
Abstract
Alzheimer’s disease (AD) is one of the most devastating and costly diseases, and prevalence of AD increases with age. Furthermore, females are twice as likely to suffer from AD compared to males. The cessation of reproductive steroid hormone production during menopause is hypothesized to cause this difference. Two rodent AD models, APP21 and APP+PS1, and wild type (WT) rats underwent an ovariectomy or sham surgery. Changes in learning and memory, brain histology, amyloid-β (Aβ) deposition, levels of mRNAs involved in Aβ production and clearance, and synaptic and cognitive function were determined. Barnes maze results showed that regardless of ovariectomy status, APP+PS1 rats learned slower and had poor memory retention. Ovariectomy caused learning impairment only in the APP21 rats. High levels of Aβ42 and very low levels of Aβ40 were observed in the brain cortices of APP+PS1 rats indicating limited endogenous PS1. The APP+PS1 rats had 43-fold greater formic acid soluble Aβ42 than Aβ40 at 17 months. Furthermore, levels of formic acid soluble Aβ42 increased 57-fold in ovariectomized APP+PS1 rats between 12 and 17 months of age. The mRNA encoding Grin1 significantly decreased due to ovariectomy whereas levels of Bace1, Chat, and Prkcb all decreased with age. The expression levels of mRNAs involved in Aβ degradation and AβPP cleavage (Neprilysin, Ide, Adam9, and Psenen) were found to be highly correlated with each other as well as hippocampal Aβ deposition. Taken together, these results indicate that both ovariectomy and genotype influence AD markers in a complex manner.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) causes roughly 60–80% of all cases of dementia. In addition to increasing age, female gender is also a predisposing risk factor for the development of AD. In fact, the estimated risk of developing AD is double in women compared to men [1]. Estrogens have numerous neuroprotective effects relevant to the prevention of AD and the loss of estrogens in females during aging and the menopausal period may accelerate AD-like pathology [2]. Ovaries are the main organs that produce female steroid sex hormones including both estrogens and progesterone. Ovarian function and hormone production decline with age and menopause marks the onset of significant follicular depletion and subsequent reduction in reproductive hormone production in women and experimental animal models [3, 4]. There is also recent evidence that suggests that higher levels of luteinizing hormone may be responsible for cognitive impairment in females with reduced estrogen levels due to menopause or an ovariectomy procedure [5–7].
Bilateral ovariectomies have been widely used to elucidate the therapeutic and/or adverse effects of ovarian hormones on women’s health and disease including cognitive well-being [8–11], metabolic disorders such as osteoporosis [10, 12], and cardiovascular diseases [13]. Ovariectomies preceding the onset of menopause are associated with an increased risk of cognitive impairment or dementia in an age dependent manner [14]. The decline of the ovarian hormone estrogen has been proposed as one of the major causes of cognitive impairment in women. The higher incidence of AD in females [15, 16] has been attributed to gradual estrogen withdrawal. Restoration of cognitive function after estrogen therapy in menopausal women [15, 18] and ovariectomy-induced menopause in animal models [19, 20] provide further evidence of estrogen’s role in cognitive health. Furthermore, ovariectomy and estrogen have been shown to influence levels of mRNA-encoding proteins involved in AD in the brains of experimental animals [21] and cultured cells [18].
Animal models are an invaluable resource that enable further understanding of disease cause, progression, and treatment. APP21 rats that overexpress human AβPP with Indiana and Swedish mutations [22] and the double transgenic APP+PS1 rats that overexpress the same AβPP transgene as well as human presenilin 1 with the L166P mutation [23] are two such AD models. Both the APP21 and APP+PS1 rat strains have a Fischer 344 background and are homozygous for human AβPP transgene; the APP+PS1 rats are also hemizygous for the human PS1 transgene. The APP21+PS1 rats have been shown to develop Aβ plaques and cerebral amyloid angiopathy (CAA), whereas the APP21 rats have not been shown to develop these pathologies [24]. Although APP21 rats do not develop pathologies such as Aβ plaques or CAA most often associated with AD in humans, they do develop subtle behavior and pathologic changes associated with the initial stages of AD [25]. The APP+PS1 rats display greater cognitive deficiencies compared to APP21 rats [23, 24].
In this study, we investigated the effect of ovariectomy-induced estrogen and progesterone withdrawal as a model for menopausal cognitive function as well as molecular and histological changes in brains of APP21, APP+PS1, and wild type (WT) Fischer 344 rats. Our hypothesis was that ovariectomy would lead to memory deficits in all ovariectomized rats and an increase in Aβ accumulation in the brains of APP+PS1 rats. Five months after bilateral ovariectomy when the rats were 7 months old, cognitive function was assessed and molecular and histological changes in the brain were assessed at 12 and 17 months of age given Aβ accumulation in the brains of APP+PS1 rats begin around 1 year of age. Reproductive cycle in female rats begin approximately at 6 weeks of age and thus, 2-month-old rats are comparable to 16-20-year-old humans. At 7 months of age, intact rats have a large number of antral follicles that produce ovarian hormones; however, rats 12 months and older have a drastically reduced number of antral follicles [4] which leads to reduction in sex hormone production. Rats that are 12 and 17 months of age are comparable to the perimenopause and menopause stages in humans. Thus, postmortem examinations conducted when rats are 12 and 17 months old should reflect the long-term effects of ovariectomy and aging, but not differences in levels of sex hormones. On the other hand, behavioral assessments should reflect the long-term effects of ovariectomy as well as differences in sex hormone levels.
MATERIALS AND METHODS
Animals
A total of 74 rats were used in this study: 21 APP21, 25 APP+PS1, and 28 WT. The APP21 and APP+PS1 rats were littermates and were born in the facility in which the experiments were performed. The APP21 rats were homozygous for the human amyloid-β protein precursor transgene with Swedish and Indiana mutations. The APP+PS1 rats had the APP21 background and thus they were homozygous for the AβPP transgene and hemizygous for the human PS1 transgene with L166P mutation. The crossing of APP21 and APP+PS1 rats results in approximately 50% human AβPP homozygous (APP21) rats and 50% human AβPP homozygous and human PS1 hemizygous (APP+PS1) rats. The wild type Fischer 344 (WT) rats were purchased from Envigo (Indianapolis, IN) at 4 weeks of age. At 8 weeks of age, rats were ovariectomized or underwent a sham operation that left their ovaries intact. Thus, there were 6 experimental groups: Ovariectomized APP21 (APPOvEx; n = 9), APP+PS1 (PS1OvEx; n = 13), WT (WTOvEx; n = 14), sham-operated APP21 (APPSham; n = 12), APP+PS1 (PS1Sham; n = 12), and WT (WTSham; n = 14). A timeline of the experiments is outlined in Table 1. Behavioral tests were conducted when the rats were 7 months old. Molecular and histological changes in the brain were determined at 12 and 17 months of age to allow further aging in order to increase amyloid deposition in APP+PS1 rats. Half (n = 37) of the rats were euthanized at 12 months of age and the remaining half were euthanized at 17 months of age.
Timeline of the experiments and number (n) of animals used.
Rats were housed in conventional cages at 20–25°C with free access to food and water. All of the animal studies performed were approved by the University of Missouri’s Animal Care and Use Committee and were in accordance with the guidelines of the Institute for Laboratory Animal Research Guide for the Care and Use of Laboratory Animals.
Bilateral ovariectomy
For bilateral ovariectomy, the rats were anesthetized using an IP injection of a combination of Ketamine (80 mg/kg) and Xylazine (9 mg/kg), and subcutaneous injection of buprenorphine (0.05 mg/kg) was used as analgesic to prevent post-operative stress. Standard surgical procedures were used to enable access to the ovaries. The skin of the dorsum was sagittally incised (∼10–15 mm in length) at the midline at the level of the para-lumbar fossa. The skin was separated from the underlying tissue by blunt dissection, and the muscle layers and the peritoneal wall were opened via incision and spread to create a window about 8–10 mm in length that permitted retraction of the ovarian fat pad. Each ovary was gently grasped with tissue forceps and retracted/extracted until the ovary and the oviduct was exposed and visualized under a stereomicroscope. The whole ovary including the ovarian bursa was removed to ensure total eradication of the ovary and then cauterized to prevent bleeding. The uterine horn was then carefully replaced into the peritoneal cavity. The muscle layers were sutured with 4-0 Vicryl (Ethicon). After repeating the same procedure on the contralateral side, the skin was closed using wound clips. The ovaries of sham operated rats were exposed, but returned to the abdominal cavity without any further treatment.
The cyclicity of the rats was determined using vaginal cytology where the presence of mostly leucocytes indicated diestrus and the absence of reproductive cycling. The presence of epithelial cells and cornified epithelial cells indicated proestrus and estrus and therefore confirmed the reproductive cyclicity of the rats. Estrus cycles were not detected in the ovariectomized rats whereas sham-operated rats were confirmed to be cycling.
Behavior analysis
Barnes maze
All rats were tested for spatial memory and learning at 7 months of age using the Barnes maze. The maze consisted of a gray circular platform 122 cm in diameter, surrounded by a wall that was 30.5 cm in height. The maze was elevated 83.8 cm above the floor by a stand. Twenty holes measuring 10.2 cm in diameter were evenly spaced around the perimeter. A rectangular gray goal box (28 cm in length×12.7 cm wide×7.6 cm high at the area closest to the maze tapering to 16.5 cm high) could be placed beneath any hole. The goal box included an entry ramp that provided easy entry access for the rat. Black curtains were hung around the maze and above the maze walls to surround the apparatus and ensure that rats could only use the visual cues provided in the maze, rather than the distal cues within the testing room, to navigate around the maze. Four visual cues consisting of various shapes (triangle, square, circle, cross) were placed at evenly spaced intervals on the inside of the maze walls. Two 86-W, 120-V floodlights producing 1690 lumens hung above the platform served to brightly light the maze in order to create a potentially mildly aversive environment to help motivate the rats to escape from the brightly lit, open surface in favor of the dark environment of the goal box.
Each rat was assigned a goal box location and the goal box location was alternated across rats to eliminate odor cues for consecutively tested rats. The goal box location remained constant for any individual rat across test trials and the location of the goal box was also counterbalanced across experimental groups. Before the start of behavioral testing, each rat was pre-exposed to the goal box for 90 s. Behavioral testing consisted of 8 acquisition trials (2 trials/day) over a period of 4 days.
On each day that behavioral testing occurred, rats were transported to the testing room from the colony room in their home cages and acclimated in the testing room for a minimum of 30 min prior to the first trial. All rats were run in the Barnes Maze in the mornings during the light portion of the light/dark cycle. An acquisition trial began by placing the rat under a gray, opaque start box (23 cm length×23 cm wide) positioned in the center of the platform. After 30 s, the box was lifted and the rat had a maximum of 5 min (300 s) to locate and enter the goal box. Latency (time it took for the rat to find and enter the goal box) and total errors (nose-pokes into non-escape holes) were recorded. If the rat did not enter the goal box within 5 min, it was gently guided there by the experimenter’s hand. Once the rats had entered the goal box, the entrance of the goal box was covered to prevent escape back onto the maze platform. After 30 s, the rat was removed from the goal box and returned to its home cage. Rats were allowed to rest in their home cage in the testing room for 30 min before starting their subsequent daily trial. The platform and goal box were cleaned after every trial with a 20% ethanol solution. After the fourth day, testing abated for 14 days, after which retention was evaluated with 2 additional trials (1 day), conducted exactly like previous acquisition trials. The following day, rats were given reversal training, in which the rats were given trials exactly like those of original acquisition training, except that the goal box was moved to a new location in the opposite quadrant. Reversal training continued for a total of 4 days (2 trials/day).
Aβ40 and Aβ42 ELISA
TBS soluble fraction
To determine free Aβ, each parietal brain cortex was ground using liquid nitrogen mortar and pestle and 40–50 mg of tissue was placed in a tube and kept on dry ice until homogenization. Samples were homogenized on ice using a glass homogenizer with 500μl TBS buffer containing 140 Mm NaCl, 3 mM KCl, 25 mM Tris-HCl, pH7.4, 4 mM EDTA, 2 mM 1,10-Phenanthroline, and protease inhibitor cocktail (Sigma catalog number P8340). Homogenized samples were transferred into 1.5 ml tubes and centrifuged at 20,900× g for 1 h at 4°C. Supernatant (TBS soluble fraction) was stored in a –80°C freezer.
Formic acid soluble fraction
To determine the remaining Aβ content, pellets obtained from the TBS soluble fraction were dissolved in 500μl 70% formic acid containing 4 mM EDTA, 2 mM 1,10-Phenanthroline, and protease inhibitor cocktail by pipetting up and down followed by sonication for 5 s on ice. Samples were transferred into 1.5 ml tubes and centrifuged at 20,900× g for 1 h at 4°C. Formic acid was neutralized by adding 20 volumes of 1 M Tris base. Neutralized samples were stored in a –80°C freezer.
Brain and serum Aβ40 and Aβ42 ELISA
Tris buffered saline and FA soluble brain cortex and serum Aβ40 and Aβ42 levels were analyzed using Invitrogen (Carlsbad, CA) human Aβ40 (catalog number KHB3481) and Aβ42 (catalog number KHB3441) kits. The TBS and FA soluble fractions of brain cortex and serum samples were further diluted with the addition of a protease inhibitor cocktail. The TBS and FA soluble fractions were transferred to Aβ40 and Aβ42 ELISA plates and human Aβ40 or Aβ42 detection antibody was added to each well. Plates were then incubated at room temperature for 3 h. Following incubation, wells were washed and incubated with antirabbit IgG HRP followed by incubation with chromogen solution for 30 min each. The reactions were stopped and absorbance was determined at 450 nm and the Aβ concentration in the samples was calculated from standard curves.
Real-time RT PCR
The parietal brain cortices of ovariectomized and sham-operated APP21, APP+PS1, and WT rats at 12 and 17 months of age were used for real-time RT-PCR. Total RNA was isolated using Tri reagent (Molecular Research Center Inc., Cincinnati, OH) and quality was determined using agarose gel electrophoresis. Reverse transcription reactions were carried out using 1μg total RNA, 0.5μg of each oligo dT and random hexamer primers, and GoScript reverse transcriptase (Promega, Madison WI). The cDNAs that were amplified included ADAM metallopeptidase domain 9 (Adam9), beta-secretase 1 (Bace1), choline O-acetyltransferase (Chat), growth associated protein 43 (Gap43), glutamate ionotropic receptor NMDA type subunit 1 (Grin1), insulin degrading enzyme (Ide), membrane metallo endopeptidase (Mme; neprilysin), protein phosphatase 2 catalytic subunit alpha (Ppp2ca), protein kinase cAMP-activated catalytic subunit beta (Prkacb), protein kinase C beta (Prkcb), presenilin enhancer gamma secretase subunit (Psenen), ubiquitin C-terminal hydrolase L1 (Uchl1) and hypoxanthine phosphoribosyltransferase 1 (Hprt1). Real-time PCR reactions were carried out using 100 ng cDNA, gene specific primers (Supplementary Table 1) and iTaq Universal Sybr Green Supermix (Bio-Rad, Hercules, CA) using the Bio-Rad CFX384 Touch Real-Time PCR Detection System. Hprt1 was used as the house keeping gene to correct for pipetting differences between samples. The threshold cycle (Ct) for the Hprt1 was subtracted from the Ct for the test mRNA to obtain the change (Δ) in Ct (ΔCt). The relative amount of each mRNA was calculated by using the equation 2–ΔCt.
Immunohistochemistry
Brains from each experimental group at 12 and 17 months were used for immunohistochemical staining. Brains were fixed in 10% neutral buffered formalin for 48 h at room temperature and embedded in paraffin. Eight μm thick tissue sections were placed on poly-L-lysine-coated slides and incubated at 60°C for 1 h. Deparaffinization and rehydration of tissue sections were done by washing in xylene (3 changes; 4 min each) followed by washing in 100, 95, and 70% ethanol (4 min each) and rinsing with distilled water 3 times. Antigen retrieval was done using target retrieval solution (Dako S1699). Slides were incubated in dual endogenous enzyme blocking reagent (Dako S2003, ready-to-use) for 10 min at room temperature for quenching. Slides were washed with Dako wash buffer (Dako S3006; 3 changes, 4 min each) before incubation with primary antibody for 24 h at 4°C. After washing, they were incubated in EnVision+ dual link system-HRP secondary antibodies (Dako K4064) for 30 min at room temperature and washed 3 times with Dako wash buffer (4 min each). They were incubated with substrate-chromogen for 10 min (DAB; Dako K3468) and the reaction was stopped using distilled water.
The primary antibodies that were used were the Aβ antibody (6E10; Catalog # 803004; diluted 1:200; Biolegend Inc.), the lba1 antibody (Catalog #MABN92; diluted 1:2000; EMD Millpore, Temecula, CA, USA), and the GFAP antibody (Catalog #MU020-UC; diluted 1:8000; BioGenex, Fremont, CA).
Statistical analysis
The general linear models (GLM) procedure in SAS/STAT® (SAS 9.4; SAS Institute Inc., Cary, NC, USA) was used to analyze the ELISA, real time RT-PCR, and immunohistochemistry results and effects of age, OvEx, and transgene were determined. All behavioral statistical analyses were conducted using SPSS (SPSS Statistics 23, IBM Corp., Armonk, NY, USA) and analysis of variance (ANOVA). For the Barnes maze, both daily latency and error performance were averaged across the two trials each day and the days were used in the repeated-measures analysis. Day was used as the within-subjects factor and genotype and ovariectomy were used as between-group factors. Comparisons of performance were analyzed separately for each training phase (acquisition, retention, reversal) and Tukey HSDi post-hoc tests were used for pairwise comparisons. Differences were considered significant at p < 0.05 for all analyses.
RESULTS
Ovariectomies were performed at approximately 2 months of age and all rats were weighed before the surgery. The weights of the rats were similar between groups (p > 0.05) before the ovariectomy procedure. However, after surgery, the body weights of all ovariectomized groups were greater than the sham-operated groups starting one month after the ovariectomy up through 13 months of age (p < 0.01; Supplementary Figure 1).
Barnes maze
Acquisition
Overall, both error and latency performance improved for all groups with over acquisition training (ps < 0.001; data not shown), and pairwise comparisons revealed that WT animals made fewer errors than APP21 rats (p < 0.04) and APP+PS1 rats (p > 0.07), as shown in Fig. 1A. Additionally, sham animals had significantly shorter latencies than OvEx animals (p < 0.04; Fig. 1B), but not fewer errors than OvEx animals (p > 0.53; Fig. 1A).
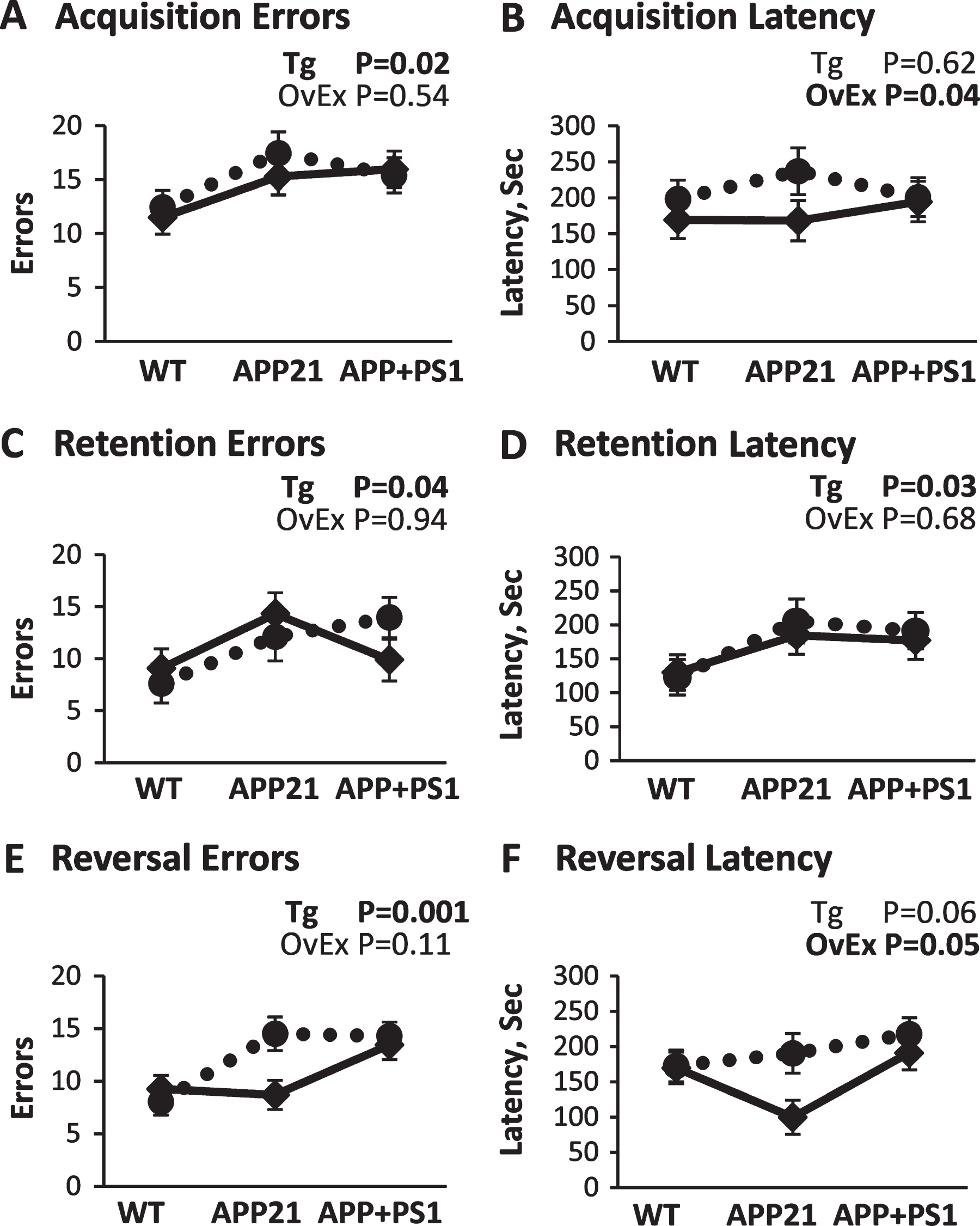
Barnes Maze results. A) Error performance during the acquisition training phase collapsed over trials and training days. B) Latency performance during the acquisition training phase collapsed over trials and training days. C) Error performance during the retention probe phase collapsed over trials. D) Latency performance during the retention probe phase collapsed over trials. E) Error performance during the reversal training phase collapsed over trials and training days. F) Latency performance during the reversal training phase collapsed over trials and training days. Solid lines represent sham-operated and dotted lines represent ovariectomized rats. p-values for the main effects of genotype (Tg) and ovariectomy (OvEx) are displayed in each graph.
Retention
Performance during the retention test showed that APP21 rats made statistically more errors (p < 0.04) and had longer latencies (p = 0.05) than WT rats, as shown in Fig. 1C and 1D. APP+PS1 rats did not differ from the other two groups on these two measures (ps > 0.14; Fig. 1C, D). There was no significant main effect of ovariectomy on retention performance (ps > 0.05; Fig. 1C, D).
Reversal
Overall, both error and latency performance significantly improved for all groups with reversal training (ps < 0.01; data not shown). As shown in Fig. 1E and 1F, pairwise comparisons found that overall, APP+PS1 animals made significantly more errors than WT animals (p < 0.002), but not APP21 animals (p > 0.14), and had significantly longer latencies than APP21 rats (p < 0.03), but not WT rats (p > 0.31). In addition, OvEx animals demonstrated significantly longer latencies than sham animals, as shown in Fig. 1F. Ovariectomy had a more detrimental effect on the APP21 animals than the APP+PS1 animals, as most clearly seen in Fig. 1E and 1F. There was a significant two-way interaction between day and ovariectomy for errors (p < 0.05; data not shown), such that OvEx rats started off making a lot of errors but improved over days while sham animals made fewer errors overall but did not improve as much over time. There was also a significant genotype and ovariectomy interaction for errors (p < 0.05; Fig. 1E). Ovariectomy had little to no effect on errors in the WT and APP+PS1 rats, such that both WT groups performed well, whereas both APP+PS1 groups performed poorly. APP21 rats, on the other hand, were more sensitive to the ovariectomy manipulation; sham animals made few errors and performed similarly to the WT, while the OvEx animals made more errors and performed similarly to the APP+PS1 animals. A similar observation was made in latency performance, although the genotype and ovariectomy interaction was not statistically significant (p > 0.19; Fig. 1F). Additionally, there was a three-way interaction (p < 0.001; data not shown) for latency, such that ovariectomy resulted in performance improvements over days for both of the AD groups, but not for the WT rats. For sham operated rats, the WT and APP21 animals showed improved latency scores over days, but the APP+PS1 animals did not. Somewhat surprisingly, APPSham rats had the fastest numerical latencies than any of the other groups (Fig. 1F).
Real time RT-PCR
Relative amounts of mRNAs encoding Adam9, Ide, Mme, and Psenen, Chat, Grin1, Bace1, Prkcb, Ppp2ca, Prkacb, and Uchl1 were determined by real time RT-PCR (Fig. 2). Levels of mRNAs encoding enzymes involved in AβPP cleavage and Aβ degradation (Adam9, Ide, Mme, and Psenen) followed a distinct pattern and PS1OvEx and APPOvEx rats had lower levels of these mRNAs compared to their intact counterparts at 12 months. At 17 months, expression levels of these mRNAs increased in PS1OvEX and APPOvEx rats, but decreased in all the other experimental groups. Furthermore, levels of these mRNAs were highly correlated (r = 0.63 to 0.83; Fig. 2A-2D and Table 2). Ovariectomized rats had lower levels of Grin1 both at 12 months and 17 months (p = 0.04). Levels of Chat and Bace1 mRNAs were significantly reduced at 17 months compared to 12 months (p = 0.007 and p = 0.04, respectively). Correlation analysis of mRNAs showed that Grin1 and Bace1 were highly correlated (r = 0.79; Fig. 2E, F and Table 2). The level of Prkcb mRNA significantly (p = 0.05) was increased at 17 months compared to 12 months. Expression of Gap43, Ppp2ca, Prkacb, and Uchl1 mRNAs were not influenced by age, ovariectomy, or transgene (ps > 0.1). Levels of another group of mRNAs including Ppp2ca, Prkacb, and Prkcb which are involved in signal transduction as well as Uchl1 were also correlated with each other (rs = 0.54 to 0.80; Table 2).

Brain cortex real-time RT-PCR analysis of ADAM metallopeptidase domain 9 (Adam9), membrane metallo endopeptidase (Mme), insulin degrading enzyme (Ide), presenilin enhancer gamma secretase subunit (Psenen), choline O-acetyltransferase (Chat), glutamate ionotropic receptor NMDA type subunit 1 (Grin1), beta-secretase 1 (Bace1), protein kinase C beta (Prkcb) in 12- and 17-month-old APP21 (open black), APP+PS1 (black), and WT (gray) rats. Solid lines represent sham-operated and dashed lines represent ovariectomized rats. 12M, 12 months of age; 17M, 17 months of age. p-values for the main effects of genotype (Tg) and ovariectomy (OvEx) are displayed in each graph.
Correlation of mRNA and Aβ deposition levels
The mRNA levels of the parietal brain cortex were detected by real time RT-PCR and APP21, APP+PS1, and WT rats were used for these correlations. Hippocampal dense and diffuse (Dif) Aβ deposition was detected by immunohistochemistry using 6G10 antibody and parietal brain cortex formic acid (FA) soluble Aβ40 and Aβ42 levels were detected by ELISA and only APP+PS1 rats were used for correlations of Aβ deposition. Upper section of the table shows correlation coefficients and highlighted values indicate r≥0.5. p-values are shown in the lower section in italic numbers.
Brain Cortex Aβ40 and Aβ42 ELISA
A subset of 17-month-old WT (n = 4) and APP21 (n = 4) brains were tested to determine TBS and FA soluble Aβ40 and Aβ42 levels in brain cortex and were not found to have detectable levels of TBS or FA soluble Aβ40 or Aβ42. Thus, Aβ40 and Aβ42 measurements included only APP+PS1 (n = 25) rats. The APP+PS1 rats did not have any measurable levels of TBS soluble Aβ40 in their brain cortices at 12 months. Levels of TBS soluble Aβ42 were significantly (p = 0.01) increased in 17-month-old APP+PS1 rats compared to 12-month-old rats. FA soluble Aβ42 was significantly greater in 17-month-old APP+PS1 rats compared to 12-month-old APP+PS1 rats (p = 0.0002). Five months of aging between 12 and 17 months led to a 13-fold increase in TBS soluble Aβ42 in APP+PS1 rats (Fig. 3A). Levels of FA soluble Aβ40 were comparable between 12-month and 17-month-old APP+PS1 rats (Fig. 3B). There was no significant age (p = 0.08) or ovariectomy effect (p = 0.6) nor an interaction between age and OvEx treatment (p = 0.9). Age significantly (p < 0.001) increased FA soluble Aβ42 levels; however, there was no ovariectomy effect (p = 0.4) and no interaction between age and ovariectomy (p = 0.4). Although not significant, the PS1OvEx rats had 69% more FA soluble Aβ42 compared to PS1Sham rats at 17 months of age (Fig. 3C). 17-month-old APP+PS1 rats had 37 times more FA soluble Aβ42 compared to Aβ40 (p < 0.0001; Fig. 3D). PS1OvEx rats had numerically greater FA soluble Aβ42/Aβ40 compared to PS1Sham at both 12 and 17 months, but this was not statistically significant (p = 0.17) and there was no interaction between age and ovariectomy (p = 0.2). Furthermore, FA soluble Aβ42 levels were significantly and negatively correlated (r = 0.54) with Chat mRNA levels (Table 2).
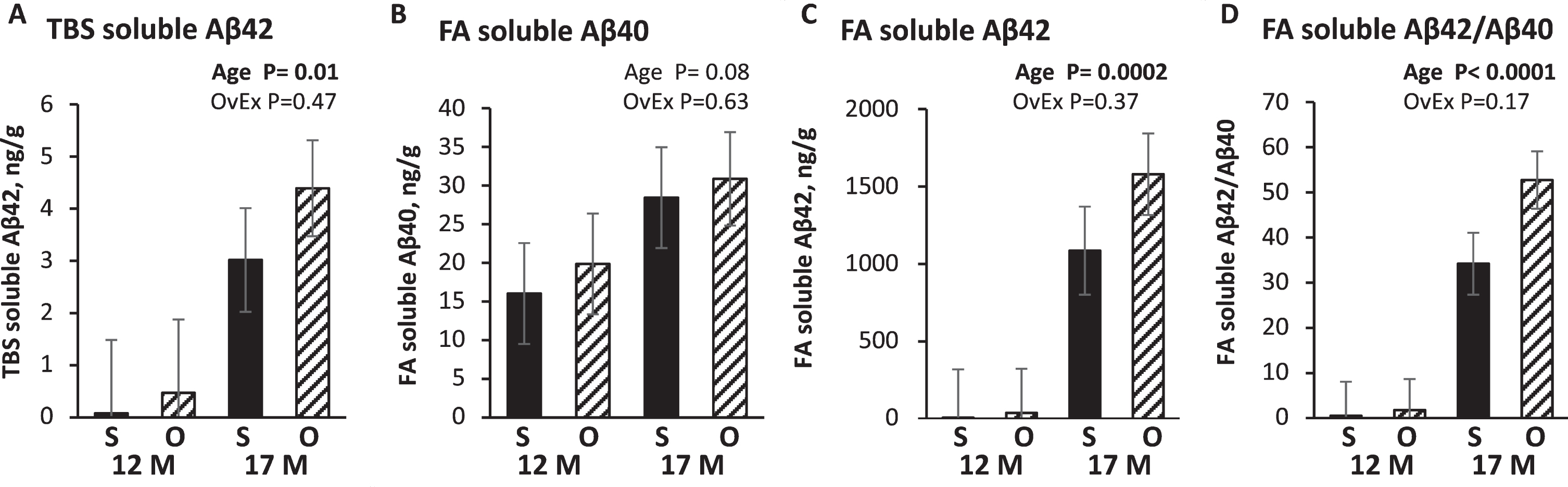
Brain cortex Tris buffered saline (TBS) and formic acid (FA) soluble Aβ40 and Aβ42 levels in APP+PS1 rats. S, Sham operated group; O, Ovariectomized group; 12M, 12 months of age; 17M, 17 months of age. Data is presented to show Aβ levels in each experimental group; p-values are displayed in each graph to show the main effects of ovariectomy (OvEx) and age.
A limited number of serum samples from 17-month-old APP21 and APP+PS1 rats were included in the ELISA analysis of Aβ40 and Aβ42 due to suboptimal sample processing. Serum Aβ40 levels of APP21 (254 pg/ml) and APP+PS1 (295 pg/ml) rats were not significantly different (p = 0.5). APP+PS1 rats were found to have significantly (ps < 0.02) greater serum Aβ42 compared to APP21 rats, 65 and 23 pg/ml, respectively. There was no ovariectomy effect (ps > 0.5) and no interaction between ovariectomy and transgene (p > 0.3) on serum Aβ40, Aβ42, or Aβ42/Aβ40 levels.
IHC analysis
Amyloid-β deposition
Aβ deposition using 6E10 antibody followed a distinct pattern in all APP+PS1 rats. Sporadic dense plaques were observed in the cortex and hippocampus at 12 months. Extensive diffuse plaque accumulation started in the subiculum region at 12 moths and extended to under the alveus hippocampi (ALV) at 17 months (Fig. 4A, B). Some of the diffuse Aβ staining revealed large reactive gemistocytic astrocytes that are associated with blood vessels. 17-month-old APP+PS1 rats had 16 times more diffuse plaques compared to 12-month-old APP+PS1 rats (p < 0.0001; Fig. 4C). Similarly, they had 15 times more dense plaques compared to 12-month-old APP+PS1 rats (p = 0.0002; Fig. 4D). There was no significant effect of ovariectomy and there was no interaction between ovariectomy treatment and age (ps > 0.7) for either dense or diffuse plaques. Furthermore, dense Aβ deposition is correlated (r = 0.49) to Adam9, Ide, and Psenen mRNA levels in APP+PS1 rats (Table 2).
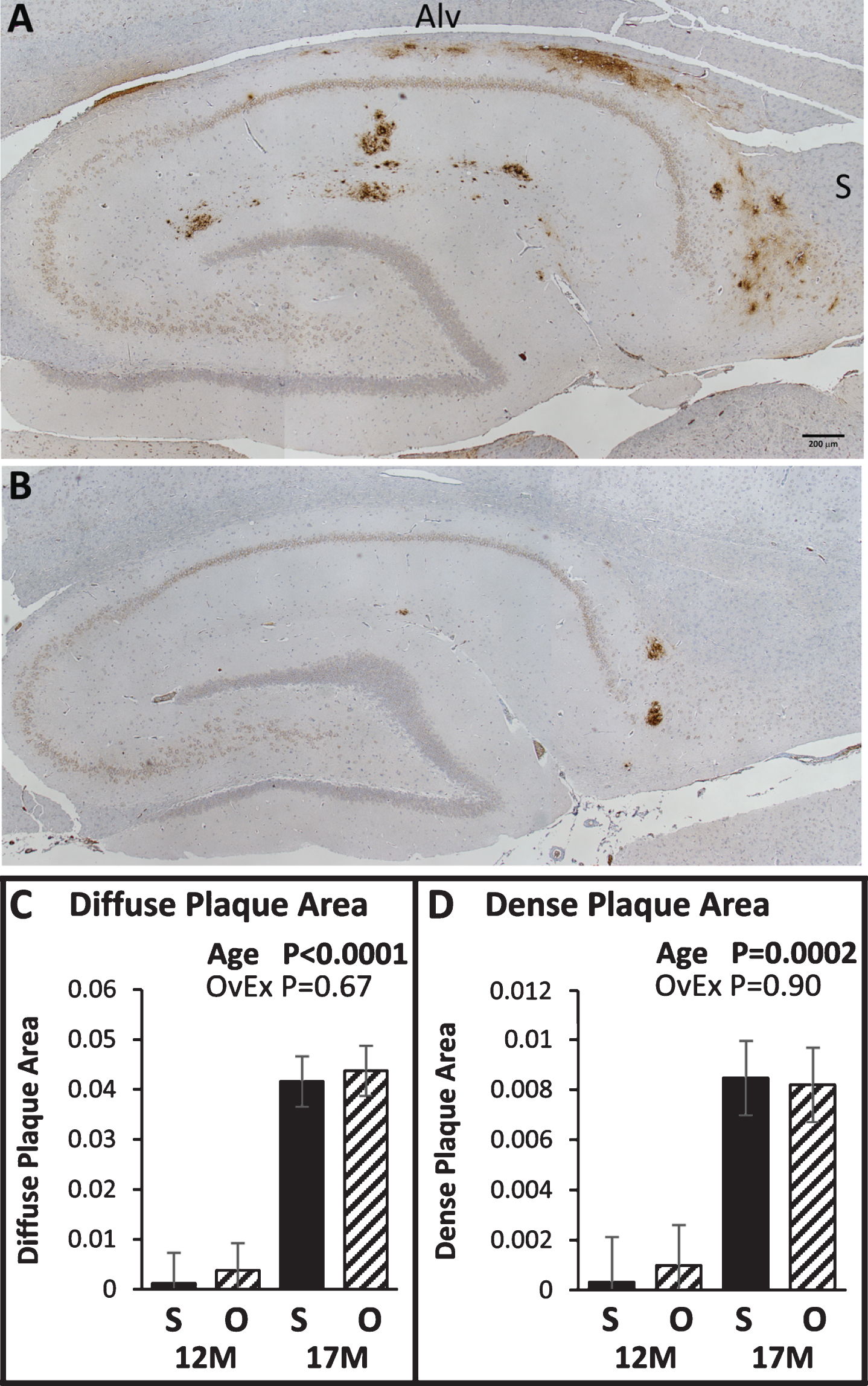
Hippocampus Aβ deposition in 17- (A) and 12-month-old (B) APP+PS1 rats. Brown staining indicates areas that reacted with the 6E10 antibody. The area of diffuse (C) and dense (D) Aβ plaques in the hippocampus was determined by dividing the plaque area by the hippocampal area. S, Sham operated group; O, Ovariectomized group; 12M, 12 months of age; 17M, 17 months of age. p-values for the main effects of genotype (Tg) and age are displayed in each graph.
Glia cells
The APP+PS1 rats had numerically more microglia cells that reacted with the Iba antibody compared to APP21 and WT rats (p = 0.08). There was no age (p = 0.52) nor ovariectomy effect (p = 0.80) on Iba+ microglia cells (Fig. 5). Astrocytes in the hippocampal area that were stained with the GFAP antibody were not affected (ps > 0.1) by transgene, age, nor ovariectomy (data not shown).
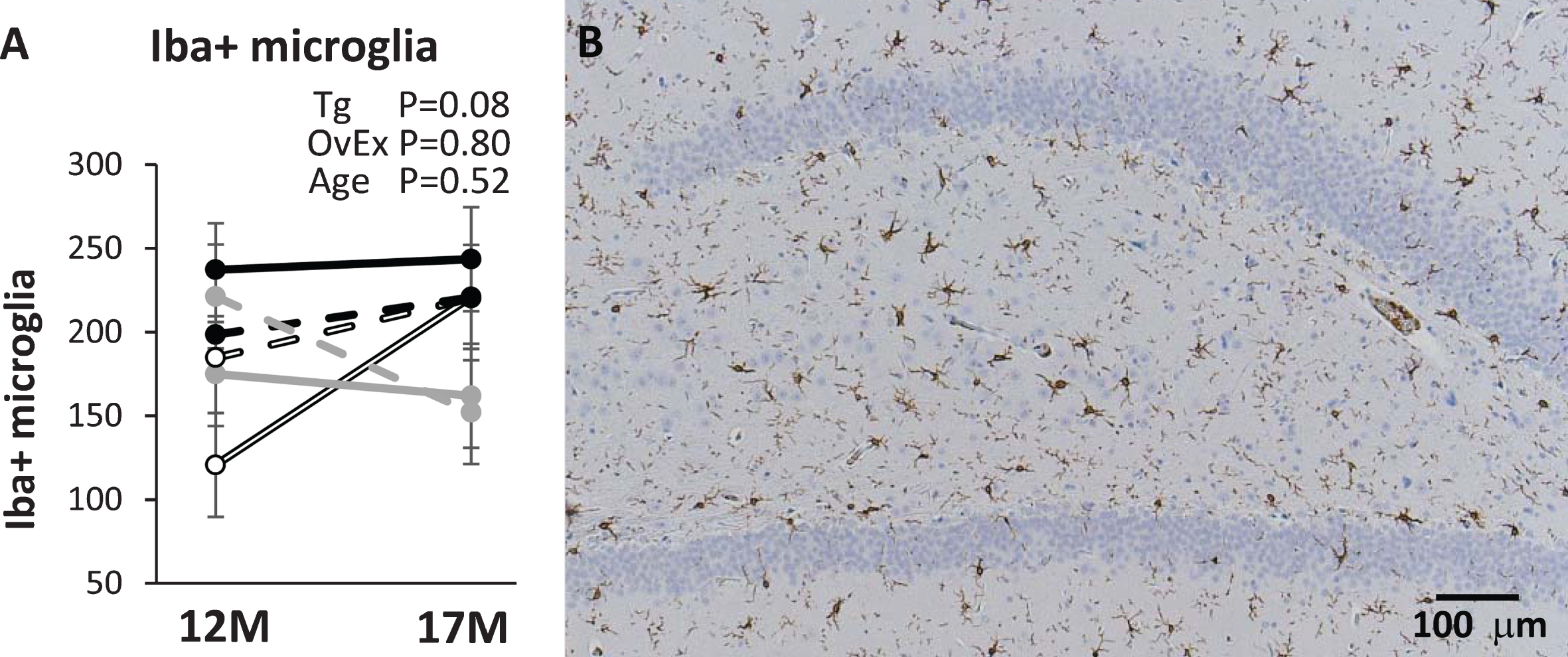
A) Number of microglia cells stained with iba antibody in CA3 region of the hippocampus in 12- and 17-month-old APP21 (open black), APP+PS1 (black), and WT (gray) rats. Solid lines represent sham-operated and dashed lines represent ovariectomized rats. 12M, 12 months of age; 17M, 17 months of age. p-values for the main effects of genotype (Tg), ovariectomy (OvEx), and age are displayed in the graph. B) CA3 region of an APP+PS1 rat stained with Iba antibody.
DISCUSSION
This study was designed to determine the longitudinal effects of ovariectomy in rat models of AD. Ovariectomy was used as an accelerated model for menopause in humans in relation to cognitive and biochemical changes due to ovarian hormone deficiency. There has been remarkable progress in the treatment of cancer after the introduction of novel chemo- and radiotherapy protocols. However, these treatments alone or in combination with oophorectomy procedure causes premature menopause and leads to serious disorders such as obesity, osteoporosis, heart diseases, as well as, cognitive impairment due to sudden or gradual decline in ovarian hormones. On the other hand, ovariectomy at young age may not be an optimal approach for modeling the conditions where gradual decline in ovarian hormones in older age takes place. However, ovariectomy in young animals may be more practical to reduce time and costs of experiments that investigate naturally occurring ovarian hormone decline. Behavior was assessed using the Barnes maze 5 months after ovariectomy and showed that estrogen depletion had impaired learning and cognitive shifting (flexibility) only in the APPOvEx group compared to the APPSham group. The APP21 rats showed behavioral inflexibility in combination with white matter microgliosis, which has also been observed in early (pre-diagnosis) stages of AD as reported by Levit and colleagues [25]. Longer latencies observed in the OvEx rats maybe be due to the slightly heavier body weights of the OvEx animals. The APPOvEx, PS1Sham, and PS1OvEx groups had comparable levels of learning and memory performance which were significantly lower than APPSham, WTSham, and WTOvEx groups. Thus, ovarian hormone depletion effects on cognition varied based on the genotypes tested in the current study. These findings indicate a complex interaction of ovarian hormone levels and genotype on cognitive abilities. Ten and 15 months following ovariectomy when the rats were 12 and 17 months old, the long-term effects of ovarian hormone depletion on molecular and histological changes in the brain were determined. Reproductive performance of rats normally begins to decline at 12 months of age [4] and thus the behavioral assessments in this study were conducted when the sham-operated rats had high levels of circulating reproductive hormones whereas ovariectomized rats did not. A comparison of the brains at 12 and 17 months allowed for the long-term effects of ovariectomy to be assessed. However, it should be noted that after 12 months of age, gonadal estrogen production in the intact rats is expected to be markedly declined and may be comparable to that of ovariectomized rats; this decrease is expected to be even more evident at 17 months of age [4].
The APP+PS1 rats had very low amounts of Aβ40 levels in their brain cortices. TBS soluble Aβ40 was not measurable and formic acid soluble Aβ40 did not increase with age. In contrast, FA soluble Aβ42 levels were much higher in 17-month-old APP+PS1 rats. Furthermore, compared with 12-month-old rats, 17-month-old APP+PS1 rats had 57-fold greater FA soluble Aβ42 levels. Although not statistically significant, ovariectomized rats accumulated over 30% more Aβ42. In contrast, serum Aβ40 levels were as much as 14-fold greater for APP21 rats and 4-fold greater than Aβ42 levels for APP+PS1 rats. Greater serum Aβ40 and very low brain Aβ40 levels indicate that both APP21 and APP+PS1 rats had very little endogenous PS1 in their brains and suggest that the combination of human AβPP with the Swedish and Indiana mutations and PS1 with the L166P mutation leads to the generation of substantially greater amounts of Aβ42 in the brain. In contrast, the serum of both APP21 and APP+PS1 rats had greater amounts of Aβ40 compared to Aβ42, indicative of greater amounts of endogenous PS1 in extracerebral tissues which led to a greater levels of serum Aβ40. These observations indicate an extracerebral origin of serum Aβ40.
We tested levels of 12 mRNAs known to be involved in Aβ metabolism, synaptic and cognitive function, and signal transduction. The only mRNA that was significantly downregulated due to ovariectomy was Grin1 which encodes the zeta subunit of NMDA receptors and plays a role in the plasticity of synapses and is involved in memory and learning. In addition to reduced expression of Grin1 in ovariectomized rats, levels of Grin1 were lower in 17-month-old rats compared with 12-month-old rats. Furthermore, sham-operated APP+PS1 rats had the greatest levels whereas the ovariectomized APP+PS1 rats had the lowest levels of Grin1 expression. Similarly, Grin1 expression was also found to be reduced in an ovariectomized AD mouse model [21]. Interestingly, levels of Bace1, which cleaves AβPP at the beta site and which is required for the generation of Aβ peptides, was observed to be highly correlated with Grin1.
Levels of mRNA encoding Chat, which catalyzes biosynthesis of acetylcholine, decreased with age in all the experimental groups. Interestingly Chat expression was greatest in APP+PS1 rats at 12 months as well as sham operated APP+PS1 rats at 17 months, but sharply reduced in 17-month-old ovariectomized APP+PS1 rats. Similar results were reported by Gibbs [26], which found that Chat mRNA levels decreased in older ovariectomized rats compared with their younger counterparts. The increase in mRNA levels of Chat in APP+PS1 rats who also had high levels of Aβ42 is consistent with the influence of estrogen on cholinergic neurons [27] and activation of Chat with Aβ42 peptides [28]. We also showed that Chat mRNA levels negatively correlated with FA soluble Aβ42 which is consistent with reduction of Chat levels in patients as dementia progresses [29]. Although we did not test the cognitive ability of 17-month-old rats, based on the higher levels of Aβ peptide accumulation in the ovariectomized APP+PS1 rats, it is expected that older rats would have had even greater cognitive decline.
The mRNA encoding Adam9, which is involved in AβPP cleavage [30, 31], decreased in all the experimental groups at 17 months of age, with the exception of the APPOvEx and PS1OvEx groups. Estrogen has been previously shown to induce Adam9 expression in cultured neural cells [32]. The expression pattern of mRNAs involved in AβPP cleavage, Adam9 and Psenen, as well as Aβ degrading enzymes neprilysin (Mme) and Ide were all found to be highly correlated. Although the RNA was obtained from the brain cortex, levels of these mRNAs were correlated with dense Aβ plaques in the hippocampus, indicating that high levels of these mRNAs lead to greater Aβ deposition and degradation. Compared to expression at 12 months, expression of these mRNAs increased with age only in ovariectomized APP21 and APP+PS1 rats at 17 months and decreased in all the other 17-month groups. Conversely, another AβPP cleaving enzyme, Bace1, decreased in all 17-month experimental groups, with ovariectomized APP21 and APP+PS1 exhibiting the lowest expression. Real-time RT-PCR revealed interesting expression patterns and showed differences in gene expression due to ovariectomy, genotype, and aging.
Aβ deposition followed a distinct pattern in APP+PS1 rats. Amyloid plaques appeared to start in the subiculum around 12 months of age and progressed to under the ALV; their size also increased with age. All the 17-month-old APP+PS1 rats had a significant amount of Aβ deposition in the subiculum, under the ALV, as well as other areas of the hippocampus and cortex. The subiculum plays a role in the processing of information about space, movement, and memory, in addition to a regulatory role in the inhibition of the hypothalamic-pituitary-adrenal axis [33]. Furthermore, the subiculum receives inputs from the CA1 as well as cortical and subcortical areas including the entorhinal cortex and integrates and processes information before conveying it to permanent storage locations [34]. The development of initial Aβ deposition in the subiculum region may lead to learning and memory deficiency before widespread Aβ deposition in other regions of the hippocampus and cortex. This important role of the subiculum in memory processing may underlie the memory and learning impairments observed in the APP+PS1 animals at 7 months. However, it is important to note that Aβ load often does not correlate with cognitive impairment in humans and animal models [35, 36], which highlights the importance of assessing both behavioral and physiological markers to get a more accurate estimate of disease progression.
In conclusion, ovariectomy led to learning impairments in the APP21 rats. The performance of APPOvEx rats was comparable to APP+PS1 rats with or without ovariectomy. Furthermore, both groups of WT rats performed better regardless of ovariectomy status. Ovariectomy led to a numerical increase in hippocampal Aβ deposition as well as cortex Aβ42 and Aβ40 levels and the only mRNA that was downregulated due to ovariectomy was Grin1. Furthermore, levels of mRNAs encoding proteins that degrade or cleave Aβ and AβPP were found to be highly correlated and their pattern of expression was different in ovariectomized APP21 and APP+PS1 rats. Altogether, these results indicate that the effect of ovariectomy is complex and can differ by genotype.