score (GRS); 2) included at least one test measuring cognitive performance; 3) the publication was in English; 4) it was either cross-sectional or longitudinal. Articles were excluded if they were: 1) case only studies, case reports or review articles; 2) animal studies; or 3) conducted in a clinical population.
Abstract screening and article selection
Article citations and abstracts were imported into Covidence [35], rated against the selection criteria, and nominated independently for inclusion in full-text screening by SJA and GPM. Subsequently, full-text articles were assessed for inclusion in the final review. When the two reviewers differed, the article was discussed until a consensus was reached.
Data extraction
For articles included in the systematic review, the following variables were extracted: 1) study design (i.e., longitudinal or cross-sectional; candidate SNPs, gene-based or GWAS analysis; statistical test); 2) sample characteristics (i.e., sample size, age, education, gender, ethnicity/population, follow-up, and cognitive status); 3) genetic variants examined; 4) cognitive tests examined; and 5) reported associations (i.e., non-significant result, positive association, negative association). Given the heterogeneity in the measures with which the reviewed articles assessed cognitive performance, all the cognitive tests were coded within conventional cognitive domains [33] (Supplementary Table 2). These domains are based on the typical taxonomy found in the neuropsychological literature and were used in pervious previous meta-analyses on the effect of APOE on cognitive performance [33, 36]. Cognitive domains included: attention (AT), episodic memory (EM), executive function (EF), global cognition (GC), perceptual speed (PS), working memory (WM), verbal ability (VA), and visuospatial skill (VS). Two general cognition clusters were included: fluid cognition (Gf) and crystallized cognition (Gc). Study quality was evaluated using the 11-item Quality of Genetic Studies (Q-Genie) Tool [37] (Supplementary Material). Inter-rater reliability was assessed by calculating a two-way consistency average-measures interclass correlation coefficient (ICC).
Novel AD loci
The initial screen did not include the 16 novel loci identified by Marioni et al. [28], Janssen et al. [29], and Kunkle et al. [30] (ADAM10, KAT8, ACE, ADAMTS4, HESX1, CLNK, CNTAP2, APH1B, ABI3, ALPK2, ACO74212.3, OARD1, TREM2, IQCK, WWOX, and ADAMTS1) as these studies were published after the database search and article screening were conducted. As such, for the loci reported in these studies we limited our search to articles citing either the BioRxiv pre-print article or the published article as of March 2019. Additionally, where GWAS summary statistics were available for cognitive phenotypes, we extracted the reported associations for these loci.
RESULTS
Systematic literature search
The PubMed search identified 2,446 references and follow-up screening of reference lists identified two additional articles. 2,395 references were removed based on the inclusion/exclusion criteria. Seventy-one full-text articles were reviewed, 21 were excluded as follows: 1) fifteen due to selected AD risk loci not reported, 2) one was an updated analysis of a previous study, 3) two because summary statistics were not made publicly available, and 4) three as the study was conducted in adolescents. Forty-nine articles were included in the systematic review (Supplementary Figure 1).
For each study we report study characteristics (Table 1), study design (Table 2), individual cognitive tests and the respective cognitive domains tested (Supplementary Table 2), and individual SNPs genotyped (Supplementary Table 3). Of the forty-nine studies, 23 employed a cross-sectional design and 26 a longitudinal design. 29 selected SNPs based on a candidate gene approach, 7 employed gene-based analyses, 6 reported AD risk loci as a secondary outcome in GWAS, and 17 included a GRS, with 8 studies only using a GRS. Episodic memory (n = 31, 63.27%) and global cognition (n = 23, 46.94%) were the most commonly assessed cognitive measures.
Study Characteristics
*Median (range); †Median (IQR); UTAI, Up to and Including.
Description of the Methods used for each study
The overall average quality rating was ‘good’, with four studies obtaining a ‘moderate’ score. The distribution and mean rating for each item and the average score per study are presented in Supplementary Figures 2 and 3. The ICC was in the excellent range (ICC = 0.88 95% CI: 0.79 - 0.93), indicating that reviewers had a high degree of agreement in the overall quality of the included studies.
Association of AD genetic risk loci with cognitive performance and change
In the following narrative, we report all gene-cognition associations that are statistically significant (p < 0.05) (Figs. 1 and 3). However, it should be noted that the majority (84.3%) of the reported associations were non-significant (Supplementary Table 4). The number of studies investigating the association of each LOAD loci with cognitive function and the number of studies reporting at least one significant association for each gene-cognitive domain combination is reported in Supplementary Table 4. Across cognitive domains/clusters, GC had the highest proportion of reported significant associations (30.2%, 77/255) followed by VS (30%, 3/10), VA (14.29%, 16/112), EM (14.29%, 32/224), AT (13.33%, 6/45), EF (11.86%, 14/118), PS (11.79%, 23/195), Gf (7.46%, 5/67), WM (4.05%, 3/74), and Gc (0%, 0/38). The largest studies to report an association between the AD risk loci and GC, were two GWAS meta-analyses inclusive of the UK Biobank (n = 269,867 and 300,486) [38, 39] and a multi-trait analysis of intelligence and educational attainment (n = 248,482) [40]. Davies et al. [39] found 18 loci associated with GC (MEF2C, HBEGF, SPPL2A, IQCK, ABI3, FERMT2, CELF1, CR1, CNTNAP2, SLC24A4, AC074212.3, CLU, ABCA7, ADAM10, PTK2B, CD2AP, CLNK, and WWOX), of which only MEF2C, HBEGF, and SPPL2A were genome-wide significant. Savage et al. found 11 loci to be associated with GC (MEF2C, HBEGF, SPPL2A, CR1, SLC24A4, OARD1, CNTNAP2, WWOX, ZCWPW1, CELF1, and ABCA7), of which MEF2C, HBEGF, and SPPL2A were also genome-wide significant [38]. Finally, Hill et al. [40] identified 13 loci associated with global cognition (MEF2C, HBEGF, CELF1, ZCWPW1, SPPL2A, WWOX, HLA-DRB1, SLC24A4, ADAMTS4, ALPK2, ACE, SORL1, and PICALM), of which MEF2C, HBEGF, CELF1, and ZCWPW1 were genome wide significant.
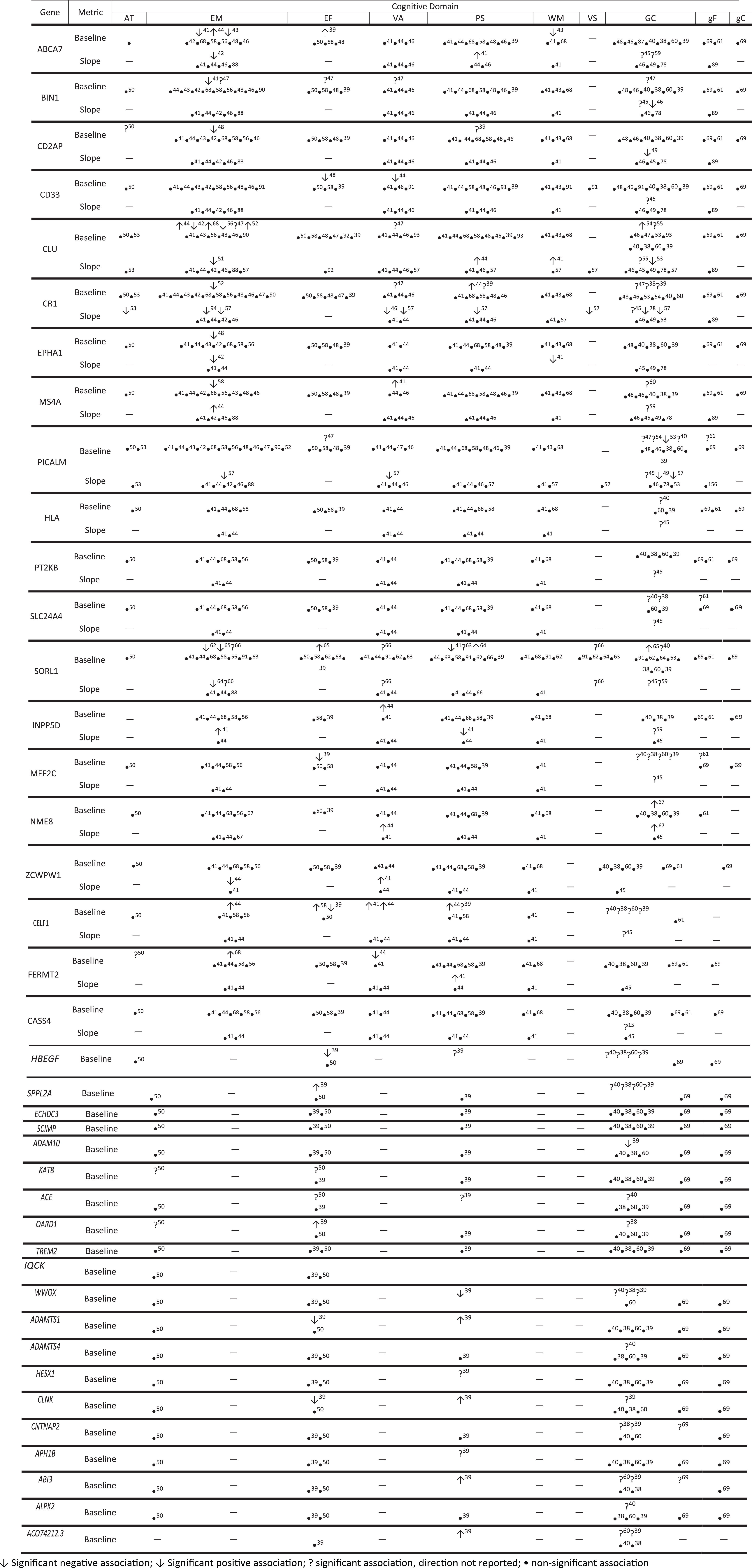
Reported gene – cognitive domain associations.
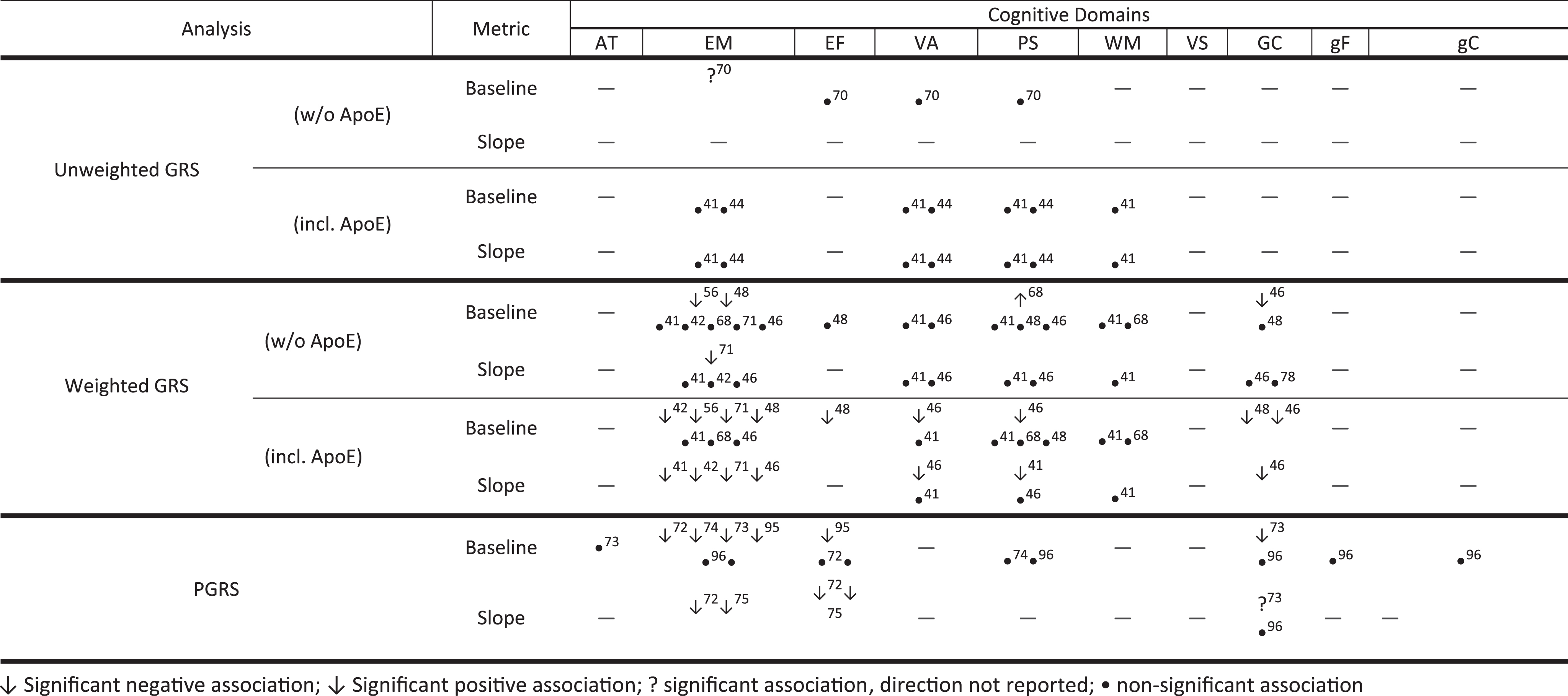
Reported genetic risk scores – cognitive domain associations.
ABCA7
rs3764650(G) was associated with worse baseline performance and slower decline in EM [41]. In a second study, rs3764650(C) was associated with faster decline in EM in cognitively normal participants who converted to mild cognitive impairment (MCI)/Alzheimer’s disease (AD), but not in participants who remained cognitively normal [42]. Additionally, rs3752246(G) was associated with worse performance in EM and WM at baseline [43], whereas rs4147929(A) was associated with better baseline EM [44] and EF [39] performance. Change in GC was associated with rs115550680(G) in African-Americans and with the ABCA7 gene-region in a female only and a male only cohort [45].
BIN1
rs744373(G) was associated with worse baseline EM performance [41] and a faster rate of decline in global cognition [46]. In univariate (7 SNPs) and haplotype analyses (two 3-SNP windows), significant associations were observed for cognitive performance in EM, EF, VA, and GC [47]. The BIN1 gene region was associated with change in GC in females [45].
CD2AP
rs9349407(C) and rs9296559(G) were associated with worse EM performance and a faster rate of decline in GC respectively [48, 49]. The CD2AP gene region was also associated with performance in AT [50] and PS [39].
CD33
rs3865444(C) was associated with worse baseline performance in EF [48], and in African-Americans rs3865444(A) was associated with worse baseline performance in VA [44]. The CD33 gene region and rs3865444 were associated with change in GC in females [45].
CLU
rs11136000(C) was associated with faster decline in WM [41] and EM in participants who converted to MCI/AD, but not in participants who remained cognitively normal [51]. rs11136000(C) was also associated with better performance in EM in a combined cohort of case/controls, but not in non-demented subjects only [52]. In a follow-up study, rs11136000(G) was associated with worse baseline performance in EM [42]. rs11136000(T) minor allele was associated faster decline in GC [53]. Mengel-From et al. [54, 55] investigated the association of four separate SNPs in the CLU locus with cognitive function. They reported that rs11136000(T) was associated with better baseline GC, rs9331888(G) and rs9331908(T) were associated with slower decline and rs11136000(T) and rs1532278(T) were associated with faster decline [54, 55]. Bressler et al. [44] observed that rs9331896(C) was associated with better baseline performance in EM and a reduced rate of decline in PS. rs2279590(A) was associated with worse performance in EM [56] and two separate 3-SNP haplotypes were significantly associated with baseline performance in EM and VA [47].
CR1
rs3818361(T) was associated with faster decline in AT [53], while rs3818361(A) was associated with baseline performance in GC and faster decline in VA [47, 46]. Additionally, in African-Americans rs3818361(A) was associated with worse performance in EM in both a combined case/control cohort and non-demented control only subjects [52]. rs6656401(A) was associated with improved baseline performance in PS in African-American [44] and with faster decline in EM, semantic memory, PS, VS, and GC [47, 57]. Finally, a 3-SNP haplotype and 2-SNP haplotype was associated with VA and GC, respectively [47]. The CR1 gene region was associated with change in GC in females [45], PS [39], and GC [38].
EPHA1
rs11767557(C) and rs11767557(T) were associated with worse EM performance [48] and faster decline in WM, respectively [41]. Additionally, rs11767557(A) was associated with a faster rate of decline in EM in participants who converted to MCI/AD, but not in participants who remained cognitively normal [42].
MS4A
MS4A6A-rs983392(G) was associated with worse EM performance [58] and in African-Americans with change in GC [59]. MS4A4E-rs670139(T) was associated with better baseline WM [41] and slower decline in EM [44]. The MS4A4E and MS4A6A gene regions were associated with GC [60].
PICALM
rs3851179(A) and rs3851179(G) were associated with better baseline GC [54] and faster decline in GC respectively [49]. rs7110631(G) was associated with faster decline in EM, VA, and GC [57], while rs541458(C) was associated with an earlier age at midpoint in decline in a non-linear trajectory of GC [53]. In univariate analysis 4 SNPs (rs10501604, rs10792821, rs11234532, rs10501608) were associated with EF, while in haplotype analyses 12 3-SNP windows were associated with EF [47]. The PICALM gene region was associated with Gf performance [61] GC in a multi-trait analysis of intelligence and educational attainment [40], and with change in GC in males [45].
SORL1
rs3824968(A) was associated with worse EM performance at age 70, before and after adjusting for childhood IQ at age 11 [62]. In Chinese participants, rs2070045(T) was associated with PS performance [63] and rs1699102(T) was associated with faster decline in EM and PS [64]. rs11218343(T) was associated with worse PS at baseline [41]. In African-Americans, rs11218343(C) was associated with change in GC [59]. The SOLR1 gene region was associated with change in GC in males [45] and with GC in a multi-trait analysis of intelligence and educational attainment [40]. In a Dutch population-based study, rs668387(T), rs689021(A), and rs641120(T) were associated with worse EM performance, but better EM and GC performance [65]. A further three SNPs (rs3824968(T), rs2282649(T), rs1010159(C)) were associated with better performance in EF in the family based study [65]. In three Swedish based population cohorts, five SNPs (rs11600875, rs753780, rs7105365, rs11820794, rs2070045) were variously associated with performance in EM, VA, and VS [66].
Other loci
The HLA gene region was associated with change in GC in a female only and male only cohort [45]. The PTK2B gene region was associated with change in GC in males [45]. The SLC24A4 gene region was associated with Gf performance [61] GC in a multi-trait analysis of intelligence and educational attainment [40] and in a meta-analysis inclusive of the UKBB [38], and change in GC [45]. INPP5D-rs35349669(T) was associated with better baseline VA [44], slower decline in EM, and faster decline in PS [41]. In African-Americans, the INPP5D-rs4585024(A) minor allele was associated with change in GC [59]. MEF2C-rs190982(A) was associated with decreased EF performance in the UKBB, though it was non-significant in an earlier, smaller, analysis [39]. The MEF2C gene region was associated with GC in a multi-trait analysis of intelligence and educational attainment [40], GC in two large meta-analyses inclusive of the UK Biobank [38], Gf performance [61], and change in GC in males [45]. NME8-rs12155159(G) was associated with slower decline in VA [44] and NME8-rs2718058(G) was associated with worse baseline performance and faster decline in GC [67]. ZCWPW1-rs1476679(T) was associated with slower decline in PS [41], while in African-Americans ZCWPW1-rs1476679(C) was associated with faster decline in EM [44]. For CELF1, rs6485758(A) was associated with better baseline performance in EM, VA, and PS [44], while rs10838725(C) and rs7933019(C) were associated with better baseline EF performance [58] and a slower decline in EM [41], respectively. rs10838725(T) was associated with decreased EF performance [39]. The CELF1 gene region was associated with change in GC in females [45], GC in a multi-trait analysis of intelligence and educational attainment [40], GC in three large meta-analyses inclusive of the UK Biobank [38, 39], and with PS [39]. FERMT2-rs17125944(C) with better EM performance [68], worse baseline VA [44], and accelerated decline in PS [41]. CASS4-rs927174(C) was associated with change in GC in African-Americans [59].
For the novel loci identified by Yiu et al., Marioni et al., Janssen et al. and Kunkle et al., there were no articles that reported associations of these loci with cognitive performance. Our initial search identified 6 GWAS where summary statistics were publicly available and for which we could extract the reported associations. The HBEGF and SPPL2A gene regions were associated with GC in a multi-trait analysis of intelligence and educational attainment [40], and in two large meta-analyses inclusive of the UK Biobank [38,39, 38,39]. The ADAM10 gene region was associated with GC and ADAM10-rs889555(T) was associated with worse GC performance [39]. The KAT8 gene region was associated with AT and EF [50]. The ACE gene region was associated with EF [50], PS [39] performance in the UK Biobank, and GC [40]. The CLNK gene region was associated with PS and GC, while CLNK-rs6448453(A) was associated with worse and better EF and PS performance, respectively [39]. The CNTNAP2 gene region was associated with GC in two large meta-analyses inclusive of the UK Biobank [38, 39] and general fluid intelligence [69]. The APH1B and HESX1 gene regions were associated with PS in the UK Biobank [39]. The ALPK2 and ADAMTS4 gene regions were associated with GC in a multi-trait analysis of intelligence and educational attainment [40]. ADAMTS1-rs2830500(A) was associated with worse EF and better PS [39]. The ABI3 gene region was associated with GC [39, 60] and gF [69] while ABI3-rs28394864(A) was associated with better PS. The ACO74212.3 gene region was associated with GC and ACO74212.3-rs76320948(T) was associated with worse GC [39, 60] and better PS [39]. The OARD1 gene region was associated with AT [50] and GC [38], while rs114812713(C) was associated with better PS [39]. IQCK-rs7185636(T) was associated with worse GC performance [39]. The WWOX gene region was associated with GC [38–40] while WWOX-rs62039712(A) was associated with worse PS [39].
Association of AD GRS with cognitive performance
We found 14 studies that investigated the cumulative effect of AD risk loci on cognitive performance. Three studies investigated the effect of an unweighted GRS on cognitive performance. An unweighted GRS composed of PICALM, BIN1, and CLU, was associated with reduced EM performance [70]. In contrast, an unweighted GRS composed of the IGAP risk loci was not associated with either both cognitive performance or cognitive decline [38, 41]. Weighted GRSs that include APOE have shown more consistent results. GRS composed of SNPs identified in the initial GWAS have been associated with worse cognitive performance in EM [42, 48], EF [48], VA [46], PS [46, 48], and GC [46, 48]. Studies that have used a GRS including the IGAP LOAD risk loci have also reported associations with worse performance in EM [41, 71] and PS [41]. However, these associations largely reflect the effect of APOE as the majority are not statistically significant after the exclusion of APOE. Pathway specific risk scores for Aβ clearance, cholesterol metabolism, and immune response were also constructed but were non-significant [68].
Five studies have utilized a GRS approach, whereby a GRS is calculated based on all genome-wide significant SNPs, plus all nominally associated variants at a given significance level (PT). Two GRS (PT = 0.01) were associated with worse baseline EM and faster decline on EF and [72] and with worse EM and GC and faster decline in GC [73]. A third GRS composed of all LOAD-related SNPs (PT = 1) except for those within 500 kb of APOE was associated with worse baseline EM [74]. One study found that GRS across a range PT ranging from 1e-7 to 1e-2 was associated with faster EM and EF performance decline in Aβ+, but not Aβ- individuals [75].
DISCUSSION
This is the first systematic review to evaluate the role of non-APOE LOAD GWAS risk loci in cognitive decline. Based on a synthesis of data from 49 published studies, the results between individual risk loci and specific cognitive domains were largely non-significant for both baseline/cross-sectional cognitive performance and for longitudinal cognitive change. Of the significant gene-cross-sectional/longitudinal cognition associations that were reported (n = 128), the majority (n = 96) were not reproduced; other reviewed studies reported non-significant associations. Moreover, inconclusive patterns emerged for significant associations that were reproduced by one or more studies. Specifically, three reported significant effects in the same direction, three reported significant associations, but with inconsistent directions of effect, 12 were reproduced as significant by studies that did not report the direction of effect, and finally, 12 were reported as significant but no direction of effect was reported. However, it should be noted, where significant associations were reported and reproduced, the majority of further replication studies reported non-significant associations results. Overall, global cognition was the most extensively examined cognitive domain, with 77/255 significant associations reported. This low rate of significance and the concomitant lack of reproducibility of significant associations were observed across all the cognitive domains.
In contrast to univariate and gene-based analysis, we found more studies reporting consistent significant results of genetic risk scores associated with episodic memory performance. GRS composed of GWAS top hits and APOE were associated with worse cognitive performance in episodic memory, with 4/7 cross-sectional studies and 4/4 longitudinal studies reporting significant associations. However, these effects were largely driven by APOE, with only 2/7 baseline associations and 1/4 longitudinal associations retaining significance after APOE was excluded from the GRS. GRS composed of all nominally associated variants at a given significance level were also consistently associated with worse episodic memory performance, with 5/6 of the studies reporting significant associations. Given these results, future studies should focus on the use of GRS rather than individual variants, where the effects are likely too small to be reliably detected in a univariate analysis [76]. Furthermore, aggregating risk variants based on biological function may offer a more powerful approach to evaluating the association of genetic variants with specific endophenotypes [68].
Sample size/statistical power
A major limitation of the reported studies is small sample sizes and consequently low statistical power. In order to detect a genetic variant explaining 1% of cognitive variance at 80% power, early analyses suggested a sample size of 800–1,000 [77], but more recent genome-wide associations analyses estimate 10,000–15 000 is required [78]. Of the included studies, 37/49 had a sample size greater than 1,000, but only 9/49 studies had greater than 10,000. The two largest GWAS of cognitive performance to date, conducted as a meta-analysis of the UK Biobank and other consortia (n = 300,486 [39] & n = 269,867 [38]), found three LOAD gene-regions reaching genome-wide significance: MEF2C, HBEGF, and SPPL2A. However, it should be noted that HBEGF and SPPL2A were associated with dementia proxy case/control status in the UK Biobank and in both of these studies the majority of the samples (∼30%) originated in the UKBB. The UK Biobank has two limitations relevant to this review: it is limited to a cross-sectional design and the cognitive assessments used are brief non-standard tests that are susceptible to floor/ceiling effects [79]. Future studies, particularly longitudinal studies, should recruit larger sample sizes, or alternately, greater efforts should be made to harmonize data across studies to facilitate meta-analysis.
Phenotypic heterogeneity
Phenotypic heterogeneity between studies due to the use of different cognitive tests can limit replication [61]. While cognitive test results are highly correlated, some tests may lack the sensitivity to identify associations with small effect sizes, such as Mini-Mental State Examination (MMSE) [80], a commonly used GC test. MMSE was designed as a screening test for dementia and not a measure of cognitive abilities. It therefore exhibits strong ceiling effects, limiting its ability to differentiate between medium and high cognitive performers [81]. There was vast between-study variability in the specific measures used to assess the different cognitive domains. Although most of the cognitive measures used were psychometrically sound, replication of genetic effects on a specific cognitive domain may have been tested using measures that differed in validity, reliability, or sensitivity [82]. Additionally, when evaluating the effects of AD risk loci on cognitive aging a broad range of relevant cognitive domains should be assessed using multiple cognitive tests per domain. The construction of latent variables or composite scores offer several advantages over using single cognitive tests scores [83]. For example, latent variables use multiple indicators, rather than a single measure, thus representing a more comprehensive cognitive construct that by design reduces the impact of varying psychometric properties [84]. Alternatively, when examining cognition as an endophenotype of LOAD, a cognitive test battery focused on cognitive domains more directly affected in pre-clinical AD, such as episodic memory, may be warranted. Given these findings, future studies should 1) focus on specific cognitive domains rather than global tests; 2) choose cognitive tests specifically for their sensitivity to measure subtle cognitive differences; 3) use multiple tests to assess cognitive function of a single domain; and 4) be robust to test-retest effects.
Sample characteristics
Variation in sample characteristics such as age, sex, education, ethnicity, and medical comorbidities can limit replicability. In particular, inclusion/exclusion of individuals who develop dementia during a study may affect results. Of the studies included in this review, 26/49 were conducted in non-demented populations, 11/49 included participants with prevalent or incident dementia, while 12/49 studies did not report the cognitive status of its participants. The reported associations of LOAD risk loci in populations that retain prevalent or incident cases of cognitive impairment may be driven by pathological cognitive decline [61, 85]. In contrast, in studies that selectively exclude participants with a clinical diagnosis of dementia, the inadvertent inclusion of individuals in prodromal stages of dementia may also drive the reported genetic effects [85]. Evidence to suggest this effect has been reported in studies that separately assessed associations in participants who eventually converted to dementia and those who remained cognitively normal for ABCA7, EPHA1, and CLU. Similar effects have been observed for APOE*ɛ4 carriers [85]. In cognitively normal APOE*ɛ4 carriers, participants with a high Aβ PET levels experienced a faster rate of decline then carriers with low Aβ PET levels, suggesting that cognitive decline observed in APOE*ɛ4 carriers reflects the effect of APOE exacerbating Aβ-related decline rather than an APOE-independent effect [86]. Accordingly, future studies should evaluate the association of LOAD risk loci with cognitive function using neuroimaging or cerebrospinal fluid biomarkers to inform the classification of preclinical AD in ‘cognitively normal’ individuals. Furthermore, sensitivity analysis should be conducted to evaluate if the inclusion/exclusion of participants with MCI or dementia drives potential association of genetic variants on cognitive function.
Limitations
There are several limitations to this review. First, the heterogeneity in the methodologies (cognitive tests, genetic polymorphisms, and study design) of the included studies precluded performing a meta-analysis, which would offer increased power to detect associations and increased precision in the estimation of the magnitude of the effect. Second, we emphasize that we have reported significant associations that were p < 0.05 but as such the number of ‘true’ associations is probably smaller than the number reported here due to multiple testing and undetected publication bias. Third, the literature search used a single database, PubMed, which could limit the sensitivity of our search strategy. However, PubMed is by far the most populated database for publications for general medical and biomedical science offering a higher likelihood of retrieval of relevant publications. In addition, we followed up reference lists for all included studies and this retrieved less than 5% of studies eventually included, suggesting an acceptable sensitivity for the bibliographic database searches. Finally, while we adapted our search strategy from a published filter for detecting causation studies that favored sensitivity, it is possible that not all relevant studies were identified as our search strategy relied on the gene names or SNP identifiers being present within the title or abstract of a publication.
Conclusion
This is the first study to systematically evaluate the role of non-APOE LOAD risk loci with cognitive performance and decline. We found that the majority of associations between individual LOAD risk loci and cognitive function were non-significant, suggesting that current samples sizes are too small to detect individual risk loci effects on cognition. In contrast, consistent findings were observed for GRS, with increased LOAD genetic risk associated with deleterious effects on episodic memory performance and decline. Future research should focus on the use of GRS, recruitment of larger sample sizes or harmonization of findings across studies, and improved phenotyping of cognitive abilities. Consideration of these factors in future study designs may allow for more reliable associations of LOAD-related genetic variants with ageing-related cognitive performance and change.
Footnotes
ACKNOWLEDGMENTS
SJA is funded by the Australian Research Council Centre of Excellence in Population Ageing Research, ARC grant CE1101029. KJA is funded by NHMRC Research Fellowship No. 1002560. RAD acknowledges funding from the National Institutes of Health (National Institute on Aging, R01 AG008235) and the Canadian Consortium on Neurodegeneration in Aging (with funding from the Canadian Institutes of Health Research and partners).