
Abstract
Alzheimer’s disease (AD) is neuropathologically defined by two key hallmarks: extracellular senile plaques composed primarily of amyloid-β (Aβ) peptide and intraneuronal neurofibrillary tangles, containing abnormally hyperphosphorylated tau protein. The tau protein is encoded by the MAPT gene. Recently, the H1 and H2 haplotypes of the MAPT gene were associated with AD risk. The minor MAPT H2 haplotype has been linked with a decreased risk of developing late-onset AD (LOAD). MAPT haplotypes show different levels of MAPT/Tau expression with H1 being ∼1.5-fold more expressed than H2, suggesting that MAPT expression level could be related to LOAD risk. In this study, we investigated whether this moderate difference in MAPT/Tau expression could influence Aβ-induced toxicity in vivo. We show that modest overexpression of tau protein in Drosophila exacerbates neuronal phenotypes in AβPP/BACE1 flies. The exacerbation of neuronal defects correlates with the accumulation of insoluble dTau oligomers, suggesting that the moderate difference in level of tau expression observed between H1 and H2 haplotypes could influence Aβ toxicity through the production of oligomeric tau insoluble species.
INTRODUCTION
Alzheimer’s disease (AD) is the most common cause of dementia worldwide. Clinically, AD is characterized by progressive memory loss and behavioral disorders, associated with cerebral atrophy and synaptic loss [1]. Neuropathologically, AD is defined by two key hallmarks: extracellular senile plaques composed primarily of amyloid-β (Aβ) peptide and intraneuronal neurofibrillary tangles, containing abnormally hyperphosphorylated tau protein.
The genetics of AD is heterogeneous. Some families with autosomal dominant early-onset AD harbor extremely rare mutations of APP, PSEN1, and PSEN2 genes, which are directly linked to the production and/or aggregation of the Aβ peptide [2]. In the majority of AD cases, the determinism is complex. Genetic risk factors include the strong and common APOE ɛ4 allele, ∼25 common loci with a modest effect, and rare variants in TREM2, SORL1, and ABCA7 that are considered as moderate-to-strong risk factors (for review, see [3]). Most of these factors have been shown to interfere with Aβ production, clearance, aggregation, or toxicity [4, 5].
MAPT haplotypes have also been associated with AD risk. The MAPT gene, which encodes tau protein, resides within a ∼900 kb inversion polymorphism that generates a ∼1.3 Mb region of linkage disequilibrium defined by two extended haplotypes termed H1 and H2. The H1 haplotype clade is the most common, having an allele frequency of about 70% in European populations and a greater frequency in other populations [6–8]. The minor H2 haplotype has been associated with a decreased risk of developing late-onset AD (LOAD) (odds ratio, OR = 0.90, 95% CI = 0.85–0.95) [6, 7]. Several mechanisms could explain this effect. First, there is evidence that alternative splicing events of MAPT mRNA are under haplotype-specific control. The protective MAPT H2 haplotype significantly expresses less exon 10 + MAPT mRNA [9, 10] and more exons 2 + 3+ MAPT [11, 12] transcripts than the H1 haplotype. Second, MAPT haplotypes show different levels of MAPT expression [6, 13–15], with H1 haplotype being ∼1.5-fold more expressed than H2 haplotype [16], suggesting that MAPT expression level could influence LOAD risk.
Several studies established that tau and Aβ cooperate to induce AD pathology [17]. On the one hand, tau depletion protects against Aβ-associated cognitive deficits, synaptic loss, and neuron death in mouse [18–21]. Conversely, wild-type tau overexpression leads to synergistic effects on Aβ pathology, with exacerbation of cognitive defects, synapse loss, and memory impairment [22, 23]. However, all these animal and cellular models rely on massive overexpression of the human wild-type tau protein. To our knowledge, the effect of a moderate increase in tau expression on Aβ-induced toxicity has not been reported so far.
In this study, we focused on the question of whether MAPT expression level could influence LOAD risk. We investigated if a difference in MAPT expression level similar to that observed between H1 and H2 haplotypes could influence Aβ-induced toxicity in vivo. We used two Drosophila models: one expressing the Drosophila tau (dTau) protein ∼1.6-fold over endogenous level [24] and a second co-expressing the human wild-type AβPP and BACE1 transgenes [25]. In the latter model, neuronal co-expression of AβPP and BACE1 in flies results in production of Aβ40 and Aβ42 peptides, abnormal accumulation of Aβ in fly brain, wing expansion defects, memory and locomotor deficits [25], and synaptic defects [26]. Importantly, these phenotypes depend on AβPP amyloidogenic processing, as feeding flies with a γ-secretase inhibitor significantly reduces Aβ load in fly brain and rescues neuronal defects and locomotor deficits.
MATERIAL AND METHODS
Fly genetics
Drosophila were maintained on a 12 : 12 h light/dark cycle on standard cornmeal-yeast agar medium at 25°C. The following transgenic Drosophila strains were used in the study: UAS- AβPP; UAS-BACE1 [25], UAS-dTau::1D4 [24]. The elavC155-Gal4 line was obtained from Bloomington Stock Center (ID458).
Wing defects analysis
More than 1,000 flies per genotype were analyzed for unexpanded wing phenotype one day after hatching with a binocular stereomicroscope Leica M80 at 7.5 magnification (Leica Biosystems, Wetzlar, Germany).
Climbing assay
Flies were collected in tubes in groups of 10 on the day of hatching. They were analyzed for locomotor deficits the day after. The climbing assay consisted in gently tapping each tube three times to make the flies fall to the bottom of the tube and then these flies were given 10 s to climb to the top of the tube. Each tube was processed three times. The number of flies that successfully reached the upper half of the tube was counted. More than 200 flies per genotype were assessed in a total of three independent experiments.
Real-time quantitative RT–PCR
Total RNA extraction was performed on 30 newly eclosed adult fly heads using the Nucleospin RNA II kit (Macherey-Nagel GmbH and Co. KG, Düren, Germany) according to the manufacturer’s instructions. The amount of RNA per μL was measured using a NanoDrop Lite (Thermo Fisher Scientific Inc.). 1 mg of total RNA was reverse-transcribed into cDNA, using the RETROscript Reverse Transcription kit (Thermo Fisher Scientific Inc.) with Oligo(dT) primers according to the manufacturer’s instructions for 2-Step RT-PCR without heat denaturation. PCR reactions were performed in a final volume of 20 μL, using the SsoFast Evagreen Supermix (Bio-Rad Laboratories, Hercules, CA, USA) with primers at a final concentration of 250 nM (dTau: forward 5’-AACCGCTTGCTCCTCAAGAC-3’ and reverse 5’-TGGGTGAAGGCGCATGT-3’; RpL13A: forward 5’-AGCTGAACCTCTCGGGACAC-3’ and reverse 5’-TGCCTCGGACTGCCTTGTAG-3’). PCR amplifications were performed on a CFX96 Real-Time System thermal cycler (Bio-Rad Laboratories) using the following cycling steps: enzyme activation at 98°C for 2 min; denaturation and annealing/extension, respectively, at 98°C for 10 s and 60°C for 15 s (40 cycles). The comparative –ΔΔCt method was then used to determine quantitative values for gene expression levels in each sample using RpL13A as the housekeeping gene.
Protein extraction
For sequential extraction of soluble and insoluble proteins, 40 adult fly heads were dissected from newly eclosed adults and crushed with a pestle on ice in 150 μL Radio Immunoprecipitation Assay (RIPA) buffer (25 mM Tris–HCl pH 7.6, 150 mM NaCl, 1% NP-40, 1% sodium deoxycholate, 0.1% SDS) (Pierce Biotechnology, Rockford, IL, USA), supplemented with a cocktail of protease inhibitors (Sigma-Aldrich). Lysates were then centrifuged at 11,300 g for 20 min at 4°C. The supernatant, which contains the RIPA-soluble fraction of proteins, was reserved in a separate tube while the pellet was washed once in 75 μL of RIPA and centrifuged under the same conditions and the resulting supernatant was pooled with the first one. The remaining pellet was then homogenized in 225 μL of SDS-PAGE sample buffer (80 mM Tris–HCl, pH 6.8, 2% SDS, 10% glycerol, and 0.02% bromophenol blue) and centrifuged at 11,300 g for 30 min. The supernatant was collected as the RIPA-insoluble fraction. Protein concentrations of the soluble fractions were measured using the DC Protein Assay Kit (Bio-Rad Laboratories). Soluble and insoluble proteins were loaded for SDS-PAGE migration in a proportion of 1 : 2. For total protein extraction, dissected fly heads were directly homogenized in SDS-PAGE sample buffer.
Immunoblot analysis
Proteins were resolved by TGX Stain-Free 10% gels (Bio-Rad Laboratories), and then transferred onto nitrocellulose membrane using the Trans-Blot Turbo system (Bio-Rad Laboratories). After blocking the membrane with 5% non-fat milk, it was incubated with appropriate antibodies. Gel loading was normalized by Stain-Free detection of total proteins using a Geldoc TM EZ imager (Bio-Rad Laboratories). The Stain-Free signal obtained in each lane was quantified (ImageLab TM software, Bio-Rad Laboratories). The following antibodies were used: anti-Drosophila tau 13114 [27] (1 : 3,000), anti-Drosophila tau SK6427 [28] (1 : 10,000), peroxidase-labeled anti-rabbit antibodies (1 : 10,000) from Jackson Immunoresearch Laboratories (WestGrove, PA, USA). Signals were detected with chemiluminescence reagents (ECL Clarity, Bio-Rad Laboratories) and acquired with a GBOX (Syngene, Cambridge, UK), monitored by the GeneSnap software (Syngene). The signal intensity in each lane was quantified using the Genetools software (Syngene), and normalized with the Stain-Free signal quantified in the corresponding lane.
Quantification of Aβ peptide
Total Aβ was isolated as previously described in [29] with minor modifications. Briefly, 300 fly heads were homogenized with a pestle in 1.5 mL protein low binding tube containing 100 μL of extraction buffer (50 mM HEPES, 5 M Guanidinium chloride, 5 mM EDTA). After homogenization, the samples were plunged for 5 min in a sonic bath. The homogenates were then centrifuged at 15,000 g for 5 min, the supernatant was collected and protein concentration was measured using Protein Assay Dye Reagent (Bio-Rad Laboratories). The quantification of Aβ peptides were performed using the V-PLEX Aβ Peptide Panel 1 (6E10) kit (Meso Scale Diagnostics, Rockville, MD, USA) according to manufacturer’s instructions. Protein extracts were diluted 1 : 25 in Diluent 35. The plate was analyzed using a MESO QuickPlex SQ 120 instrument (Meso Scale Diagnostics). Aβ quantities were normalized with total protein amounts measured in each sample.
Statistical analysis
Statistical analyses were performed using either a two-tailed Student’s t-test with Welch’s correction (comparison between two groups) or a one-way Analysis of Variance (ANOVA) and Tukey’s post-test (multiple comparisons). The statistical methods used are specified in each corresponding figure legend. Statistical analyses were performed with GraphPad Prism software (GraphPad, San Diego, CA, USA). All n reported are biological replicates. Statistical significance was defined as ns (not significant) p > 0.05, *p < 0.05, **p < 0.01, ***p < 0.001. All data are presented as mean±standard error of the mean (SEM).
RESULTS
Moderate increase in dTau expression level exacerbates neuronal phenotypes in AβPP/BACE1 Drosophila
To mimic in flies the moderate difference in MAPT expression level observed between H1 and H2 haplotypes in human brains, we used transgenic flies that express a 1D4-tagged dTau protein [24]. We crossed randomly inserted UAS-dTau lines with the elavC155-Gal4 driver line to express exogenous dTau in all post-mitotic neurons of the central and peripheral nervous system. Total RNA extraction from fly heads and qRT-PCR analysis allowed us to select two independent Drosophila lines displaying a ∼1.6-fold (Fig. 1, p = 0.0016) or ∼3.5-fold (Supplementary Figure 1, p = 0.0042) increase in dTau mRNA steady-state level compared to control flies.
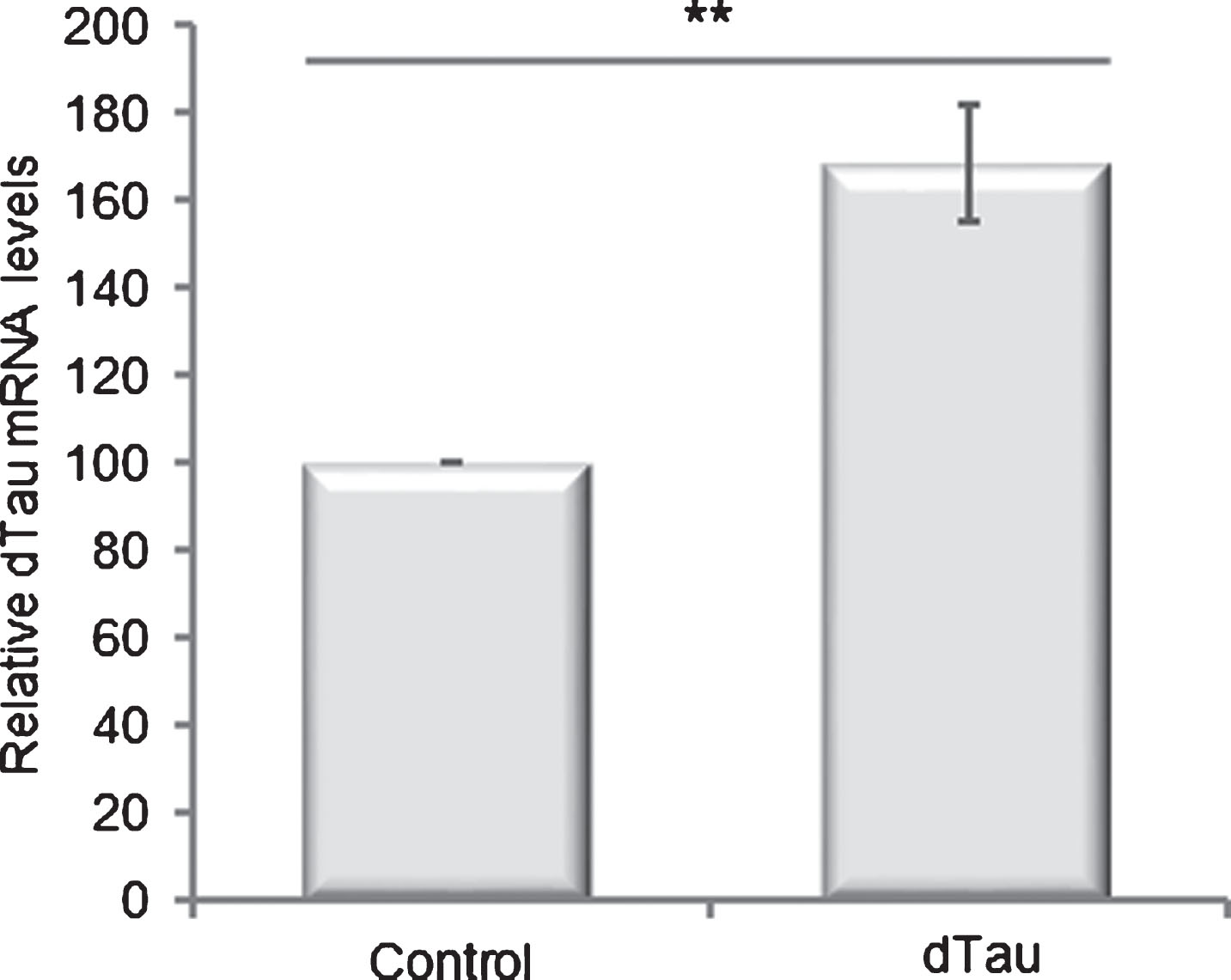
Selection of a Drosophila line with moderate overexpression of dTau transgene. Quantification of dTau mRNA levels by RT-qPCR in total mRNA extracted from 1-day-old transgenic flies expressing or not dTau transgene under the control of the elavC155-Gal4 driver line. Control flies: elavC155-Gal4> +. Controls were arbitrarily set at 100. Bar graph: mean±standard error of the mean (SEM) of relative dTau mRNA levels. **p < 0.01. p-values were calculated using a Student’s t-test (n = 8). dTau overexpression results in a modest but significant (p = 0.0016) increase in dTau mRNA steady-state levels.
To assess the effect of a moderate increase in dTau expression level on Aβ toxicity, we crossed these dTau flies with AβPP/BACE1 flies [25], and used firstly the wing expansion defect phenotype as a read-out. Wing expansion is a post-eclosion event regulated by the secretion of the insect neurohormone bursicon by a subset of neurons in the ventral nerve cord [30–32]. As reported by others [25], AβPP/BACE1 co-expression led to wing expansion defects in approximately 14.1% of newborn flies (Fig. 2A). A ∼1.6-fold increase in dTau expression level resulted in a significant increase in the rate of flies displaying wing abnormalities (27.6%; p = 3.6 10–8). Note that control flies and flies expressing dTau alone did not show wing expansion defects. Similarly, a 3.5-fold increase in dTau expression level also resulted in a marked increase in the rate of flies displaying a wing expansion defect phenotype (Supplementary Figure 1). The fact that similar data were obtained using these two independent transgenic lines ruled out the effect of integration site, and indicated that this genetic interaction is dependent on dTau expression.
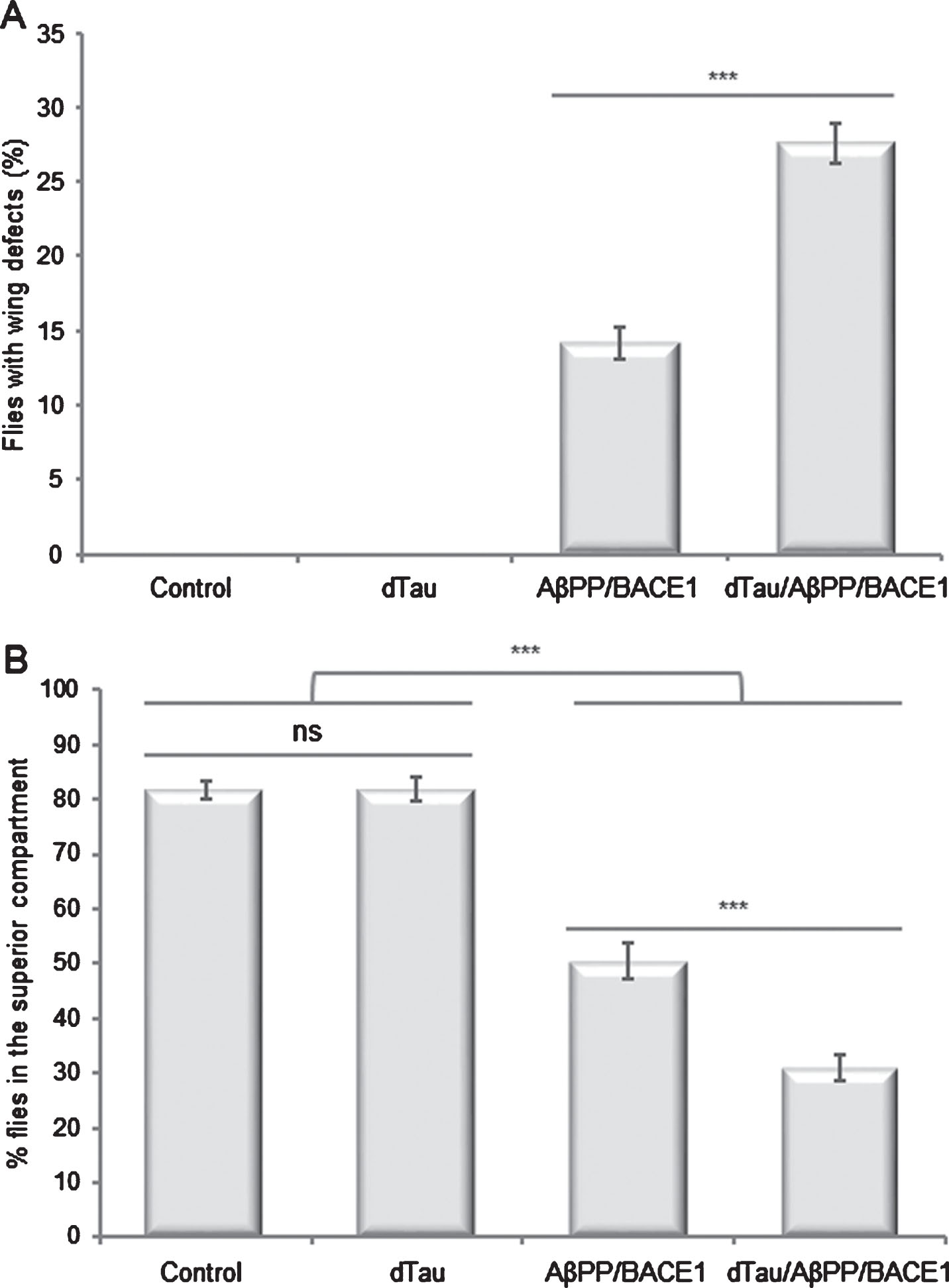
Moderate overexpression of dTau exacerbates AβPP/BACE1-induced neuronal phenotypes in Drosophila. Quantification of wing expansion defects and climbing abilities in control flies (elavC155-Gal4> +), flies overexpressing dTau (elavC155-Gal4 > UAS-dTau), AβPP and BACE1 (elavC155-Gal4 > UAS-AβPP; UAS-BACE1) or dTau, AβPP and BACE1 (elavC155-Gal4 > UAS-dTau, UAS-AβPP; UAS-BACE1). A) Quantification of flies presenting unexpanded wings the day following hatching. Wing expansion defects were observed in 14.1% of AβPP/BACE1 flies. Combined expression of dTau with AβPP and BACE1 resulted in a significant increase (27.6%; p = 3.6 10–8) in the rate of flies displaying wing abnormalities. Note that wing expansion defects were absent in control and dTau flies. Bar graph: mean±standard error of the mean (SEM). **p < 0.01, n > 1,000. p-values were calculated using a Student’s t-test. B) Climbing abilities of flies the day following hatching. One-way analysis of variance (ANOVA) and Tukey’s post hoc analyses were performed to determine the differences in climbing abilities between genotypes (ANOVA F (3,32) = 104.4, p < 0.0001, n = 3). dTau flies displayed similar climbing abilities as controls (control: 81.8%; dTau: 81.9%; p > 0.05). In contrast, severe climbing deficits were observed in AβPP/BACE1 flies (50.5%; p < 0.0001). Moderate overexpression of dTau combined with AβPP and BACE1 potentiated the climbing deficits (31.1%; p < 0.0001). Bar graph: mean±standard error of the mean (SEM). Ns, not significant p > 0.05; ***p < 0.001.
To validate this genetic interaction with an independent phenotype, we next assessed the impact of a moderate increase in dTau expression level on the locomotor abilities of these flies. Because the Drosophila line displaying a ∼1.6-fold increase in dTau mRNA steady-state level is the more relevant to mimic the difference in MAPT expression level observed between H1 and H2 haplotypes in human brains, we then decided to focus on this line for the following analyses. Briefly, flies display a negative geotaxis reflex when they are subjected to a mechanical stimulus. When tapped to the bottom of a tube, flies quickly climb to the top of the tube. Using a climbing assay, we measured the ability of newborn flies to climb to the upper half of the tube, 10 s after a mechanical stimulus and compared genotype performances using ANOVA. As expected, most of the control flies climbed to the top of the tube, with 81.8% of flies reaching the upper compartment (Fig. 2B). Increase in dTau expression did not alter the climbing abilities of the flies (81.9%; p > 0.9999), compared to control flies. In contrast, as reported by others [25], AβPP/BACE1 co-expression resulted in a significant decrease in climbing performances, with only 50.5% of the flies able to reach the upper half of the tube (p < 0.0001). Combined expression of dTau with AβPP and BACE1 exacerbated the climbing deficits with only 31.1% of flies reaching the upper compartment (p < 0.0001).
Altogether, these data indicate that a ∼1.6-fold increase in dTau expression level exacerbates neuronal phenotypes in AβPP/BACE1 Drosophila.
Moderate increase in dTau expression does not change the Aβ peptide steady-state level
To determine the mechanisms associated with the exacerbation of neuronal phenotypes in AβPP/BACE1 flies with a ∼1.6-fold increase in dTau expression level, we first tested whether dTau expression modulated the accumulation level of Aβ peptides in our experimental model. We quantified Aβ peptides using the ELISA MSD technology. This technology allows the parallel quantification of Aβ38, Aβ40, and Aβ42 peptides in a single assay. After normalization with total protein amounts, we detected 768.2 pg of Aβ40 and 466.1 pg of Aβ42/mg of total proteins in AβPP/BACE1 flies (Table 1). Expression of dTau in this genetic context did not modify the accumulation level of either Aβ peptide (783.0 pg of Aβ40/mg of total proteins; 472.0 pg of Aβ42/mg of total proteins). Note that Aβ38 peptide remained below the detection range for both genotypes and, as expected, control flies did not accumulate any human Aβ peptide. These data indicate that a ∼1.6-fold increase in dTau expression level did not modulate AβPP amyloidogenic processing nor Aβ peptide accumulation or stability in vivo.
Moderate increase of dTau expression does not modulate the Aβ peptides steady-state levels
Quantification of the Aβ38, Aβ40, and Aβ42 peptides by MSD analyses on protein extracts from control (elavC155-Gal4> +), AβPP/BACE1 (elavC155-Gal4 > UAS-AβPP; UAS-BACE1), or dTau/AβPP/BACE1 (elavC155-Gal4 > UAS-dTau, UAS-AβPP; UAS-BACE1) flies. Aβ quantities were normalized using the total protein content in each sample.
Moderate increase in dTau expression level in AβPP/BACE1 flies leads to the accumulation of insoluble dTau oligomers in vivo
As tau proteins are recovered as insoluble aggregates in AD patients’ brains, we next investigated whether expression of AβPP and its derivatives could modulate the overall levels of accumulation, solubility, and/or aggregation of dTau. Western blotting analysis, performed on total proteins extracts using the anti-Drosophila tau 13114 antibody [27], showed in control flies that endogenous dTau proteins migrated as a single band with an apparent molecular mass of ∼60 kDa (Fig. 3). Neuronal co-expression of AβPP and BACE1 did not significantly modify amounts of endogenous dTau (p > 0.05). In agreement with qRT-PCR analysis (Fig. 1), expression of the exogenous dTau protein led to a moderate but significant increase in total dTau accumulation (218.4%; p < 0.0001). Compared to dTau, co-expression of AβPP/BACE1 and dTau did not significantly modulate total amounts of dTau (p > 0.05), although we detected a decreasing trend (174.5%).
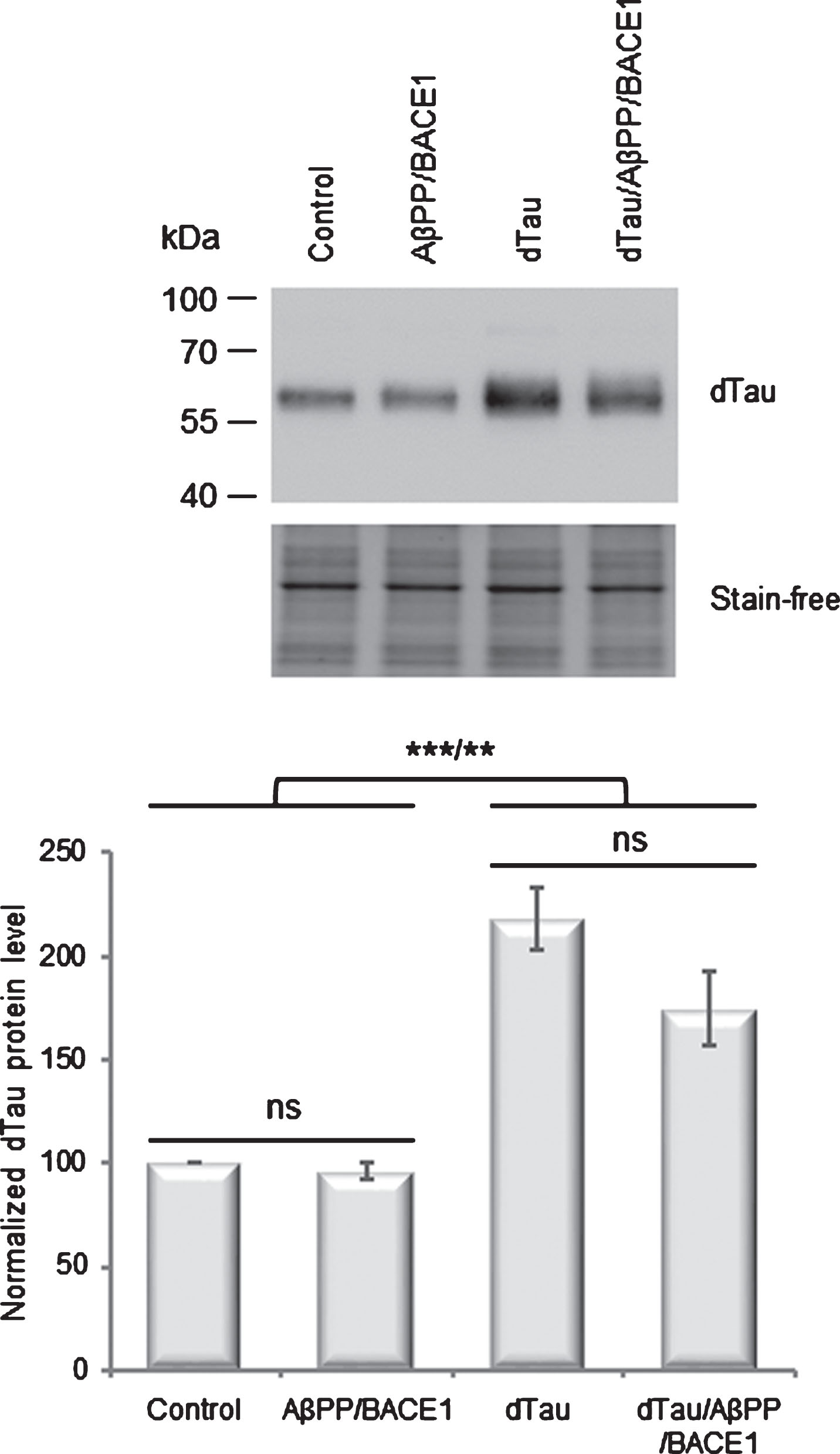
AβPP/BACE1 expression does not modulate dTau protein steady-state level. Western blot analyses of total protein extracts from control flies (elavC155-Gal4> +), flies overexpressing dTau (elavC155-Gal4 > UAS-dTau), AβPP/BACE1 (elavC155-Gal4 > UAS-AβPP; UAS-BACE1), or dTau/AβPP/BACE1 (elavC155-Gal4 > UAS-dTau, UAS-AβPP; UAS-BACE1), using dTau 13114 antibody. Total protein was used as loading control using stain-free technology and the normalized expression of the dTau protein is reported in the graphs. Bar graph: mean±standard error of the mean (SEM) of relative dTau protein levels. Controls were arbitrarily set at 100. One-way analysis of variance (ANOVA) and Tukey’s post hoc analyses were performed to determine the differences in dTau accumulation levels between genotypes (F (3,12) = 25.47; p < 0.0001; n = 4). AβPP/BACE1 and control flies accumulated similar amounts of dTau proteins (96.1%; p > 0.05). In dTau flies, we observed a moderate and significant increase in dTau accumulation as compared to controls (218.4%; p < 0.0001). This increase was not significantly modulated in dTau/AβPP/BACE1 flies (174.5%; p > 0.05). Ns, not significant p > 0.05; **p < 0.01; ***p < 0.001.
To analyze the solubility and the aggregation of dTau proteins on AβPP/BACE1 co-expression, we sequentially extracted soluble and insoluble proteins and analyzed both fractions by western blot (Fig. 4). In the soluble fraction, dTau proteins were recovered as monomers (arrow head) in all genotypes. In the insoluble fraction, only a faint signal corresponding to monomeric species (arrow head) was detected upon longer exposure, indicating that dTau monomers are predominantly recovered as soluble species. Neither co-expression of AβPP/BACE1 nor expression of exogenous dTau modified the profile of these monomeric species. However, when AβPP, BACE1 and dTau were expressed together, we specifically detected an additional band in the insoluble fraction (arrow). Given its apparent molecular weight (∼130 kDa), this band may correspond to dTau dimers. Importantly, similar data were confirmed using another anti-dTau antibody [28] (Supplementary Figure 2).
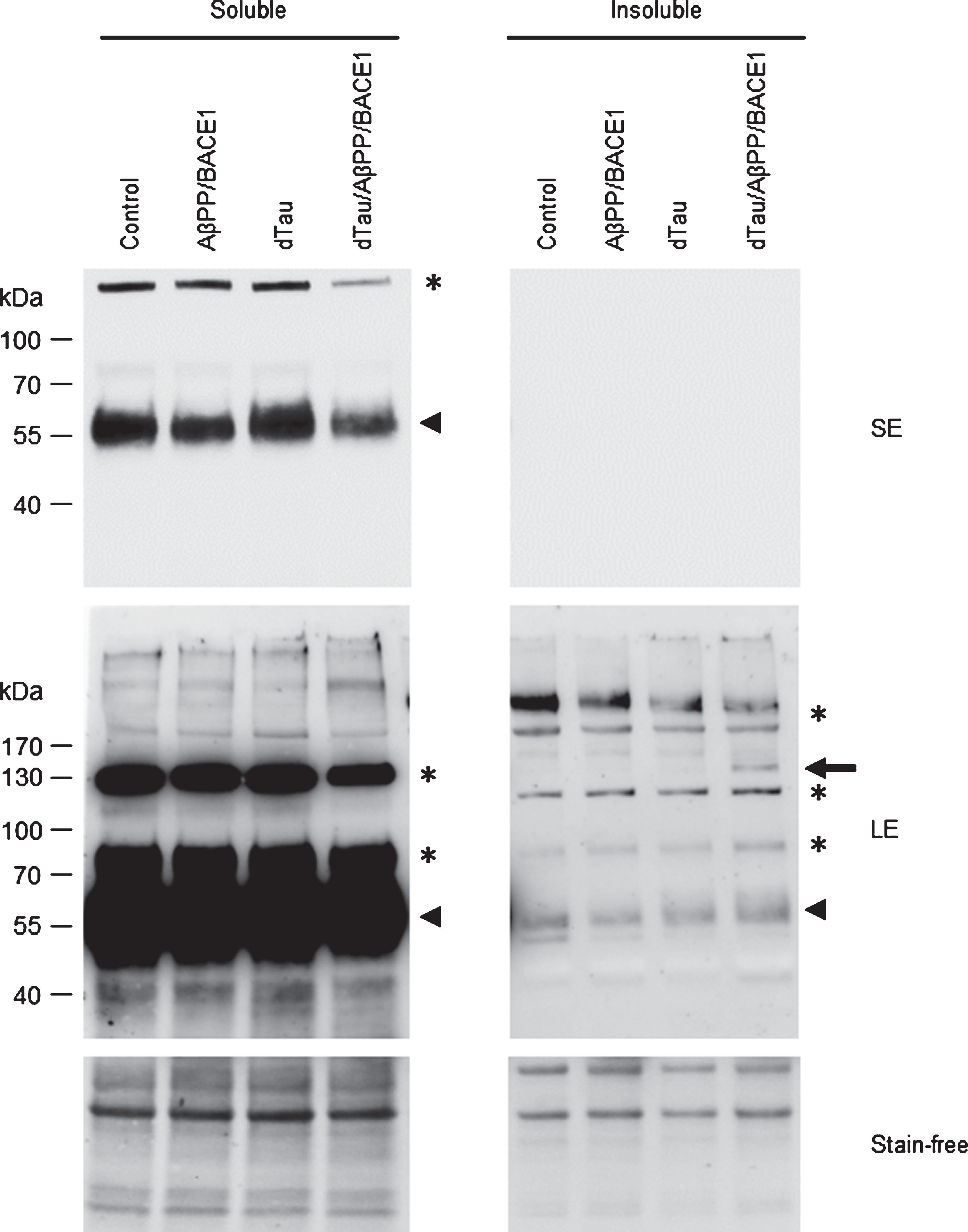
Moderate overexpression of dTau in AβPP/BACE1 flies leads to the accumulation of insoluble dTau oligomers. Western blot analysis of sequentially extracted proteins from control flies (elavC155-Gal4> +), flies overexpressing dTau (elavC155-Gal4 > UAS-dTau), AβPP/BACE1 (elavC155-Gal4 > UAS-AβPP; UAS-BACE1), or dTau/AβPP/BACE1 (elavC155-Gal4 > UAS-dTau, UAS-AβPP; UAS-BACE1), using the dTau 13114 antibody. dTau monomers (Arrowhead) were predominantly recovered as soluble species. Similar migration profiles were detected in all genotypes, except for the presence of an additional band corresponding to oligomeric species at ∼130 kDa (Arrow) in the insoluble fraction from dTau/AβPP/BACE1 flies. SE, short exposure; LE, long exposure. *unspecific signal.
Thus, a ∼1.6-fold increase in dTau expression level in AβPP/BACE1 flies resulted in the accumulation of insoluble dTau oligomers in vivo.
DISCUSSION
In the present report, we investigated whether the difference in MAPT expression level observed between H1 and H2 haplotypes could influence Aβ-induced toxicity in vivo, and therefore could be related to H1-associated LOAD risk. To mimic the difference in MAPT expression level observed between H1 and H2 haplotypes, we selected a Drosophila transgenic line that expresses dTau protein ∼1.6-fold over endogenous level. We previously showed that, in contrast to human tau proteins that are hyperphosphorylated and bind very poorly to microtubules when overexpressed in flies, the exogenous dTau protein binds efficiently to microtubules [24], and therefore likely retains its biological activities in vivo. Importantly, this ∼1.6-fold increase in dTau expression level in neurons did not result in any obvious phenotype in flies upon eclosion. We also used a Drosophila model co-expressing the human AβPP and BACE1 genes [25]. The expression of AβPP and BACE1 recapitulates AβPP amyloidogenic processing (notably the production of Aβ40 and Aβ42 peptides) in flies and leads to several neuronal phenotypes, including wing expansion defects, memory and locomotor deficits. and synaptic defects.
We found that a ∼1.6-fold increase in dTau expression level exacerbated neuronal phenotypes with wing expansion defects and locomotor deficits in AβPP/BACE1 flies. The exacerbation of these neuronal phenotypes correlated with the accumulation of insoluble dTau oligomers, suggesting the potential involvement of these small tau aggregates in the toxicity processes. Consistent with these data, tau oligomers have emerged as the pathogenic species in tauopathies and possible mediators of Aβ toxicity in AD [33, 34]. In patients’ brains, tau oligomers are mainly constituted of dimers and trimers and they can, later in the disease process, form filaments and tangles [35]. Numerous studies have already demonstrated that tau oligomers are toxic both in vitro and in vivo. Low-n oligomeric tau is neurotoxic by injuring spine morphology and density when applied to primary neurons at very low concentrations [36, 37]. In mice, tau oligomers have been implicated in synaptic loss [38–40], correlate with memory deficits through synaptic and mitochondrial impairments [38, 41] and potentiate neuronal damage, leading to neurodegeneration following traumatic brain injury [42, 43].
These dTau insoluble oligomers appear specifically in dTau/AβPP/BACE1 triple transgenic flies. No dTau oligomer was detected with dTau overexpression alone, indicating that expression of AβPP and its derivatives is essential to induce dTau oligomerization. These data are fully consistent with numerous studies showing that AβPP and Aβ expression enhances tau aggregation in in vivo and in vitro models [44]. Several mechanisms have been proposed to explain how Aβ acts to enhance tau aggregation. First, studies realized in cellular and animal models, including Drosophila models of AD have suggested the involvement of Aβ in the pathological phosphorylation of tau through the activation of several tau kinases (GSK3β, PAR-1/MARK, CAMKK2-AMPK) [45–51]. The pathological phosphorylation of tau would then induce its self-assembly presumably by altering the charge and structure of the protein. The Aβ peptides could also act as direct seeds for tau aggregation. Indeed, in vitro analysis has demonstrated that low amounts of Aβ oligomers are able to induce tau oligomerization [52].
Importantly, no dTau oligomers were apparent when AβPP and BACE1 were expressed on a wild-type genetic background (no overexpression of dTau), suggesting that the expression level of dTau is crucial and that a slight increase in dTau expression level allows the production of oligomeric species. Different mechanisms can be proposed to explain these results. We previously showed that exogenous dTau protein binds efficiently to microtubules [24]. Tau interacts with microtubules via a kiss-and-hop mechanism in neuronal processes [53]. Tau dwells on a single microtubule for a very short time (∼40 ms) before it hops to the next, suggesting a very dynamic equilibrium between microtubule-bound and unbound tau fractions. Thus, we hypothesize that the moderate increase in dTau steady-state level could lead to a rise in the microtubule-unbound dTau fraction, and therefore in the number of dTau molecules that could be targeted directly or indirectly by Aβ species. Second, the binding of tau protein to the microtubules is known to be regulated by coordinated actions of kinases and phosphatases [54, 55]. Thus, Aβ-mediated tau phosphorylation could also interfere with dTau-microtubule-binding, and therefore increase the amount of modified tau proteins that can form oligomeric species. Lastly, numerous studies have shown that the aggregation reaction depends on monomer concentration. Given that it is commonly accepted that the tau oligomers detected in patients’ brains are composed of both abnormally modified tau protein and normal forms of the protein [56, 57], the slight increase in the amount of dTau protein in our model could also contribute to the formation of oligomers via the recruitment of normal monomeric species. Importantly, these mechanisms are not mutually exclusive and may work together to promote the formation of dTau oligomers.
In this study, we focused on the effect of the difference in MAPT expression level observed between H1 and H2 haplotypes and its contribution to LOAD risk. Of course, disease risk associated with MAPT haplotypes may also involve other pathogenic mechanisms, such as alteration in MAPT gene splicing and imbalance of tau isoforms. These mechanisms are not mutually exclusive, and may both be involved in disease processes.
To conclude, in this study, we have shown that modest overexpression of tau protein in Drosophila exacerbates neuronal phenotypes in APP/BACE1 flies. The exacerbation of the neuronal defects correlates with the accumulation of insoluble dTau oligomers. Our data suggest that a moderate difference in tau expression level similar to that observed between H1 and H2 haplotypes could influence Aβ-induced toxicity in vivo, and thus could contribute to LOAD risk.
Footnotes
ACKNOWLEDGMENTS
We thank Dr. Daniel Marenda and the Bloomington Drosophila stock center for providing fly stocks and Dr. Linda Partridge for providing the Drosophila Tau SK6427 antibody. This work was supported by a grant from the Fondation pour la Recherche Médicale (FRM, grant # DEQ20170336711) and the European Union and the Région Normandie. Europe gets involved in Normandie through the European Regional Development Fund (ERDF). The authors are grateful to Nikki Sabourin-Gibbs, Rouen University Hospital, for her help in editing the manuscript.