
Abstract
As we age, our olfactory function declines. In addition to occurring in normal aging, more rapid decrement of olfactory decline has been associated with several neurodegenerative diseases including Alzheimer’s disease (AD) and Parkinson’s disease (PD). It has been argued that since olfactory deficits occur less frequently or are absent in diseases such as progressive supranuclear palsy, corticobasal degeneration, and multiple system atrophy, olfactory deficits can be used for differential diagnoses of AD and PD. The purpose of this review is to provide a survey of current knowledge about the molecular bases and differential patterns of olfactory deficits present in normal aging, AD, and PD. As substantial research has been conducted in this area, the majority of the content of this review focuses on articles published in the past decade. We hypothesize that olfactory deficits in normal aging, AD, and PD may have different underlying causes, and propose the use of model organisms with small, tractable nervous systems and/or easy to manipulate genomes to further investigate the cellular mechanisms responsible for these deficits.
INTRODUCTION
Our sense of smell is essential for recognizing environmental hazards, and it has a considerable impact on flavor detection and appetite. Under normal conditions, odorant molecules enter the nose and reach the olfactory epithelium (OE), where they diffuse through the mucus and bind to odorant binding proteins that activate G-protein coupled olfactory receptors (GPCRs) (Fig. 1) [1]. Odorants bind to GPCRs located on the cilia of olfactory receptor neurons and activate cyclic adenosine monophosphate (cAMP), leading to the influx of sodium and calcium ions, ultimately generating an action potential [1, 2]. This signal is relayed to the glomeruli of the olfactory bulb (OB), primary olfactory cortex, and then arrives at the orbito-frontal cortex for conscious perception [2]. There are thousands of glomeruli in young individuals; however, this number decreases with age [3]. It is interesting to note that olfaction is the only sensory modality that is not processed in the thalamus.

Human Olfactory System: The olfactory epithelium is found in the nasal cavity and contains olfactory receptor cells. The olfactory bulb is located in the forebrain on the inferior side of the cerebral hemisphere. Axons of the olfactory receptor cells in the olfactory epithelium terminate in the olfactory bulb and activate olfactory bulb neurons in clusters known as glomeruli. These glomeruli are formatted from receptor cell branching ends and interneuron branches called mitral cells. Once the odorant molecules enter the nose and reach the olfactory epithelium that houses olfactory receptor cells, cyclic adenosine monophosphate (cAMP) is activated and an action potential is generated.
Olfactory functioning (as measured by the ability to detect odorants at different thresholds, identify, discriminate, and recollect them; Fig. 2) decreases with age [4, 5]. The human olfactory system is dynamic with continuous neurogenesis of nose sensory neurons and OB inhibitory interneurons [6]. The olfactory system relies on the hippocampus and on intact neurogenesis of the OB and the neuroepithelium [7, 8]. Although human OE undergoes neurogenesis throughout life, adult hippocampal neurogenesis has been an area of debate [9, 10], with a recent study showing that adult neurogenesis is present in the hippocampus of healthy human subjects [11]. This continuous plasticity of the olfactory system persists throughout adulthood and there is a wide consensus that newly differentiated neurons are found among the elderly [12]. However, neurogenesis declines with advancing age and most people over the age of 65 present with hyposmia or anosmia [13, 14].
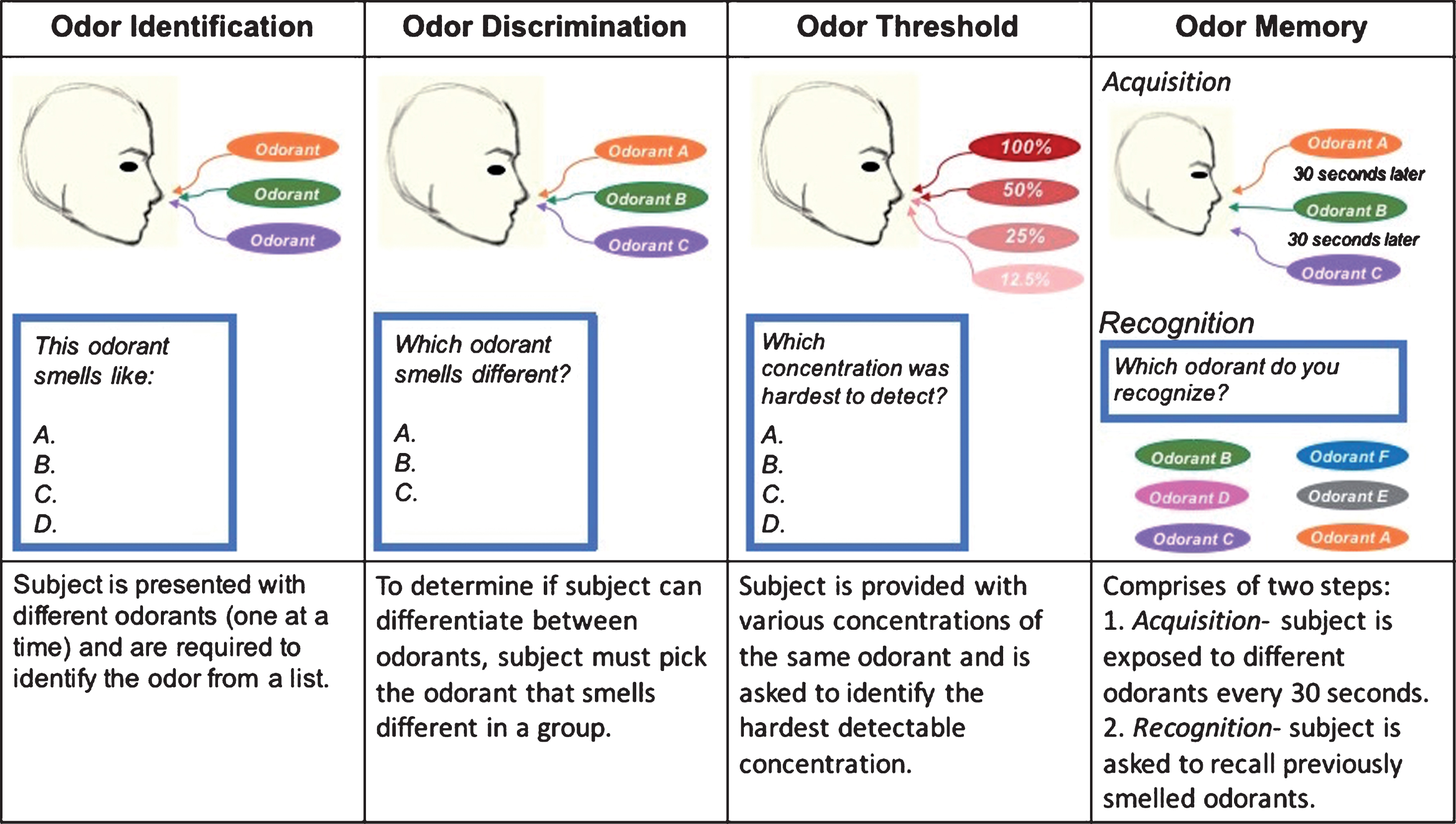
Olfactory Functioning: Assessed by measuring four odor-related tasks: identification, discrimination, threshold, and memory.
In addition to decreased olfactory functioning in normal aging, recent research has also focused on the role of olfactory deficits in two of the most common forms of neurodegenerative disorders, Alzheimer’s disease (AD) and Parkinson’s disease (PD). Although AD and PD have been primarily diagnosed using observable clinical symptoms [15], neuropathology and neuroimaging data have provided evidence that the pathological onset of these diseases begin years before the manifestation of clinical symptoms. Olfactory impairments in AD and PD are more prevalent than other non-cognitive and non-motor features of these diseases, and they occur earlier than many clinical symptoms. Previous studies have demonstrated that in conjunction with other robust tests, olfactory impairments can be used as preclinical biomarkers of both AD and PD [16], predicting a clinical diagnosis in the upcoming years with high sensitivity and specificity [17, 18]. As a result, multiple olfactory tests have been proposed (Table 1), including the University of Pennsylvania Smell Identification Test (UPSIT) [19]. This review will discuss findings for olfactory deficits in normal aging (Table 2), AD (Table 3), and PD (Table 4), and will compare and contrast the similarities and differences between olfactory deficits in these groups (Table 5).
Commonly Administered Olfactory Tests
UPSIT, University of Pennsylvania Smell Identification Test; B-SIT, Brief Smell Identification Test; SDOIT, San Diego Odor Identification Test; CCCRC, Connecticut Chemosensory Clinical Research Center Test; SOIT, Scandinavian Odor Identification Test.
Articles assessing age-related olfactory deficits
B-SIT, Brief Smell Identification Test; MRI, magnetic resonance imaging; Aβ, amyloid-β; APOE, apolipoprotein E; UPSIT, University of Pennsylvania Smell Identification Test; OSN, olfactory sensory neuron; OB, olfactory bulb.
Articles assessing Alzheimer’s disease-related olfactory deficits
AD, Alzheimer’s disease; UPSIT, University of Pennsylvania Smell Identification Test; fMRI, functional magnetic resonance imaging; MCI, mild cognitive impairment; SMI, subjective memory impairment; APOE, apolipoprotein E; OB, olfactory bulb; aMCI, amnestic mild cognitive impairment; Aβ, amyloid-β; OE, olfactory epithelium; AβPP, amyloid-β protein precursor.
Articles Assessing Parkinson’s Disease-Related Olfactory Deficits
PD, Parkinson’s disease; MSA, multiple system atrophy; PSP, progressive supranuclear palsy; B-SIT, Brief Smell Identification Test; SAI, short latency afferent inhibition; MCI, mild cognitive impairment; Aβ, amyloid-β; PET, positron emission tomography; OB, olfactory bulb.
Summary: Olfactory Impairments in Normal Aging, Alzheimer’s Disease, and Parkinson’s Disease
SEARCH STRATEGY AND INCLUSION CRITERIA
A literature search of English-language studies of olfactory impairments in aging, AD, and PD was conducted via electronic databases (PubMed, Medline, Web of Science, and Google Scholar) with a focus on studies published within the last decade. Search terms included “olfaction”, “aging”, “neurotransmitters”, “air pollution”, and all related genetic and pathologic terms associated with AD and PD, including “apolipoprotein E”, “presenilins”, “amyloid-β (Aβ)”, “amyloid-β protein precursor (AβPP)”, “tau”, “neurofibrillary tangles”, “α-synuclein”, “Lewy bodies”, “leucine-rich repeat protein kinase 2 (LRKK2)”, “parkin ubiquitin E3 ligase (PRKN, or PARK2)”, and “PTEN induced putative kinase 1 (PINK1)”. Original articles with adequate data of control and experiment groups, and review articles with highlighted opinions were included.
OLFACTORY DEFICITS IN AGING
Age is the most common risk factor for atrophic changes and impairments of the OE [20]. As we age, our olfactory functions decline and this decline is accelerated after the age of 70 years [21]. Approximately 50% of individuals between 65–80 years of age present with some type of olfactory dysfunction, and this trend increases to 80% in those over the age of 80 years [13, 22]. Aging results in the thinning of the epithelium and a decrease in the number of olfactory receptor cells due to a decline in cell turnover as well as apoptosis [3]. The volume of the OB also decreases with age and this reduction has been associated with a decline in olfactory functioning [23].
Other factors have also been associated with olfactory loss. One of the most common causes of olfactory deficits in individuals without neurodegenerative diseases is viral infections [24]. In one study, biopsies of 13 patients with olfactory disorders after upper respiratory viral infections (URVI) showed reduced numbers of olfactory receptor cells and nerve bundles [25]. Later, Lalancette-Hebert et al. used transgenic mice to investigate the transcriptional activation of microglia and toll-like receptor 2 (TLR2), key intermediaries in responding to pathogens [26]. They found that after transcriptional induction and activation of TLR2/microglia in response to focal cerebral ischemic injury of the left middle cerebral artery (MCA), pro-inflammatory responses were not restricted to the site of injury, but also spread to the OB. Exposure to environmental factors such as airborne pollutants have also been implicated in olfactory deficits. Calderon-Garciduenas et al. examined 62 healthy individuals living in Mexico City with a high level of air pollution and 14 healthy individuals living in a less polluted environment. They found that exposure to air pollution for a sustained period of time was associated with olfactory deficits, respiratory tract inflammations, and brain abnormalities [27]. There is also considerable data suggesting that the ability to identify odorants is influenced by sex and history of cigarette smoking in both young and old individuals [28]. Therefore, olfactory deficits in individuals without neurodegenerative diseases may occur as secondary symptoms due to complications such as infections of the upper respiratory tract, history of smoking, or exposure to airborne pollutants (Fig. 3).

Reasons for olfactory deficits in normal aging.
Moreover, positive associations between olfactory deficits and mortality rates have been demonstrated [29]. It is possible that olfactory impairments predict mortality as it is a marker of potential underlying neurodegenerative predispositions [30]. A recent cohort study by Liu et al. including 2,289 adults aged 71–82 years at baseline reported that compared to those with good olfaction, participants with poor olfaction were at a 46% increased risk of death by year 10 (risk ratio 1.46, 95% CI 1.27–1.67) and a 30% increased risk of death by year 13 (risk ratio 1.30, 95% CI 1.18–1.42) [31]. In fact, olfactory dysfunction is one of the best predictors of 5-year mortality rates in elderly adults [32]. Gopinath et al. found that over 5 years, 21.8% of their 1,636 participants with olfactory impairments died compared to less than 10% of their participants with normal olfaction [33]. Pinto et al. demonstrated that after controlling for age, sex, and health related variables, anosmic elder subjects had a rate of mortality that was three times higher compared to individuals with normal olfaction. This pattern may be indicative of a decrease in cellular regeneration or a biomarker of environmental toxin accumulation [32]. Interestingly, the correlation between risk of death and olfactory deficits surpasses risks such as heart failure, lung disease, and cancer [32]. In a longitudinal study of 1,169 participants, Devanand et al. investigated the association between those whose olfactory function declined most with age and mortality rates [34]. After controlling for age and sex, a positive correlation between lower UPSIT scores (indicative of anosmia) and mortality rates was reported (a 45% mortality rate in anosmic participants versus 18% in normosmic participants). This association between mortality rate and olfactory deficits was supported independent of dementia [35]. Loneliness and depressive symptoms among older individuals are also associated with worse olfaction [36]. Therefore, it is recommended that elders presenting with olfactory impairments be assessed for mental well-being. A list of current findings of olfactory deficits in normal aging can be found in Table 2. Summarizing recent literature of olfactory deficits in normal aging suggests multiple potential causes for a decrease in olfactory functioning, which result from a combination of etiologies such as OE degeneration, disruption of olfactory receptor cells, shrinkage of the OB volume, and incidents such as infections, high air pollution, and smoking.
OLFACTORY DEFICITS IN ALZHEIMER’S DISEASE
AD is the most common cause of dementia that impacts memory, thinking, and behavior. Olfactory losses in AD are often drastic and most likely the result of central olfactory system degeneration [37]. Ryo et al. studied 318 psychiatric outpatients and their first-degree relatives who all underwent open essence olfactory testing [38]. Participants were asked to identify 12 individual odorants as an assessment to their olfactory function. Those diagnosed with AD had more severe olfactory deficits compared to participants with other neuropsychiatric disorders [38]. In a cross-sectional study, Woodward et al. investigated the use of olfactory deficits as a biomarker of AD [39]. The UPSIT was used to measure smell identification in 262 AD patients, 110 patients with amnestic mild cognitive impairments, and 194 controls. It was found that disease status and odor identification ability were significantly correlated. Also, a total of 36.4% of participants with impaired olfaction converted to AD compared to only 17.3% of participants with intact olfaction (p = 0.03) [39]. Further studies of olfactory deficits in AD patients indicate that odor recognition and odor identification are more impaired than memory for visual stimuli, suggesting that AD-related olfactory impairments are not the result of a global memory deficit [39–42]. Therefore, it has been proposed that olfactory impairments in combination with other robust AD tests may be a useful screening tool for early AD diagnosis.
Moreover, AD-associated olfactory deficits tend to be highly correlated with cognitive state. Individuals with a rapid progression of olfactory deficits develop worse cognition, lower independence, and in general more severe AD symptoms than those with slower progression. Roberts et al. examined the association between impaired olfaction and incidents of mild cognitive impairments in 1,630 cognitively healthy participants [43]. They found that olfactory impairments were associated with both amnestic mild cognitive impairment and its progression to AD dementia [43]. Devanand et al. reported that in individuals without dementia, olfactory decline was associated with AD diagnosis 2–5 years prior to clinical symptoms [44]. In the Heinz Nixdorf Recall (HNR) study, 2,640 randomly sampled participants aged 45–75 years underwent cognitive assessment and olfactory testing every 5 years [45]. The results showed that anosmic participants had the worst cognitive performance and normosmic participants had the best cognitive performance. Moreover, in a study of 57 individuals with sporadic AD and 24 older individuals without dementia, UPSIT scores at baseline were correlated with Mini-Mental Sate Examinations (MMSE, assessing cognitive ability) [46]. These data are similar to what is seen in healthy older adults: olfactory impairments also predict faster cognitive decline. Recently, Dintica et al. investigated 380 dementia-free participants [47] who underwent olfactory testing, completed annual cognitive assessments, and were followed up for 15 years. Compared to normal olfaction, olfactory impairments were associated with a faster rate of cognitive decline [47]. In an earlier United States (US) national longitudinal study of 2,906 older adults, olfactory identification was measured at baseline to study its predictability for future dementia development [48]. It was found that participants with olfactory dysfunction were twice as likely to develop dementia 5 years later than normosmic participants [48]. However in AD, olfactory decline is more rapid and happens at a similar rate compared to cognitive decline [49]. Therefore, the severity of olfactory deficits in AD patients seems to be associated with the rate of cognitive decline.
It has also been suggested that genetics plays a role in AD-associated olfactory impairments. Apolipoprotein E (APOE) is a human gene located on chromosome 19 with three common alleles: ɛ2, ɛ3, and ɛ4 [50]. The ɛ4 allele is strongly correlated with sporadic AD. Up to 60% of those with two copies ofɛ4 will develop AD by the time they reach 85 years, compared to 15% of the general public [51]. Apolipoprotein E is associated with cholesterol transportation and is expressed in the OB, the OE, as well as in the rest of the central nervous system [52]. In a study of 38 elderly healthy participants, APOE4 carriers showed more impairments in odor recognition memory compared to ɛ4 negative healthy participants [53]. In the same study, female APOE4 carriers with AD had more odor recognition memory impairments compared to APOE4-negative females with AD. In a later study of 1,236 individuals aged 45–80 years, APOE4 carriers aged 70–80 years showed more olfactory deficits than non-carriers, even after controlling for their cognitive ability, suggesting that the ɛ4 allele has a negative impact on the olfactory functioning of older individuals [54]. An association between dementia and olfactory deficits was also observed, yet dementia did not mediate the effect of APOE4 and age, suggesting that non-demented ɛ4 carriers with olfactory deficits may not necessarily develop cognitive decline in the future. Supporting these findings, another study of homozygous APOE4 /4 elderly adults with AD demonstrated odor identification and odor memory deficits. In comparison with APOE4 /3 heterozygous and ɛ3/ɛ3 homozygous, APOE4/4 homozygous adults may have increased impairments in these olfactory domains [55]. Also, there may be an interaction between mutations in APOE4 and environmental stressors as carriers of the APOE4 allele who lived in Mexico City with a high level of air pollution had amyloid-β (Aβ) accumulation, signs of oxidative stress, and mitochondrial damage in their OB neurons [56]. In summary, APOE4 may increase the risk of toxic neuropathology in the olfactory system, impairing olfaction without leading to a diagnosis of dementia.
Furthermore, Aβ protein deposits are found in high quantities in the brains of those with AD. Aβ is generated from the amyloid-β protein precursor (AβPP), an integral membrane glycoprotein that is cleaved by the gamma-secretase enzyme [57]. Many familial AD cases are caused by mutations in two genes, presenilin 1 and presenilin 2, which are part of the active site of gamma-secretase [57]. Kim et al. found that compared to healthy controls, AD patients expressed higher levels of the AβPP processing enzyme, gamma-secretase, and the gamma-secretase complex component, presenilin 2, in their OE [58]. Examination of postmortem brains of AD patients also show evidence of Aβ deposition and tau neurofibrillary tangles [59], inflammation as a result of microglia activation in response to Aβ plaques [60], cerebral hypoperfusion in both regional and global regions [61], hypometabolism in brain areas with Aβ plaques [62], and oxidative stress [63]. Moreover, Growdon et al. assessed cross-sectional associations between odor identification and biomarkers of neurodegeneration in 215 clinically normal elderly participants [64]. They found that participants’ odor identification ability was associated with markers of neurodegenerations such as smaller hippocampal volume, increased Aβ burden, and poor episodic memory [64]. Adding to these findings, a magnetic resonance imaging (MRI) volumetric analysis of an AD mouse model with double AβPP mutations showed volumetric losses in the cortex, hippocampus, pons, and the substantia nigra that preceded Aβ deposits [65]. Therefore, the deposition of Aβ in the brain’s olfactory circuit may play a role in AD-associated olfactory deficits. Many studies on mice further corroborate these findings. Xu et al. studied the anterior piriform cortex of AD mouse models and found that the presence of Aβ led to deficits in odor habituation (decreased response with repeated presentations), but basic odor discrimination was intact [66]. Odor habituation involves pathways upstream of the primary olfactory pathway [66], suggesting that odor identification deficits in AD may be the result of impairments in pairing the respective odorant to its associated label in cortical areas upstream of the olfactory circuit. In addition to behavioral changes, AβPP has been attributed to physiological changes in the OB neurons. Kim et al. used manganese-enhanced magnetic resonance imaging (MEMRI) to demonstrate that axonal transport of the OB neurons was compromised in an AD mice model with AβPP overexpression [67]. Wang et al. were able to reverse this deficit using a selective gamma-secretase inhibitor which also significantly decreased the quantities of soluble and insoluble Aβ [68]. Together, these findings support the hypothesis that AD-attributed physical/physiological changes in the brain can lead to olfactory deficits prior to the development of cognitive impairments.
Various hypotheses have been proposed regarding the etiology of olfactory deficits in AD. It has been suggested that both Aβ monomers and oligomers are behaving in a prion-like manner, spreading from the OB to other connected regions of the brain via neural connections [69]. In a recent in vivo study using mice, human insoluble Aβ monomers and oligomers were injected into the mouse OB and then tracked for potential spreading to other areas [69]. As expected, both Aβ monomers and oligomers spread to other connected brain regions and led to neuronal apoptosis. Interestingly, both findings in this study and previous in vitro studies suggest oligomeric Aβ induced more damage than monomeric Aβ [70, 71]. This shows that Aβ accumulation in the OB can spread via olfactory projections to other connected brain regions. Moreover, the role(s) of microRNAs (miRNAs) in olfaction have also been examined. microRNAs, short single stranded RNAs, play a role in guiding molecules during post-transcriptional genetic processes [72]. Once a miRNA is bound to an mRNA, the mRNA is usually degraded and translation is repressed. One such miRNA found to be elevated in brains of AD patients, miRNA-206, is known for its role in regulating muscle development and tumor suppressor functions in cancer [73]. Moon et al. investigated the impact of miRNA-206 expression level in the OE of early AD patients [74]. They found a strong correlation between miRNA-206 overexpression in the OE and patients’ cognitive assessments, suggesting that miRNA-206 may be used as a biomarker for early AD diagnosis [74].
Considering these data together, in most cases odorant identification deficits in AD are the earliest to be observed, followed by deficits in odorant detection and discrimination [41]. AD-related olfactory deficits are correlated with cognitive state [44], and APOE4 increases the risk of neuropathology accumulation such as amyloid plaques in the olfactory system [55]. Despite advancements in this area (Table 3), there is no clear consensus on the cause of olfactory impairments in AD.
OLFACTORY DEFICITS IN PARKINSON’S DISEASE
PD is a systematic degenerative disease with both motor and non-motor symptomologies. In over 90% of individuals with sporadic PD, olfactory deficits are among the earliest symptoms, typically occurring 5–10 years before clinical diagnosis [75]. Olfactory deficits seem to be more prevalent in PD than in other neuromuscular disorders and therefore may be used to differentiate between PD and disorders such as multiple system atrophy (MSA) [76] and essential tremor [77].
PD-related olfactory deficits are observed in odor detection, discrimination, as well as odor identification [78], and result from a decline in central nervous system function [79]. In PD, hyposmia is more prevalent than anosmia, and in most cases patients can respond to strong odorants [78]. In a study of 400 PD patients from Australia, Germany, and the Netherlands, 45% had anosmia, 52% had hyposmia, and only around 4% had normal olfaction [80]. Furthermore, approximately 75% of PD patients had some type of olfactory impairment even after controlling for age. In another study, 45 healthy participants and 45 PD patients between the ages of 37–79 were gender-matched and used to evaluate the diagnostic accuracy of current PD tests [81]. These tests consisted of motor evaluations assessing limb bradykinesia, tremor and balance, and non-motor tests such as depression and olfaction. Of all test results, olfactory deficits were the most accurate predictor of PD. Berendse et al. also reported that olfactory deficits in PD are dynamic and worsen with time, making olfaction a useful biomarker for disease progression in the early stages [82].
Studies have also reported correlations between the degree of olfactory deficits in PD and other features of this disorder such as executive function impairments [83], neuropsychiatric symptoms [84], and memory impairments [85]. After studying 233 PD patients assessed for olfaction and neuropsychiatric symptoms, Hong et al. found a positive correlation between olfactory deficits and emotional dysfunction [84]. In particular, PD patients with a low olfactory ability had a higher prevalence of apathy compared to those with normal olfaction [84]. This may be the result of different odorants inducing vivid emotional states, or the simultaneous loss of the dopamine and serotonin neurons that regulate emotions [86]. Patients with PD who experience mild cognitive deficits are also more likely to show severe olfactory impairments than those with normal cognition [87]. In a cross-sectional study of 248 PD patients, Morley et al. found that psychotic symptoms were more likely to be present in patients with worse olfaction and that odor identification ability was a significant predictor of cognitive performance [88]. Kawasaki et al. also studied 31 PD patients without dementia and 20 age-matched controls, and found that PD patients with mild cognitive impairments had signs of a loss of awareness about their hyposmia [89]. These data suggest that olfactory testing could help in early diagnosis of those at risk of PD.
Although olfactory screening on its own is not a suitable test for PD because of its lack of specificity, it has been suggested to be used in combination with other tests. In a study of 148 PD patients and 148 healthy controls, it was found that using only three odors from the 16 item Sniffin’ sticks olfactory test were helpful in discriminating PD patients from controls [90]. Another study that included 4,999 participants who were at risk of PD found that those with hyposmia were at a significantly higher risk of non-motor PD symptoms including depression, constipation, and rapid eye movement sleep disorder [91]. Moreover, Berendse & Ponsen have suggested taking a step-wise approach to detecting preclinical PD [92]. They prospectively studied the risk of PD development in 361 asymptomatic first-degree relatives of PD patients and showed that the combination of olfactory assessment (first step) with dopamine transporter single photon emission computed tomography (SPECT) scanning (second step) measuring nigrostriatal cell loss offered a suitable approach for early PD diagnosis [92]. In this group, hyposmia alone was associated with a 12.5% risk of PD development during a 5-year period [92]. However, this approach’s main limitations are the expense and time for conduction of SPECT scanning on many healthy individuals. Furthermore, PD is primarily considered a motor syndrome caused by a loss in dopaminergic innervation. Although an increased number of OB dopaminergic neurons have been observed in post-mortem studies of PD patients [93], the accumulation of α-synuclein, the major component of Lewy bodies (protein aggregates that serve as the pathological hallmark of PD) in the OB leads to decreased levels of dopamine neurotransmission [94]. In an observational trial, 280 participants underwent olfactory testing and dopamine transporter (DAT) imaging to find whether this two-step screening strategy could predict PD development [95]. Compared to participants with no DAT deficits, the relative risk of PD diagnosis in hyposmic participants with DAT deficits was 17.47 (95% CI 7.02–43.45) [95]. Together, these findings support the use of olfactory testing in combination with other PD biomarkers for early identification of individuals at risk of PD development.
As in AD, PD has both sporadic and familial forms. The most common forms of monogenic mutations associated with PD are linked to mutations in the leucine-rich repeat protein kinase 2 (LRRK2). Some studies on PD patients have shown that LRKK2 mutations are positively correlated with classic PD associated olfactory deficits [96–98]. Conversely, a study by Ruiz-Martinez et al. found that PD patients with LRKK2 mutations were less likely to have hyposmia than patients without LRKK2 mutations (36% versus 75%, p < 0.001) [99]. Note that this group consisted of 146 non-carriers and only 44 LRRK2 mutant carriers. Statistically, such an imbalance in sample sizes increases the risk of type II errors. However, unequal sample sizes are inevitable in studies of human subjects. Interestingly, not all mutations that are associated with PD have been linked to olfactory deficits. Studies of mutations in the α-synuclein gene with an autosomal dominant pattern of PD inheritance show inconsistent results of olfactory ability [100, 101]. Further, studies of mutations in the parkin ubiquitin E3 ligase (PRKN or PARK2) and PTEN induced putative kinase 1 (PINK1) genes that lead to familial PD report relatively normal olfactory abilities [102–104]. Likewise, in a recent study of 68 Chinese PD participants with Parkin mutations, patients had relatively intact olfactory functioning compared to non-Parkin-related PD, but they had poorer olfactory functioning than normal individuals [105]. Remarkably, in PD patients with PRKN mutations, Lewy body depositions are almost non-existent [106]. To date, the role of genetics in olfactory deficits of PD is not well understood.
In summary, it has been suggested that patients with PD often suffer from hyposmia and that anosmia is rarely observed in this group (Table 4). Further, many domains such as odor detection, discrimination, and identification are altered in PD patients. Importantly, olfactory deficits are suggested to be the most accurate non-motor preclinical symptom of PD, and the deposition of Lewy bodies in olfactory centers may play a role in these PD-associated impairments.
COMMONALITIES AND DIFFERENCES IN AD AND PD-RELATED OLFACTORY DEFICITS
As a result of similarities in the olfactory deficits in AD and PD, the question of a common etiology for these phenotypes arises [16]. Both AD and PD associated olfactory deficits seem to result from degeneration of the central nervous system (Table 5). Is there a common pathway that when disrupted, leads to olfactory deficits during the preclinical stages of AD and PD? Rahayel, Frasnelli, & Joubert carried out a meta-analysis on this topic and reported that individuals with either AD or PD have impairments in odor identification and task recognition, and those with PD have greater impairments in odor threshold detections compared to AD [107]. Notably, in AD cases, olfactory impairments are more associated with higher-order olfactory functions that require specific cognitive processes, such as odor identification and recognition [107]. Moreover, patients with AD are often unaware of their olfactory deficits [108], while those with PD are more self-aware [109]. In a study of 240 adult participants, scores on neuropsychological tests suggested that those unaware of their olfactory dysfunction as measured by the Scandinavian Odor Identification Test performed poorer on verbal learning, memory, and attention tasks compared to those with normal olfaction [110]. Furthermore, Doty et al. reported that of the 34 participants diagnosed with AD, while 90% suffered from olfactory impairments, only 6% were aware of this problem [111]. In contrast, in a recent study of 124 PD participants without dementia, and 154 elderly controls, the PD patients were aware of being hyposmic but they often overestimated their smell ability [112]. These results suggest that the underlying neuropathology leading to olfactory deficits in AD and PD are expressed in different regions of the olfactory system.
In both diseases, changes in neurotransmitter levels observed in the forebrain appear to be associated with olfactory functioning [16]. Nori et al. found that, in the absence of acetylcholine, insoluble Aβ deposits increased linearly, implying that acetylcholine is necessary for the stabilization of soluble, non-toxic Aβ [113]. In a mouse model of AD, Gil-Bea et al. induced cholinergic denervation to assess the interaction between cholinergic neurons and AβPP [114]. They reported that deficits in cholinergic neurons can lead to higher hippocampus vulnerability to neurotoxins such as insoluble Aβ. Similarly, as cholinergic denervation in PD increases, PD symptoms worsen and olfactory deficits increase [115, 116]. Moreover, the role of dopamine has also been implicated in both AD and PD. Periglomerular dopaminergic neuron densities have been found to be increased in AD and PD participants compared to controls [93]. This increase may be an intrinsic approach to compensate for the loss of neurons in AD and PD [93], and may play a role in olfactory deficits as the depositions of tau, Aβ, and α-synuclein decrease OB synaptic efficiency [117].
A decrease in OB neurogenesis has also been suggested as a cause of olfactory deficits in neurodegenerative diseases [118, 119]. As a result of impairments in adult neurogenesis, not only are existing neurons destroyed, but the brain’s capacity for neuronal renewal and plasticity is compromised. Adult neurogenesis takes place at specific brain areas, namely the subventricular zone and hippocampus’s subgranular zone of the dentate gyrus [119]. This hypothesis was primarily based on studies on rodents and was received with skepticism [120]. Reports have been contradictory about whether adult neurogenesis occurs in humans. A recent study demonstrated neurogenesis in the brains of healthy adult human subjects that was diminished in brains of those with AD [11]. However, there has been conflicting evidence that suggests that human adult neurogenesis falls to low and functionally insignificant rates with age [10]. Therefore, whether or not AD and PD-related pathology alter adult neurogenesis in humans has not yet been determined.
Despite some similarities between olfactory deficits in AD and PD, certain neuropathological differences provide support for a multifactorial etiology. For example, using high resolution MRI, it has been shown that the OB and tract volume in AD patients is significantly decreased in early disease stages compared to normal controls [121]. However, whether or not such decrements in OB volume are observed in PD is controversial [122, 123]. More recent studies have demonstrated a significant reduction of the OB volume and gray matter volumes of areas associated with olfaction in patients with idiopathic PD [123]. In contrast, a recent four-year longitudinal study of 25 PD participants and 24 normal controls reported a significant association between decreased basal ganglia volume and olfactory loss in patients [124]. Though normal controls lacked this association, the degree of olfactory loss over time was similar in normal and PD age-matched participants.
In conclusion, in both AD and PD, studies have demonstrated a decreased level of neurogenesis in the OB which may partially explain these olfactory deficits. However, different impairment levels are observed in distinct olfactory domains in AD compared to PD. Whereas most reports suggest that odor recognition and identification are most affected in AD, PD patients show more severe deficits in odor thresholds [107]. The different behavioral symptom profiles in AD and PD patients suggest that damage to brain regions leading to olfactory impairments is caused by different factors (Table 5).
ARE AD AND PD SIMPLY ACCELERATING A NORMAL AGING PROCESS?
Although age is the primary risk factor for developing neurodegenerative diseases, it is challenging to contrast normal aging with neurodegeneration as the link between the two remains elusive. As we age, brain metabolism changes, leading to the accumulation of protein aggregates, calcium signaling impairments, and mitochondria dysfunction [125]. Similar changes are observed in neurodegenerative diseases. For example, both aging and AD are associated with cognitive decline, neural activation, and changes in neurotransmission [126]. However, these changes are more pronounced in neurodegenerative diseases, proceed at an exacerbated rate, and exhibit different patterns [126].
Factors that are associated with olfactory deficits in normal aging and in neurodegenerative diseases seem to have more differences than similarities (Table 5). Most importantly, olfactory deficits that are observed in early courses of AD and PD appear to be highly correlated with declines in cognitive and motor functions, respectively. Yet some studies have demonstrated that olfactory deficits in healthy aging are not associated with cognitive decline [34]. Postmortem examinations have revealed Lewy bodies in the brains of older individuals without PD or dementia [127]. Lewy body pathology is found in 8–17% of those without neurological diseases over the age of 60 years [128]. Although pathological expression of Aβ and α-synuclein have been reported in the OB of both healthy individuals and those with AD or PD, the frequency and extent of these pathologies differ [129]. There are few substantive insights into the cellular and molecular properties of the aged olfactory system, creating a severe handicap in attempting to understand the mechanisms that underlie the decreased olfactory function in the aging population. In individuals free of neurodegenerative disease, olfactory deficits have been associated with age, gender (higher correlations seen in males), and cardiovascular and cerebrovascular diseases [13].
Although healthy carriers of APOE4 have an increased risk of developing olfactory deficits, this association is strongest in highly polluted areas [27]. However, it is important to note that individuals do not have to be carriers of APOE4 to exhibit olfactory deficits in areas with high air pollution. Furthermore, in normal aging groups, decreases in OB volume are correlated with age, while in the case of AD, volume of the OB decreases irrespective of age, and is inversely associated with disease duration [123]. This supports the notion that the shrinkage of the OB in individuals with AD is a feature of the disease itself. In contrast, reduction in cortical thickness and decreased hippocampal volume in older, cognitively normal individuals are associated with deficits in olfactory identification [130].
Additionally, the potential role of microglia, cells responsible for the brain’s innate immune system, in olfactory deficits of normal aging, AD, and PD, must not be overlooked. In normal cases, proper neuronal connectivity keeps microglia in a quiescent state [131]. At rest, ramified microglia have a spider-shaped look [132]. In an inflamed brain region, the coordination of neurons is disrupted, perhaps as a result of insoluble and toxic protein aggregates, and microglia are activated and begin releasing neurotoxins [131]. When activated, microglia adapt an amoeboid shape to facilitate migration, become phagocytic, and secrete cytokines to protect the brain from lesions or pathogens [133]. Studies have reported an exaggerated presence of amoeboid-shaped microglia in the anterior olfactory nucleus of both AD and PD patients [134]. This pathology, in combination with protein depositions found in the OB may be key contributors to these observed olfactory deficits. Chronic neuroinflammation is an important feature of both normal aging and neurodegenerative diseases [135, 136]. However, whether microglia activation of the OB is directly caused by Aβ or α-synuclein pathology is unknown. A causative link between microglia activation and AD or PD-linked olfactory deficits has not yet been demonstrated.
In the last decade, Richard L. Doty introduced the “olfactory vector hypothesis” [137]. This hypothesis posits that environmental pathogens can easily access the brain through the olfactory mucosa, and the onset of AD and PD is facilitated via these agents. These pathogens may include, but are not limited to, viruses and toxins [138]. In support of this hypothesis, studies have demonstrated olfactory impairments in individuals exposed to substantial air pollutants and in carriers of the APOE4 allele [27].
In summary, neuroinflammation is a common characteristic observed in normal aging as well as AD and PD and is exaggerated by exposure to pollutants. This may lead to microglial activation causing the release of neurotoxins and may contribute to olfactory deficits. Unlike normal aging, olfactory deficits in AD are associated with cognitive decline and olfactory deficits in PD are associated with both motor impairments and cognitive decline. Further, the risk factors of olfactory deficits that are associated with normal aging, such as gender and cerebrovascular diseases, are different than AD and PD related risk factors. Finally, the volume of the OB decreases in response to normal aging, but in AD the OB volume is inversely associated with disease duration. Therefore, olfactory deficits in neurodegenerative diseases seem to result from different underlying causes compared to deficits in normal aging.
To further examine the similarities between olfactory deficits in normal aging and neurodegenerative diseases such as AD and PD, more information regarding the role of risk factors such as sex and genetics is required in both human and mice studies. Tests of odor identification are often difficult to reproduce, although longitudinal studies of olfactory assessments may help overcome this limitation [139]. Also, research using rodents to study early pathogenic cellular pathways that precede cognitive and motor symptoms in AD and PD can help bridge knowledge gaps.
FUTURE DIRECTIONS
Over the past several years, there has been a focus on the potential efficacy of olfactory testing for early diagnosis of AD or PD. This can be especially beneficial for choosing suitable participants to enroll in clinical trials, prior to clinical symptomologies of AD and PD. However, few studies of humans and mammalian models have elaborated on the underlying cellular mechanisms that lead to these olfactory impairments. Throughout this review, we have covered the complex interactions between neurotransmitters, genetics, and the environment in relation to olfactory deficits, providing insight into the challenges that lie ahead for olfactory-related neurodegenerative diseases.
Many questions remain open for further exploration: Are Aβ, tau, and α-synuclein accumulation in the olfactory bulb causing olfactory dysfunction or are they simply markers of previous damage related to AD and PD? What roles do genetics and sex play in AD and PD-related olfactory impairments? Do AD and PD-related pathologies alter neurogenesis in adult humans? Is there a causative role between microglia activation and AD and PD-related olfactory deficits?
To answer these questions, novel techniques are being introduced in vivo and non-invasively to demonstrate the structural connectivity of the olfactory system to cortical and subcortical regions of the brain [140, 141]. Because the cellular mechanisms leading to olfactory loss are not understood, it is essential to highlight the importance of understanding the genetics behind olfactory deficits [142]. We suggest that model organisms with tractable nervous systems, short lifespans, and amenable to a large number of genetic tools and techniques, such as Drosophila melanogaster (D. melanogaster) and Caenorhabditis elegans (C. elegans), be considered to further investigate the mechanistic etiology of olfactory deficits in neurodegenerative diseases. These organisms can be especially advantageous in studying the associations between genes involved in AD, PD, and olfaction, and can elucidate the functions of disease-associated genes at the level of individual neurons. For example, in C. elegans overexpressing a human disease variant of LRRK2 (G2019S) in dopaminergic neurons only led to increased neurodegeneration of these neurons and locomotor defects [143], and the same can be said for pan-neuronal and dopaminergic neuron specific overexpression of PD-associated human α-synuclein (A53T) [144]. These models, combined with mammalian data, can provide molecular, neuronal, and behavioral level insights that will enhance our understanding of what causes the neurons to die and how those changes correlate with olfactory impairments.
Up until now, we have been successful in understanding the anatomical and pathological consequences of AD and PD pathologies after the onset of disease. Since both AD and PD appear to be influenced by genetics and by environmental triggers, the study of their etiology is complex. Cases are thought to be caused by a mixture of genetics and triggers such as environmental toxins, microbes, and sex [145]. Many of the genes implicated in AD and PD have orthologs in C. elegans and Drosophila. Both Drosophila and C. elegans have been used to express and study effects of the patient mutations in genes observed in AD and PD [146, 148]. Further, both model systems can be used to study forward and reverse genetics using mutants, transgenics, and easily made knockouts [149]. These genes can then be further studied in mammals to determine their pattern of up or down regulation in the olfactory system. For example, chemotaxis assays in C. elegans and in Drosophila have been used to investigate the effects of mutations on olfaction in worms and flies [150, 151]. Orthologs of disease genes, or genetically engineered human genes, can be manipulated at the nervous system or single neuron level in worms to investigate their roles in olfaction. C. elegans are transparent, have a short life-cycle (∼2 weeks), and have only 302 neurons. This allows neurons in mutant worms with olfactory deficits to be screened for degeneration over time using confocal microscopy [152]. The role of Presenilin 1 in the cleavage of Notch family of proteins was first discovered in C. elegans. Likewise, an example of how D. melanogaster can be used to understand functions of a gene implicated in AD is seen with studies of Sox102F where silencing Sox102F, the orthologue of SOX5, a transcription factor expressed in the heart, lung, and brain, leads to olfactory learning and memory impairments and severe neurodegeneration [153]. Meta-analyses of results for several human families affected by late onset AD and carrying SOX5 variants demonstrated significant association with AD. Four novel mutations in SOX5 have been discovered that are now hypothesized to play a causative role in AD.
Despite these advantages, there are disadvantages associated with using model organisms with small nervous systems to study the underlying causes of olfactory impairments. They lack many of the defined tissues and complex neural circuits that are found in the mammalian brain which make studying tissue-specific signaling and neural circuit abnormalities difficult [154]. Moreover, as they are evolutionarily far from mammals, some physiologically important systems such as adaptive immunity are absent in many invertebrate organisms.
CONCLUSION
In conclusion, predicting AD and PD only using olfactory testing is a controversial topic. However, the non-invasive nature of olfactory assessments combined with other biomarker tests may make them useful in diagnosing individuals at risk of neurodegenerative diseases. As olfactory impairments are among the first symptoms of AD and PD, understanding their etiology may also lead to important insights regarding the onset of these neurodegenerative diseases. Animal models will play a crucial role in understanding the mechanisms of olfactory decline in normal aging and neurodegenerative disease. In the future, this information could potentially aid in diagnosing, preventing or slowing the progression of AD and PD.
Footnotes
ACKNOWLEDGMENTS
The authors would like to thank Dr. Weihong Song, Troy McDiarmid, and Joseph Liang for helpful comments on an earlier draft of this manuscript. Partial funding for this work came from a Project Grant (#CIHR MOP 130287) from the Canadian Institutes for Health Research (CIHR) to CHR.