
Abstract
Exercise has been shown to be protective against the risk of dementias, including Alzheimer’s disease (AD). Intervention studies have demonstrated its ability to mitigate cognitive and behavioral impairments and reduce disease in both humans and animals. However, information is lacking in regard to the volume and intensity, as well as timing of exercise onset with respect to disease stage, which produces optimal benefits. Here, utilizing the Tg2576 mouse, a model of AD-like parenchymal amyloid pathology and cognitive impairment, we sought to understand the effects of different lengths of daily access to a running wheel on advanced stage disease. This study is the first to determine the benefits of long-term exercise (4 months of voluntary running) and different periods of daily access to a running wheel (0 h, 1 h, 3 h, and 12 h running wheel access) beginning in 14-month-old Tg2576 mice, an age with significant amyloid pathology. We found that exercising Tg2576 animals showed lower levels of some aspects of AD pathology and reduced behavioral dysfunction compared to sedentary Tg2576 animals. High intensity exercise, rather than high volume exercise, was generally most beneficial in reducing amyloid pathology. Our results suggest that engaging in vigorous exercise programs, even after living a sedentary life, may lead to a measurable reduction in AD pathology and preservation of some cognitive abilities.
INTRODUCTION
Alzheimer’s disease (AD) is a progressively debilitating neurodegenerative disorder affecting nearly 50 million individuals worldwide [1]. Currently, there are five FDA approved drugs (Razadyne, Exelon, Aricept, Memantine, and Namzaric) available to AD patients [2]. However, these pharmacological interventions are only beneficial in alleviating some AD symptoms and do not aid in blocking the progression of the disease [3–5]. Furthermore, no new drugs have successfully passed through clinical trials for more than 20 years [2]. This has led researchers to consider alternative options for AD prevention and treatment, such as modifiable lifestyle factors [4, 5]. One such lifestyle factor, cardiovascular exercise, is particularly promising for its ability to mitigate cognitive and behavioral impairments and reduce AD pathology in both human and animal studies [6, 7]. Prior studies have shown that regular physical activity promotes brain function through a variety of mechanisms, including enhanced antioxidant and oxidative damage repairing systems, exercise-induced upregulation of neurotrophins involved in neurogenesis-improved memory and brain plasticity, and increased resistance to stress and relieved depression [7–9]. Additionally, epidemiological and clinical research has pointed to the positive benefits of physical activity in lowering the risk of cognitive impairments [10].
Most previous studies have focused on the effects of cardiovascular exercise regimens on emerging AD symptomology to better understand the causal mechanisms by which it may produce beneficial effects on specific AD pathology. Some have found that exercise can increase the degradation of amyloid-β (Aβ; e.g., [11, 12]), a prominent hallmark of AD [13]. In addition to the pathological benefits, behavioral improvements have been observed by some, but not all, studies. For example, long-term voluntary exercise (unlimited access to running wheels) in younger mice improved learning on the Morris water maze and reduced Aβ deposition [11, 15]. Importantly, these studies model a disease preventative approach in humans. It is critically important to also understand if cardiovascular exercise can serve as a treatment intervention, remediating established pathology and behavioral impairments.
While this suggestive evidence exists of benefits of cardiovascular exercise on emerging AD pathology and behavioral deficits, even here important parametric questions have not been answered systematically or with attention to what human analog is being modeled. For example, while both short-term and long-term interventions have been assessed in younger AD animal models (when the pathology was in the early stages), less is known about the effects of exercise in models of more advanced disease stages. The question of whether well-established pathology can be reversed through exercise intervention in older humans is an important one, given the extra considerations and risks of initiating an exercise program in this population [16–18]. Moreover, the duration and intensity of exercise that produces optimal effects in aged animals is also an important issue to address, since it provides preclinical insights as to how much and how long one might have to exercise to see benefits in humans.
The present study sought to investigate these questions by testing the effects of long-term voluntary exercise training at variable intensities and volumes on behavior and Aβ deposition in older Tg2576 mice at an advanced disease state, when Aβ plaques and cognitive impairments are present [19]. We based our running parameters on our previous work in wild type (WT) and transgenic AD mice [20, 21], in which we also demonstrated reduced neuroinflammation and improved behavioral outcomes with exercise in TgSwDI mice, a model of cerebral amyloid angiopathy [20]. Wild-type C57BL/6 and Tg2576 mice had access to a running wheel for 0 (sedentary), 1, 3, or 12 hours per day. The 1-h access period may best model the relatively modest minimal recommendation for physical activity made by the CDC [22]. To model an intervention in previously sedentary older adults with advanced disease, the exercise intervention began when the mice were 14 months of age and continued for four months.
MATERIALS AND METHODS
Animals
Mice were obtained at 14 months of age and housed in a controlled room (22±2°C and 40–60% humidity) with a 12-h reverse light-dark cycle (lights off 0800 h). A total of 72 animals were split equally between males and females. Tg2576 (SED: 8, 1h: 9, 3h: 8, 12h: 9) and C57BL/6 (SED: 9, 1h: 10, 3h: 9, 12h: 10) mice were randomly assigned to experimental and control groups. After 4 months of exercise intervention, mice underwent various behavioral tests before the brains were harvested for histology and ELISA. Purina Lab Diet rat chow was available ad libitum for the entire experiment, and body weight and food intake were recorded weekly throughout the entire experiment. All experiments were conducted in conformity with the NIH Guide for Care and Use of Laboratory Animals (8th Ed.) and approved by the Stony Brook University Institutional Animal Care and Use Committee.
Tg2576 mice
The Tg2576 mouse model of parenchymal amyloid pathology and cognitive impairment expresses the human amyloid precursor protein (APP) harboring the Swedish familial AD mutations. The cognitive deficits and pathological characteristics of Tg2576 have been well characterized [19], making this model appropriate for studying AD amyloid pathology. Prior research has shown that Tg2576 mice develop Aβ plaques by 11–13 months of age and cognitive deficits by 9-10 months of age [19]. The exercise intervention was initiated at 14 months of age. The intervention lasted for 4 months, and mice were euthanized at 18 months of age. Thus, starting exercise treatment when Tg2576 mice were 14 months of age represents a moderately advanced disease state.
Voluntary exercise intervention
Exercise mice were transferred from their home cages to larger cages equipped with running wheels 5 days per week for 4 months for either 1, 3, or 12 h per day according to their assigned groups. Sedentary mice remained in their home cages during this time, consistent with previous studies that investigated the effects of exercise on brain and behavior [18, 21]. Voluntary wheel running was chosen over a forced running regimen in order to reduce the confound stemming from stress introduced by a forced exercise regimen [18, 23].
Behavioral assessment
All mice underwent behavioral testing 2-3 weeks after completion of the 4-month exercise intervention period. The behavioral battery included tasks designed to assess behaviors related to motor function, memory, and temperament as done previously [20, 21]. All behavior, except for rotarod, was recorded and analyzed using Any-Mazetm software.
Barnes maze
Barnes maze testing was first described by Barnes [24]. It consisted of a sealed wooden disk 91 cm in diameter, elevated 75 cm off the floor, with eight escape holes, spaced 24.5 cm apart. One of the holes led to an escape box (measuring 10 cm×8.5 cm×4 cm), the position of which varied among the mice but remained constant for a particular mouse. Visible distal cues were present in the room and remained constant throughout each day of testing. Each mouse was given two trials per day with a 15 min inter-trial interval. Testing spanned five consecutive days. When the mouse entered the escape box, the trial was stopped and they were given 1 min inside the box before being transferred back to the home cage. If the mouse did not find the escape hole within the 5-min limit, they were placed into the hole and left there for 1 min. Latency to enter the escape box was recorded during each trial.
Rotarod
The rotarod test is widely used to assess motor performance in rodents, measuring an animal’s ability to uphold itself on a rod that turns at accelerating speeds. The apparatus (model ENV-575M; Medical Associates Inc.) accelerated from 4 rpm to 40 rpm over the duration of each of the three trials (10-min inter-trial interval). The time on rod until the mouse fell (maximum time of 5 min) was recorded. Average of the best two trials was used for analysis.
Open field
Mice were placed in a square 60 cm×60 cm open field arena for 10 min. Distance traveled was measured to assess general activity and motor function and center activity was recorded to assess anxiety-like behavior. Measures of general activity and agoraphobia-related anxiety are also useful in interpretation of any differences in performance observed in the Barnes maze.
Y maze spontaneous alternation
The Y maze is a Y-shaped 3-armed maze used to measure spatial working memory [25]. Mice were introduced to the center and were allowed to freely explore the maze for 3 min. The number of arm entries was assessed as a measure of exploratory behavior in a novel and relatively complex environment (compared to the open field). Typically, rodents show a tendency to explore less visited arms. In a species typical behavior, mice should alternate traversing down arms in a circular pattern rather than repeatedly going down the same arms. Therefore, a mouse going down each of the three arms before returning to a previously visited arm is considered as one alternation. This behavior was evaluated during the trial, and number of arm entries was recorded. The order of arm entries was manually assessed, and percent alternation was calculated [# alternations/ (# arm entries – 2) *100].
Social interaction
Crawley’s three chamber paradigm was adapted to assess sociability and social memory [26]. The apparatus used is a rectangular box measuring 46 cm×21 cm consisting three compartments connected by clear walls. The side chambers measured 16 cm×21 cm, and the middle chamber measured 14×21 cm. This test comprised of three 5-min trials that occurred consecutively, with the first trial serving as habituation to the chamber. In the second trial, the middle chamber was empty, one side chamber contained an empty cup, and the other side chamber housed another mouse of the same sex in an identical cup. Sociability was assessed by comparing time spent with the mouse cup compared to the empty cup. In the third trial, a new mouse was housed in the empty side of trial two. Social memory was assessed by comparing time spent with the novel mouse cup versus familiar mouse cup.
Marble burying
Marble burying was used to assess species typical behavior such as burying harmful objects in bedding [27]. Experimental procedures were adapted from Deacon (2006)’s protocol. Mice were placed in a rat-sized tub cage filled with 5 cm of corn cob bedding and 20 marbles in a 5×4 array for 5 min. Time spent digging was recorded. Digging was defined as coordinated movements of fore or hind limbs that displaced the bedding, and marbles were considered “buried” if they were 75% deep in the bedding.
Object displacement and novel object recognition task
A joint object displacement and novel object recognition task was performed. Object displacement was used to assess spatial memory, and novel object recognition was used to assess object recognition memory. This task was performed in three trials, each lasting five minutes, with an inter-trial interval of 15 min. In the first trial, two of the same objects were placed in the open field arena, one in the middle of the front left quadrant, and one in the middle of the front right quadrant. In the second trial, one of these objects had its position moved to the back quadrant of the same side (left or right) of the arena, while the other remained in the same location as the first trial. Ability to differentiate the novel location of the object was evaluated as ratio of time spent with object in novel location (displaced object) compared to object in the same location as the first trial. In the third trial, a novel object replaced the object that was displaced in the second trial. Novel object recognition was assessed by the ratio of time spent with novel object compared to the object that remained the same (familiar object).
Light Dark Box
The Light Dark Box measured anxiety-like behavior. Mice were placed in the light-dark box apparatus for 5 min. The apparatus is rectangular in shape, measuring 43 cm×21 cm. Within the box, there are two chambers separated by a small doorway. The “light” chamber measures 29 cm×21 cm and has clear plexiglass walls. The “dark” chamber measures 14 cm×21 cm and has opaque black walls (dark). An opaque black plexiglass wall separated these chambers with a 5 cm×5 cm hole cut in the middle to allow for transitions between the two. Anxiety-like behavior is assessed by the time spent in the light chamber.
Biochemical and histological measures
Enzyme linked immunosorbent assay (ELISA) for Aβ species
ELISAs were performed to quantify whole forebrain levels of soluble (TBS-extracted) and insoluble (guanidine-extracted) forms of Aβ40 and Aβ42 in Tg2576 mice, similar to previous studies [28, 29]. Only Tg2576 mice were used for this analysis because WT mice do not develop amyloid pathology. Brain hemispheres were flash frozen, pulverized, and separated into two aliquots. A soluble fraction was obtained by homogenizing tissue with 10μl/mg TBS, 0.5 mm glass beads, and a bullet blender. Aliquots were centrifuged at 1600× g at 4°C for 20 min. The supernatant was removed, which was the soluble fraction. The remaining pellet was suspended in 5M guanidine-HCl at pH 8.0, and rotated at room temperature for 3 h. Samples were centrifuged as above and the supernatant was removed, which was the insoluble fraction. For each of the two fractions, a sandwich ELISA was performed. Well plates were coated with 1μl/well of Aβ40-specific antibody m2G3 or Aβ42-specific antibody m21F12 (both generously provided by Eli Lilly). Plates were blocked and shaken overnight at 4°C. Aβ species were detected using biotinylated-m3DG (Eli Lilly), followed by streptavidin-HRP (Amdex RPN4401V). Plates were developed using SureBlue (KPL Inc.) and read with a microtiter plate reader (Spectramax).
Thioflavin-S staining and analysis
Immunohistochemical analyses for amyloid pathology were performed on Tg2576 mice brain tissue only, as WT mice do not accumulate amyloid. These procedures were performed as previously described [20]. Brain hemispheres embedded in paraffin were sectioned at 10μM thickness and mounted on glass slides. Paraffin was removed from sections by immersion in xylene (3×5 min) and rehydrated in decreasing concentrations of ethanol (100%, 95%, 70%, 50%, 0% at 5 min each). Slides were rinsed with distilled water then stained for Thioflavin-S (0.0125% Thioflavin-S in 50% EtOH/PBS) by incubating for 15 min at room temperature. Slides were rinsed 3×with distilled water then 2×with 70% EtOH, then washed for 5 min in distilled water. Anti-fade reagent in glycerol/PBS was added to each slide and cover slipped, then sealed with mounting media. Sections were imaged using an Olympus BX60 microscope with an attached Olympus Dp72 camera. Images from cortex and hippocampus were collected from each section at 40x magnification. NIH ImageJ software was used to measure the percent area occupied by fibrillar amyloid in the cortex and hippocampus. These regions were chosen as they have been previously shown to accumulate fibrillar Aβ plaques and are involved in several of the behavioral tasks used in the current study.
Statistical analysis
Two-way ANOVAs were performed to determine the differences between treatment groups on running parameters, body mass, food intake, and all behavioral measures except Barnes maze. Three-way repeated measures ANOVA was used to assess Barnes maze performance over trials. One-way ANOVAs were performed to determine the difference between treatment groups on ELISA and histology (WT mice were excluded in these analyses). Pearson correlations were also performed to assess relationships between running parameters and amyloid pathology. Survival analysis was performed using the Log-rank (Mantel-Cox) test. Analyses were performed using Graphpad Prism v8 and SPSS. Significance was set at alpha <0.05.
RESULTS
Running patterns
In a running wheel paradigm such as this, the individual animal controls the actual amount and properties of the exercise within the assigned interval. Therefore, running behavior was recorded during each session to determine total daily exercise, as well as to investigate patterns of activity throughout sessions. Rotations performed were recorded in intervals of 1 min. The average number of total wheel rotations per session was calculated to determine running volume. The average rotations/min (running speed) and the average rotations/h (running density) were calculated as measures of the intensity of the running. Analysis of rotations, speed, and density were calculated and averaged over 4 months (Fig. 1).

Running parameters. An analysis of running parameters suggests that WT and Tg2576 mice ran similarly. A) Longer access to a running wheel resulted in a greater number of rotations performed B) Mice with 3h and 12h of access to a running wheel ran at a greater speed than 1h mice overall. C) Mice with 3h access to a running wheel exhibited a greater density of running compared to 1h and 12h mice overall. @p < 0.05 versus 1h (within that GT), #p < 0.05 versus 3h (within that GT).
Two-way ANOVA was performed to assess group differences in total running volume averaged across the entire 4-month intervention period (Fig. 1A). The main effect of exercise was significant [F(2,40) = 32.9, p < 0.0001], with a dose dependent increase in volume of exercise (12 h > 3 h > 1 h; p < 0.05 for all). Post-hoc tests showed that this pattern was similar in both WT and Tg2576 mice [p < 0.05 for all except Tg2576 1 h versus Tg2576 3 h (p = 0.10)]. There was no main effect of genotype or genotype×exercise interaction.
Running speed was calculated by averaging the number of rotations performed in 1-min bins that were not counted as a “break”, essentially averaging the number of rotations performed per minute when the number of rotations performed in that bin was greater than five averaged over the entire experiment (Fig. 1B). Two-way ANOVA showed that there was a main effect of exercise [F(2,40) = 7.861, p = 0.0013], such that mice with 3 h and 12 h of access to a running wheel ran at a greater speed than 1 h mice overall (p < 0.05 for both), and post-hoc tests revealed that these differences were significant for both WT and Tg2576 mice (p < 0.05 for all). There was no main effect of genotype or genotype x exercise interaction.
Running density was assessed by calculating mean rotations per hour averaged over the entire experiment (Fig. 1C). Two-way ANOVA showed that there was a main effect of exercise [F(2,40) = 6.058, p = 0.005] such that mice with 3 h access to a running wheel exhibited a greater density of running compared to 1 h and 12 h mice overall (p < 0.05 for both), and post-hoc tests revealed that these differences were seen in both WT and Tg2576 mice [p < 0.05 for all except Tg2576 3 h versus Tg2576 12 h (p = 0.0515)]. There was no main effect of genotype or genotype×exercise interaction.
It should be noted that there was considerable variability evident in the two intensity measures across groups, and that the 1 h group represented a low volume, and relatively low intensity group when the three parameters are considered together.
Survival analysis
A very basic measure of the general health effects of a sustained exercise program would be reflected in mortality or attrition from the study (e.g., if an animal ceased running). Therefore, a survival analysis was conducted. The Log-rank (Mantel-Cox) test found no significant difference between survival curves χ2 (7, N = 71) = 7.5 p = 0.379 of the WT groups or Tg2576 groups (Fig. 2). While not significant, it may be noted that the exercising groups all showed lower levels of mortality at the 150d time-point.
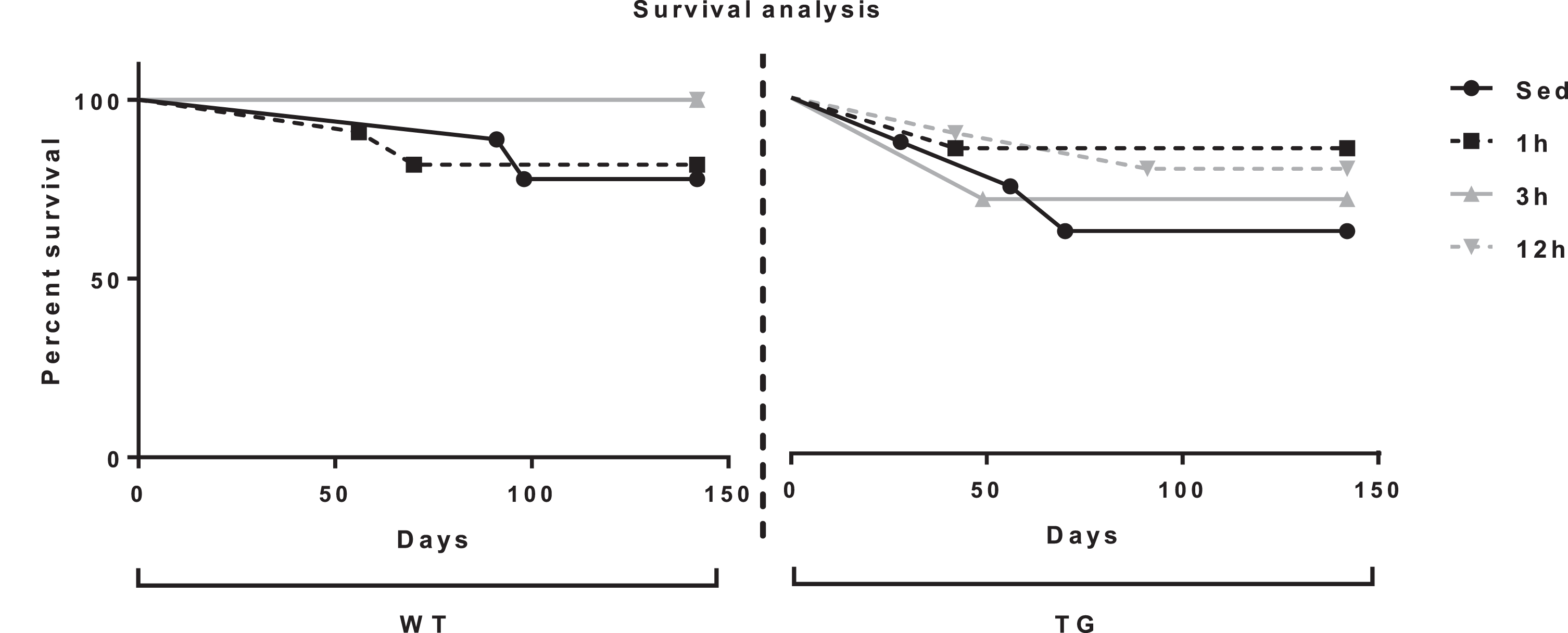
Survival curves. The Log-rank (Mantel-Cox) test found no significant difference between survival curves though exercising groups trended toward reduced mortality.
Body weight and food intake
Body weights and food intake levels also serve as a basic measure of the physiological response of the animals to sustained exercise. Two-way ANOVA showed that was no significant main effect of genotype, or a genotype x exercise interaction, for measures of body weight change or food intake. There were, however, main effects of exercise on body weight change [F(3,49) = 3.23, p = 0.0302]. As would be expected, general trends were seen with exercise mice losing weight. Post-hoc analyses, however, revealed that exercise (regardless of length of access) affected weight gain in WT mice (p < 0.05 for all), but not Tg2576 mice (Fig. 3A). Main effects of exercise were also observed in food intake [F(3,51) = 3.85, p = 0.0147]. Additionally, while all WT exercise groups had increased food intake compared to sedentary controls [p < 0.05 for all except 1 h (p = 0.06)], only 3 h Tg2576 mice ate significantly more than their sedentary (SED) counterparts (p < 0.05) (Fig. 3B). Our findings that exercising animals ate more and weighed less is intuitive and indicates that the exercise paradigms utilized were physiologically demanding.

Analysis of body weight and food intake over 4 months of exercise conditions. (Left) WT mice in exercise conditions lost more weight than their SED control counterparts. This effect is not seen in Tg2576 mice. (Right) While all WT exercise groups had increased food intake compared to SED controls only 3h Tg2576 mice ate more than their SED counterparts. *p < 0.05 versus sedentary (within that GT).
General activity, exploration, and motor competence
Rotarod, open field, and Y-maze
The rotarod, open field, and Y-maze (number of arm entries activity measure) are considered here in the context of the running and physiological analyses to examine the motor and temperamental effects of the exercise intervention. These non-mnemonic effects would reflect potentially beneficial effects on the exercise intervention on stamina, coordination, and willingness to explore novel environments (i.e., anxiety-like behavior). All of these measurements model clinically-relevant behavior in humans with functional importance to a person developing AD symptomology.
Two-way ANOVA showed that there was a main effect of genotype [F(1, 51) = 2.67, p = 0.002] on rotarod performance, such that Tg2576 mice were impaired compared to WT mice. Post-hoc analyses found that this trend was significant in 1 h mice only (p < 0.05; p = 0.0523 for 12 h). The main effect of exercise approached significance [F(3,51) = 2.67, p = 0.0573], with exercise mice showing improved performance compared to sedentary mice (p < 0.05 for all except 12 h p = 0.091). Post-hoc analyses found that exercise appears to be effective in WT mice, such that all exercise groups exhibited increased performance compared to the SED group (1 h p < 0.05, 3 h p = 0.068, 12 h p = 0.089). However, while similar trends of improved performance were seen in Tg2576 exercise groups, none of the comparisons reached significance (p > 0.20 for all) (Fig. 4A). These data are most readily interpreted as suggesting that exercise training enhances balance and coordination in the mice, and that this beneficial effect is not undermined by neuropathology in Tg2576 mice.
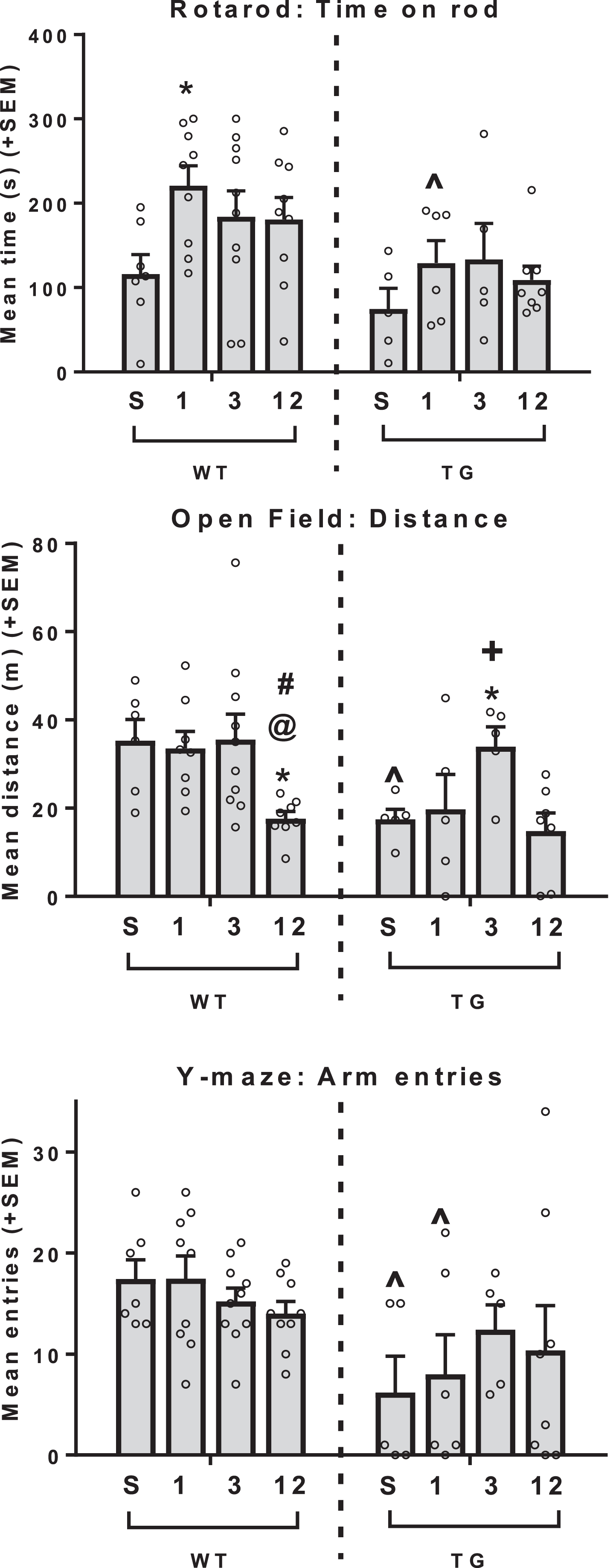
Analysis of rotarod performance as a measure of physical fitness and open field and Y-maze arm entries as measures of exploratory behavior. (Top) Tg2576 mice were impaired on the rotarod compared to WT mice. In WT mice, exercise increased performance in 1h mice. A similar trend can be seen in Tg2576 mice. (Middle) Tg2576 animals traveled shorter distance in the open field with WT and Tg2576 displaying different trends varying by exercise conditions. Within WTs, 12h mice were less active than all other groups, while in Tg2576 mice 3h mice were more active than all other groups. (Bottom) Overall, Tg2576 mice made fewer entries to the Y-maze. ∧p < 0.05 GT difference (within that exercise dose), *p < 0.05 versus sedentary (within that GT), @p < 0.05 versus 1h (within that GT), #p < 0.05 versus 3h (within that GT), +p < 0.05 versus 12h (within that GT).
Two-way ANOVA showed the main effect of genotype to be significant [F(1,46) = 6.778, p = 0.0124], with Tg2576 mice traveling shorter distances in the open field compared to WT mice. Post-hoc tests found that this difference was seen in SED (p < 0.05) and 1 h (p = 0.056) mice. The main effect of exercise was also significant [F(3,46) = 5.384, p = 0.0029]. Post-hoc tests revealed different trends of the various exercise conditions depending on genotype. Within WTs, 12 h mice were less active than all other groups (p < 0.05 for all), while in Tg2576 mice 3 h mice were more active than all other groups (p < 0.05 for all except 1 h p = 0.075) (Fig. 4B). For the measure of arm entries in the Y-maze, two-way ANOVA showed that the main effect of genotype was significant [F(1,51) = 11.91, p = 0.0011], such that overall Tg2576 mice made fewer arm entries compared to WT mice. Post hoc analyses showed that this comparison was significant within SED and 1 h (p < 0.05 for both) (Fig. 4C). The main effect of exercise and the genotype x exercise interaction were not significant. These latter two tasks both are both able to reveal a lower tendency for Tg2576 animals to actively explore novel environments compared to WT mice, and show some ability of exercise to restore activity levels closer to WT behavior.
Pathology
ELISAs for Aβ species
One-way ANOVAs were performed to determine the effect of exercise on forebrain levels of soluble and insoluble Aβ40 and Aβ42. Tg2576 3 h mice showed lower levels of soluble Aβ42 (p < 0.05; Fig. 5B) and insoluble Aβ42 (p = 0.07; Fig. 5D) compared to their SED counterparts, but the latter only approached significance. There were no group differences in soluble Aβ40 (Fig. 5A) or insoluble Aβ40 (Fig. 5C).

Analysis levels of Aβ species. A) No significant differences were observed between conditions in the levels of Aβ40S. B) Compared to Tg2576 SED mice, Tg2576 3h mice expressed lower levels of Aβ42S. C) No significant differences between conditions in the levels of Aβ40I. D) The difference between Aβ42I levels of Tg2576 SED mice and Tg2576 3h mice are approaching significance (p = 0.07), while Aβ42I levels were also significantly reduced in Tg2576 3h versus Tg2576 1h mice. *p < 0.05 versus sedentary (within that GT), @p < 0.05 versus 1h (within that GT).
Thioflavin-S staining of regional fibrillar amyloid deposition
Based on the results from the ELISAs, specific regional analyses were conducted to determine the effects of exercise on levels of amyloid deposition in the cortex and the hippocampus. There was no effect of exercise by one-way ANOVAs on levels of amyloid in the cortex (not shown). A main effect of exercise was approaching significance in the hippocampus [F(3,16) = 2.684, p = 0.0816]. Additionally, all exercise groups exhibited attenuated hippocampal plaque load compared to SED controls (p < 0.05 for all except 3 h p = 0.054; Fig. 6A). Although there were reductions in amyloid plaque load inspection of Thioflavin S stained plaques from the hippocampus did not reveal an appreciable change in the size or morphological features of the plaques in the Tg2576 exercising groups (Fig. 6B).
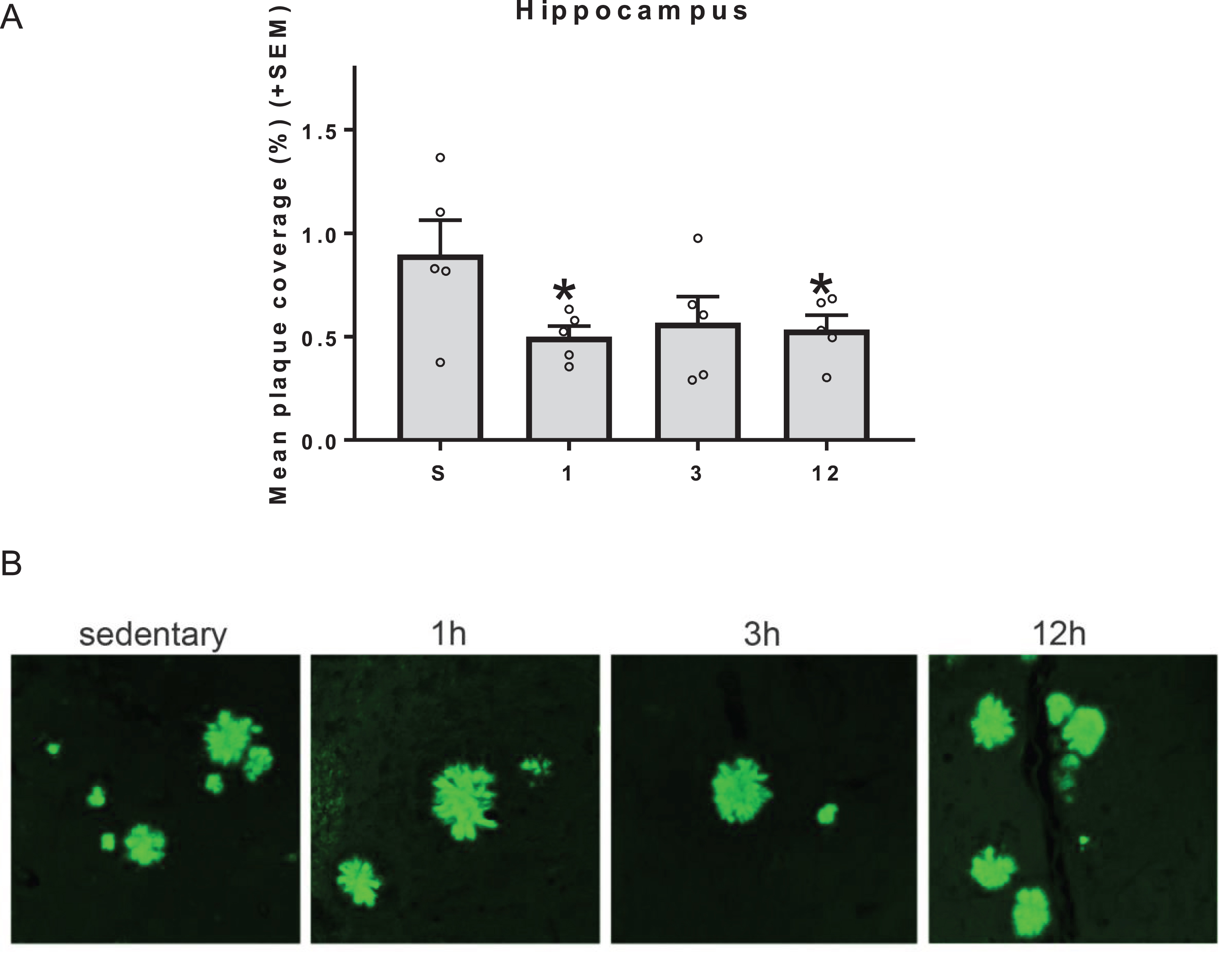
Amyloid plaque levels in the hippocampus. (Top) All exercise groups exhibited reduced hippocampal plaque load compared to SED controls (*p < 0.05 for all except 3h p = 0.054). Although the global effect was significant, there were no discernable differences in amyloid plaque size and morphology between the groups in very localized regions (Bottom).
ELISAs for Aβ species: Correlations between running parameters and Aβ species levels
It is interesting to determine which running characteristics (volume, speed, density) might best predict decreased amyloid. This is important in generalizing how much and what type of exercise may be most influential on Aβ levels. Therefore, we performed correlation analyses across all exercising Tg2576 mice to determine which running parameters were most associated with Aβ levels. These parameters were running volume as measured by total number of rotations performed per day averaged over 4 months, and intensity measured by running density (rotations/h) and running speed (rotations/min), averaged over 4 months.
Insoluble Aβ42: Pearson correlations showed a moderate, though not statistically significant, negative correlation between insoluble Aβ42 levels and running volume, r = –0.387, p = 0.077, such that mice with higher running volume had lower insoluble Aβ42 levels (Fig. 7A). A moderate negative correlation was also observed between insoluble Aβ42 levels and running speed, r = –0.471, p = 0.038, such that mice who ran at higher speed showed lower levels of insoluble Aβ42 (Fig. 7B). A moderate, non-significant negative correlation was observed between insoluble Aβ42 levels and running density, r = –0.421, p = 0.059, such that mice with higher densities showed lower levels of Aβ42 levels (Fig. 7C).
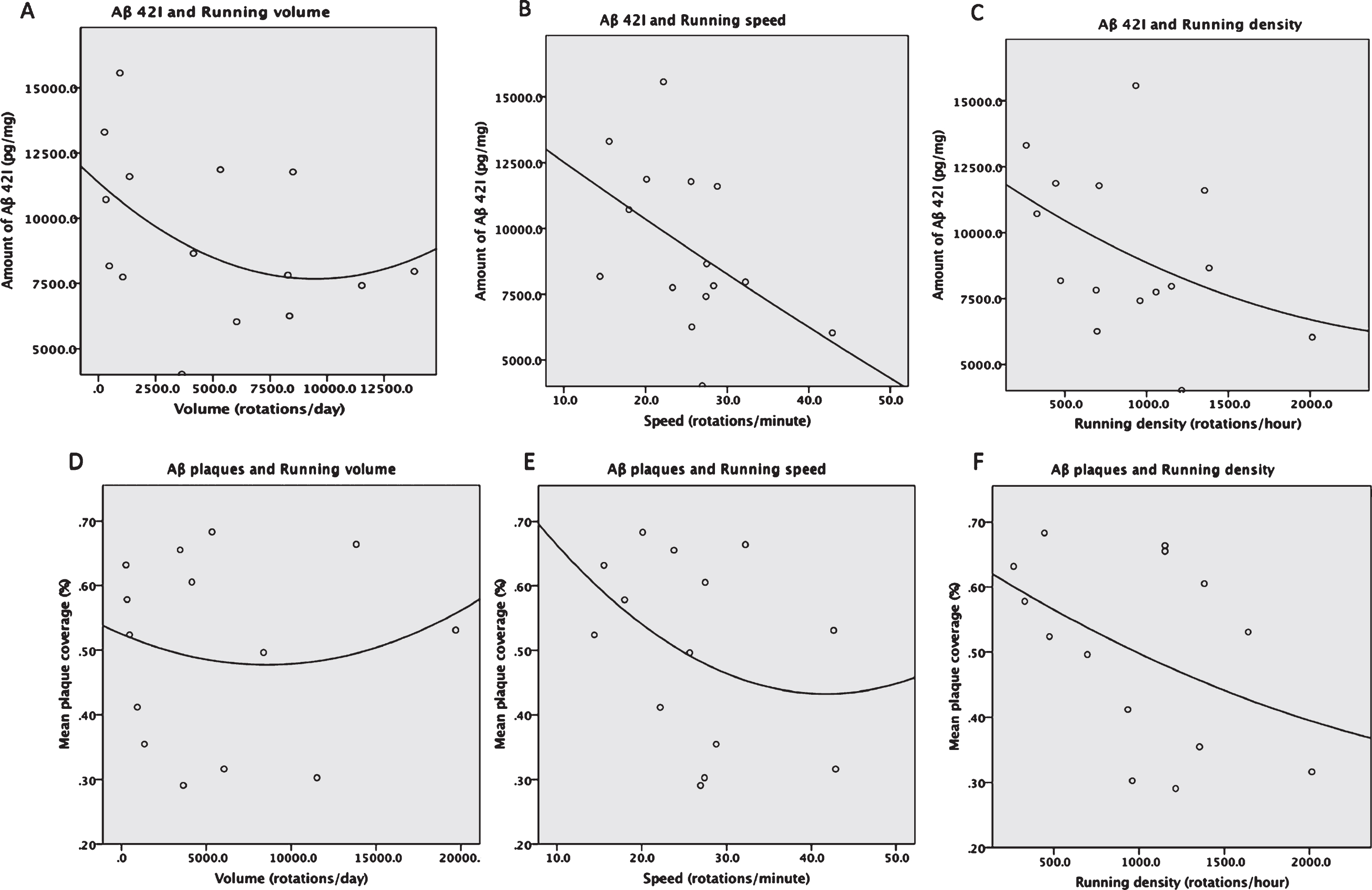
Nonlinear regression analysis between levels of running parameters and insoluble Aβ42 shows benefits of intensity measures and not volume. A) a non-significant moderate negative relationship between running volume and amount of insoluble Aβ42 was found (p = 0.07). B) A significant moderate negative correlation between running speed and amount of insoluble Aβ42 (p = 0.038) was found. C) A moderate negative correlation between running density and amount of insoluble Aβ42 was found (approaching significance p = 0.059). D) Nonlinear regression analyses between running parameters and Aβ plaques in the hippocampus show a very weak non-significant positive correlation between running volume and Aβ plaques, a moderate non-significant negative correlation between speed and Aβ plaques (E), and a moderate negative correlation between density and Aβ plaques that approached significance (p = 0.058) (F).
Other Aβ species: Correlations were also performed for assessing the relationship between running measures and other Aβ species (insoluble Aβ40 and soluble Aβ40 and Aβ42). While these showed a general trend of moderate to low negative correlations, these correlations were not significant (data not shown).
Thioflavin-S staining of regional fibrillar amyloid deposition: Correlations between running parameters and Aβ plaque coverage
Pearson correlations were performed across all exercising Tg2576 mice, as was done with ELISAs, to determine which running parameters were associated with Aβ plaque coverage in the hippocampus and cortex. These parameters include volume measured by total number of wheel rotations per day averaged over 4 months and intensity measured by running density (rotations/h) and running speed (rotations/min) averaged over 4 months.
Hippocampus Aβ plaques: There was no relationship between percent area coverage of Aβ plaques in the hippocampus and running volume, r = 0.017, p = 0.477 (Fig. 7D). A moderate non-significant negative correlation was found between percent area coverage of Aβ plaques in the hippocampus and running speed, r = –0.352, p = 0.109 (Fig. 7E). A moderate negative correlation was also found between percent area coverage of Aβ plaques in the hippocampus and running density that approaching significance, r = –0.440, p = 0.058, such that mice with higher densities showed lower levels of hippocampus Aβ plaques (Fig. 7F). Taken together, these findings suggest that running intensity, rather than volume, may be more predictive of amyloid pathology.
Spatial learning and memory
Barnes maze
The observation of exercise-associated reduced plaque load in the hippocampus raises the question as to whether performance in the Barnes maze, a spatial learning and memory task known to be sensitive to hippocampal damage, would be seen to be improved relative to non-exercising Tg2576 mice. A three-way repeated measures ANOVA was performed to determine group differences in performance on the Barnes maze over the 5 days of testing, as measured by escape latency. There was a main effect of genotype [F(1,51) = 5.486, p = 0.0231], such that Tg2576 mice exhibited a longer latency compared to WT mice (p < 0.05). The main effect of time was also significant [F(4, 204) = 25.676, p < 0.001), such that mice got progressively faster at entering the escape whole on each subsequent test day (p < 0.05 for all) (Fig. 8A). All WT groups showed a non-zero learning curve on the Barnes maze, suggesting intact spatial learning (p < 0.05 for all; Fig. 8B). Tg2576 SED mice did not exhibit a learning curve with a slope significantly different from 0, suggesting impairment on this task. Exercise appeared to “rescue” this deficit in Tg2576 mice, such that all Tg2576 exercise groups showed a non-zero learning curve (p < 0.05 for all). Slopes of the learning curves were then compared between all treatment groups. There was a significant main effect of genotype [F(1,51) = 7.45, p = 0.009], with Tg2576 mice showing a less steep learning curve compared to WT mice overall (p < 0.05), and within SED mice and 3 h mice as shown by post-hoc test (p < 0.05). The main effect of exercise was also significant [F(3,51) = 2.803, p = 0.049], with 3 h and 12 h mice showing a steeper learning curve compared to sedentary controls (p < 0.05). Post-hoc analyses revealed that within WT mice, only 3 h mice had a steeper slope compared to sedentary mice. In Tg2576 mice, the slope of the learning curve tended to be greater in all Tg2576 exercise mice compared to Tg2576 SED mice (p = 0.06 versus 1 h; p = 0.09 versus 3 h and 12 h), again suggesting a modest “rescue” effect of exercise on the Barnes maze task.
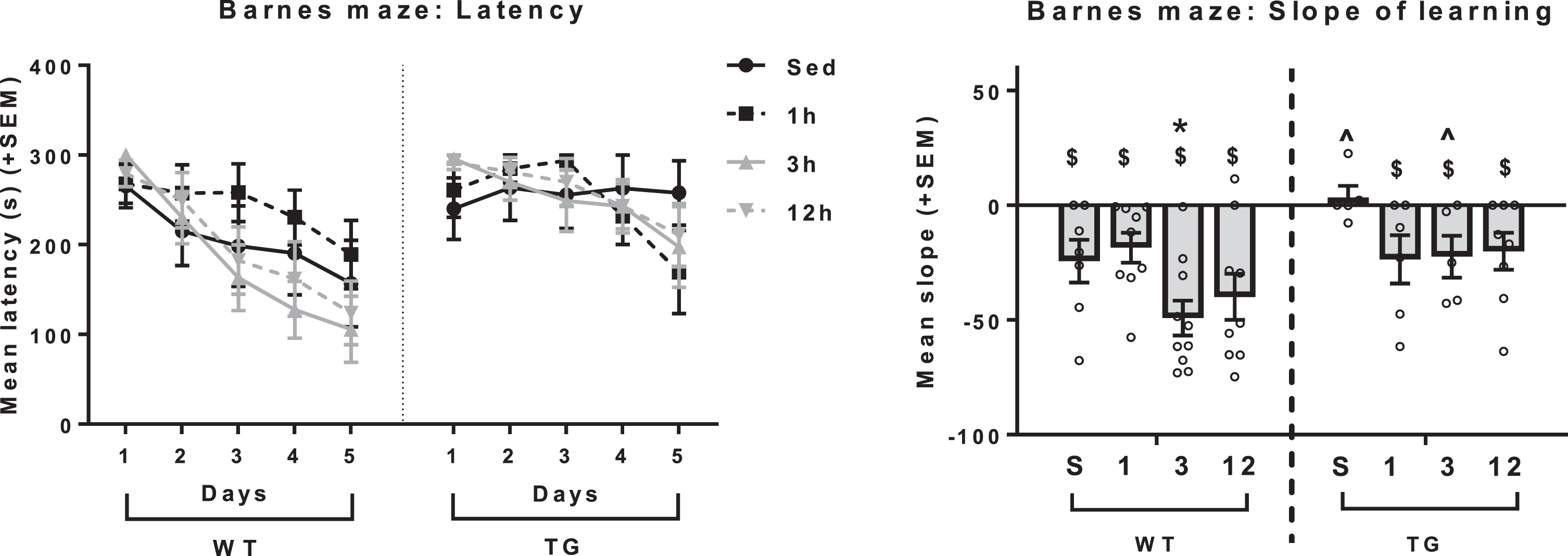
Barnes maze as a measure of spatial learning. (Left) Tg2576 mice exhibited longer latency to enter the escape box compared to WT mice. (Right) All WT groups showed a non-zero learning curve on the Barnes maze. Tg2576 SED mice did not exhibit a learning curve with a slope significantly different from 0. All Tg2576 exercising groups also showed a non-zero learning curve (p < 0.05 for all). Within WT mice, only 3h showed a steeper slope compared to WT SED. $non-zero learning slope. ∧GT difference (within that exercise dose), *p < 0.05 versus sedentary (within that GT).
Other effects of exercise on behavior
The main effect of genotype was significant for digging time [F(1,48) = 10.23, p = 0.0024], such that WT mice dug more than their Tg2576 counterparts, while exercise had no effect on this measure (data not shown). No significant differences between genotype or exercise were found in other measures or behavioral tasks performed such as object displacement, novel object recognition, social interaction, and light-dark box (data not shown).
Summary of results
Providing 1, 3, and 12 h of access to a running wheel resulted in exercise regimens that varied by volume and intensity and WT and Tg2576 mice in these groups exhibited similar exercise patterns. Exercise appeared to exert greater metabolic effects on WT as compared to Tg2576 mice and may have increased survival. While WT and Tg2576 mice performed similarly on many behavioral tasks, some deficits in Tg2576 mice were seen in measures of motor activity and spatial learning and memory. Exercise “rescued” many of these deficits while also improving performance in WT mice. Additionally, while all exercise regimens reduced hippocampal Aβ plaque load in Tg2576 mice, the highest intensity 3 h regimen appeared to be most effective in reducing whole forebrain levels of Aβ42 (soluble and insoluble). The beneficial effects of higher intensity exercise on amyloid pathology were also supported by correlation results.
DISCUSSION
Previous work has suggested that regular, sustained exercise in humans and animals is beneficial for brain health and functioning and may prevent and alleviate symptoms of brain disorders, such as AD [10, 30]. Rodent models of AD, which are useful for revealing the causal mechanisms underlying these effects, have generally, though not unanimously, confirmed the positive effects of exercise [11, 32]. Despite findings that suggest that cardiovascular exercise may be preventative or restorative for AD symptomology, many important questions about exercise that can inform human health choices and clinical interventions have not been explored. Critically, long-term exercise interventions in older mice provide an opportunity to address whether well-established AD pathologies are susceptible to remediation. Furthermore, how much and how intense the exercise needs to be to yield benefits is also important to determine, especially given the inherent difficulties and risks of higher intensity exercise interventions in previously inactive older adults. Most interventions that target older adult humans, especially those who have not been previously active, are relatively low-volume and low intensity as compared to higher volume and intensity conditions [33, 34]. Finally, it is important to know whether, in addition to cognitive benefits, general benefits of exercise such as improved mood or better coordination, balance and endurance are diminished by more advanced AD pathology. Therefore, our study assessed these questions in older Tg2576 mice using exercise regimens that varied in volume and measures of intensity.
Different access periods to running wheels lead to varying running qualities
Previous work in both animals and humans has suggested that there is a positive association between frequency and duration of cardiovascular exercise and cognitive function; however, some studies have proposed that only minimal amounts of exercise can produce significant health benefits or that even short duration, high-intensity regimens can produce benefits with regard to cardiovascular and metabolic function, musculoskeletal benefits and cognitive performance comparable to higher volume regimens [35–41]. Because we used a voluntary exercise intervention, the different patterns of running and breaks created the exercise volume and intensity. In the current study, the 1 h mice exhibited an exercise regimen of the lowest volume and low intensity. Therefore, this group can model an intervention in humans of low to moderate intensity and limited duration (e.g., an hour of vigorous walking). The 3 h group ran a moderate volume at the highest intensity, while the 12 h group ran the highest volume at moderate intensity. The actual measures of volume and intensity were used for analysis of the effects of exercise on AD pathology. It is important to note that WT and Tg2576 mice exhibited similar running patterns in regards to volume and intensity within each of the three exercise groups, indicating that pathology exhibited by Tg2576 mice did not impair their ability to exercise into older age as compared to controls. Additionally, this indicates that any differences between WT and Tg2576 mice within exercise groups cannot be attributed to genotype differences in exercise volume or intensity.
Older Tg2576 exercising mice exhibit improvements in Aβ pathology
When the levels of Aβ species were analyzed, it was found that Tg2576 3 h mice expressed lower levels of soluble Aβ42 compared to Tg2576 SED group. Though this pattern was only approaching significance in insoluble Aβ42, Tg2576 1 h and 3 h group were significantly different. Similarly, in the hippocampus, Tg2576 SED mice expressed higher levels of Aβ than all exercising groups (though the 3 h group was only approaching significance). Additionally, when the exercise volume and intensity parameters were correlated with Aβ species, it was found that reduced insoluble Aβ42 was associated with intensity measures (higher speed and density). No other Aβ species were associated with any of the running parameters. Taken together, we see some positive effects of exercise in the formation or clearance of amyloid pathology and these positive effects seem to occur more often in the higher intensity conditions.
Studies have pointed to different roles of soluble and insoluble species of Aβ. It has been suggested that insoluble Aβ plaques can be a mechanism for soluble Aβ removal [42]. Other studies have suggested that a progressive shift from soluble to insoluble Aβ plays role in the progression of AD [43]. While the specific mechanism by which insoluble Aβ42 would be selectively decreased by exercise is not discerned presently, both ELISA and histology results support that exercise intensity, rather than volume, may be a better predictor of amyloid pathology.
Exercise improved some behavior measures, including spatial learning and memory in older WT and Tg2576 mice
In general, we see Tg2576 mice performing less proficiently in the Barnes maze task as compared to their WT counterparts as expected, as spatial memory impairment in Tg2576 mice is well established by past studies using various tasks [44–47]. Specifically, we found that sedentary Tg2576 mice failed to show a non-zero slope of learning in the Barnes maze (indicating a lack of spatial learning and memory) that was exhibited by all WT groups. This was rescued by exercise, such that all Tg2576 exercise groups exhibited a non-zero learning slope similar in magnitude to WT controls. This suggests that exercise improved spatial learning and memory in Tg2576 mice at an age associated with advanced-stage disease. This effect is consistent with the pathological finding that all exercising groups expressed reduced Aβ plaques in the hippocampus, a region widely accepted for its role in spatial learning and memory [48]. Similarly, exercise dose-dependently improved Barnes maze performance in WT mice, specifically in the 3 h group, suggesting that higher intensity exercise may also be effective in boosting hippocampal-dependent memory in the healthy aging population.
Object displacement, novel object recognition, social interaction, and anxiety-like behavior (light-dark box performance) were similar between WT and Tg2576. These measures were also unaffected by exercise in both genotypes. We have previously reported that these measures are improved by the same exercise regimens in WT and another amyloid model of AD (TgSwDI) though these mice were tested at a younger age (12 months) [20, 21]. It is possible that at this advanced age of 18 months, exercise was no longer capable of exerting similar behavioral benefits. It also appears that exercise had similarly positive effects on some measures of motor competence (rotarod) in both genotypes, though the effects of exercise varied by genotype on other tasks related to motor and exploratory behavior. Specifically, exercise exerted a “rescue” effect on Y-maze arm entries in Tg2576 mice, while exercise had no effect in WT mice. This is in agreement with our previous findings using the same exercise regimens in one-year old WT and TgSwDI mice [20, 21]. Additionally, exercise exerted different effects in the open field in WT and Tg2576 mice. In Tg2576 mice, the highest intensity (3 h) exercise group also showed “rescued” behavior, as demonstrated by increased activity in the open field. This difference could signify an enhancement in exploration rather than anxiety levels since the open field test (center activity) and light-dark box task did not reveal any differences between genotypes or exercise conditions. A dose-dependent effect of exercise on open field behavior was also previously shown in TgSwDI mice [20]. In contrast to our prior study that found no effect of exercise in 12-month-old WT mice [21], the highest volume of exercise (12 h) in WT mice used in the current study decreased locomotor behavior to the level of Tg2576 mice. However, this may not suggest exercise to be detrimental in WT mice, as other tasks did not show similar patterns. Taken together, these findings suggest that the effects of exercise on behavior may be dose-, strain-, and age-dependent.
Limitations
One limitation of this study, as with any study using aged subjects, was the loss of some animals toward the end of the performance period. While minimal, this had potential to skew the results by introducing survivor bias. Future studies utilizing aged Tg2576 mice should consider using a larger sample size in anticipation of this loss, perhaps focusing on a high intensity condition. Follow-up work could also utilize treadmill running and perhaps acrobatic exercise interventions to establish the generality of the exercise effects observed presently. Finally, other processes that are influenced by exercise changes such as cerebral blood flow, neuroinflammation, hippocampal growth factors, angiogenesis, and neurogenesis could be examined as to whether they play a specific beneficial role in aged subjects [20, 50].
Conclusions
Overall, our study suggests that some Aβ levels can be diminished and behaviors can be improved through an extended period of cardiovascular exercise even in older animals with advanced pathology. Differences in Aβ species levels show greater reduction in the exercise group that ran at the highest intensity. Moreover, the intensity of the exercise, rather than the volume, was better correlated with reduced insoluble Aβ42 and deposited Aβ plaque areas in the hippocampus. The behavioral improvements could be due directly to these lowered levels of pathology observed in the exercising Tg2576 mice compared to SED Tg2576 mice or to an indirect action on other aspects of brain function or even somatic effects. These include reduced neuroinflammation, reduced oxidative stress, increased cerebral blood flow or neurogenesis, all of which that have been implicated in the effects of exercise. The various improvements across the behavioral paradigms did not show a consistent pattern of benefit across the three access groups, showing the greatest benefit sometimes in the 3 h group or all exercising groups. This may be best interpreted as the result of effects to indirect or multiple mechanisms. Translating these results to humans suggests that exercise, especially, higher intensities of exercise in individuals with emerging AD may have beneficial effects on reducing some Aβ pathology, and that people may be able to see benefits of exercise at multiple levels even if previously sedentary, and with established pathology and existing cognitive impairments.
Footnotes
ACKNOWLEDGMENTS
This work was supported by NIH grant AG048935 to J.K.R., B.J.A., and W.E.V. as multiple PIs. Antibody reagents for the Aβ ELISAs were generously provided by Eli Lilly (Indianapolis, IN). The authors would like to thank Sarima Subzwari, Regina Kim, Steven I. Beigelman, Shannon M. Fitzgerald, Joseph Arena, Rachel Rock, David A. Lituma, Sarah Amrein, Antonina E. Kuzmina, Dr. Michael Hoos, Ralph Molaro, Everett Waters, and Robert Chorley for their technical assistance.