
Abstract
Current advancements in neurovascular biology relates a mechanoceutics treatment, known as cranial osteopathic manipulation (COM), Alzheimer’s disease (AD). COM could be used as an evidence-based treatment strategy to improve the symptoms of AD if molecular mechanisms, which currently remain unclear, are elucidated. In the present pilot study, using transgenic rats, we have identified COM mediated changes in behavioral and biochemical parameters associated with AD phenotypes. We expect these changes may have functional implications that might account for improved clinical outcomes of COM treatment. Further investigations on COM will be helpful to establish an adjunct treatment for AD.
Keywords
INTRODUCTION
A 2019 World Health Organization report states that an estimated 50 million people are suffering from dementia worldwide and this number will triple by 2050. There are two classes of pharmacological agents currently used to treat Alzheimer’s disease (AD). This includes cholinesterase inhibitors (donepezil, galantamine, and rivastigmine) and an NMDA receptor blocker (memantine). While pharmacologically manipulating neurotransmitter receptor activity is still effective to manage the symptoms of AD, recent knowledge on brain metabolic waste clearance mechanisms [1–4] motivate investigators to explore novel treatment modalities to clear the amyloid-β (Aβ) oligomers, the principle perpetrator of AD pathogenesis. The existence of a CNS lymphatic system and its connection with deep cervical lymph nodes suggests how and why cranial osteopathic manipulation (COM) treatment produces a clinical response when performed for the treatment of various neurological disorders [5–7]. These clinical observations motivate us to study the molecular mechanisms of COM in the mitigation of AD, which is closely associated with age-dependent impairment of brain metabolite clearance.
It has been recently shown in animal models that the peripheral lymphatic circulation can be improved by lymphatic pump treatment, an osteopathic manipulative technique [8, 9]. Along this direction, we hypothesize that COM can improve fluid circulation around the brain that affects protein homeostasis and immune surveillance [10]. A 3D-imaging study reports that the rat brain fourth ventricle (CV4) region is anatomically similar to the human brain CV4 region [11]. Therefore, COM techniques used in humans should be able to be successfully translated to rats to study the molecular mechanisms of mitigation of brain waste clearance-related disorders like AD.
There are numerous caveats in selecting an appropriate animal model for AD, as reviewed here [12]. In the present study, we used a rat model of AD that mimics age dependent cognitive decline, and is more suitably translatable to humans compared to other models [12, 13]. In addition, the size of the rat head is relatively larger than that of a mouse, so that COM can be performed more accurately. On the other hand, aged rats are a suitable model as 95% of late AD is sporadic and only 5% is early familial; however, the aged rats do not represent all AD pathologies. Therefore, TgF344-AD rats are used for the current study. These TgF344-AD rats exhibit age dependent cerebral amyloidosis, gliosis, apoptotic loss of neurons in the cerebral cortex and hippocampus, and cognitive disturbances [13]. Nonetheless, we have also studied the effect of COM in naturally aged 18-month-old rats as another model, as recently published [14].
METHODS
The transgenic rat line TgF344-AD expresses two (K595N, M596L) gene mutations, one in amyloid precursor protein (APPsw) and one in presenilin 1 (exon-9 deleted (PS1ΔE9)), each independent causes of an early onset of familial AD. A hemizygous male rat was purchased (from Dr. Terrence Town’s laboratory, University of Southern California) and bred with a wild type F344 female rat. Pups were genotyped with the appropriate PCR primers to confirm the expression of both the transgenes (genotyping results provided in Supplementary Figure 1). Two groups of rats [untreated (UT) and COM treated] were used for this pilot study. The UT group contained four [16mo.F (uppercase letter represents sex), 15mo.F, 11mo.M, 5mo.M] and the COM group contained 5 (15mo.F, 15mo.F, 11mo.F, 11mo.M, 11mo.M) TgF344-AD rats. COM group rats received cranial osteopathic manipulative therapy every day for seven days and all rats were anesthetized with 1.5–3% isoflurane throughout the COM procedure. UT group rats were also anesthetized to nullify any differential influence of the isoflurane anesthesia procedure. During the COM procedure, the operator applied mechanical pressure over the rat’s occiput, medial to the junction of the occiput and temporal bone, and inferior to the lambda suture, to place tension on the dural membrane around the 4th ventricle. A detailed COM procedure has been recently published [14]. Briefly, a gentle pressure was applied to resist cranial flexion with the aim of improving symmetry in the cranial rhythmic impulse (CRI), initiating a rhythmic fluctuation of the cerebrospinal fluid and improving mobility of the cranial bones and dural membranes. This rhythmic fluctuation is thought to be primarily due to flexion and extension that takes place at the synchondrosis between the sphenoid and basiocciput. The treatment endpoint was achieved when the operator identified that the tissues relaxed, a “still point” was reached, and improved symmetry or fullness of the CRI was felt. Each animal received about seven minutes of COM treatment at each session. At this point, there is no quantitative measure for the pressure used in this treatment.
RESULTS
In order to study the COM induced changes in cognitive function, a Morris water maze (MWM) spatial learning and memory assay was performed every day during the seven-day COM treatment and on day-8. COM was performed morning hours (7-8 am) and MWM assay was done in the afternoon (4–6 pm) hours. Results (Fig. 1) from these experiments indicate significant differences in COM treated rats compared to UT rats, principally suggestive of improved performance. Detailed experimental procedures of MWM, positron emission tomography (PET), and western blots are recently published [14]. On day-8, after MWM assay, animals were transferred to the University of Virginia Molecular Imaging Core Lab to perform positron emission tomography (PET) imaging in live rats to quantify the Aβ plaques using a clinical grade radiotracer, Florbetapir F18. PET imaging results revealed a 20.2% overall numeric reduction in whole brain Aβ signal (Fig. 2). Three regions where the largest numeric reduction in Aβ signal was observed in COM rats were: visual cortex (31.2%), retrosplenial cortex (31.1%), and pituitary gland (24.8%). After PET imaging, rats were euthanized and brain tissue was excised.

COM alters spatial learning and memory phenotypes in TgF344-AD rats. Rats received four days (day1-4, platform visible) of training before probe trials (day5–8, platform invisible). UT and COM treated rats display a significant difference in various phenotypes, as labelled in each panel, associated with spatial learning and memory. The parameters that exhibit a significant difference in at least one of the four probe trial days (day5–8) are presented here. AnyMaze software that recorded Morris water maze assay was used for the analysis and statistics. UT, n = 4; COM, n = 5, *p < 0.05 and ***p < 0.001 of unpaired, two tailed, student t-test. NW, northwest quadrant where escape platform was located.
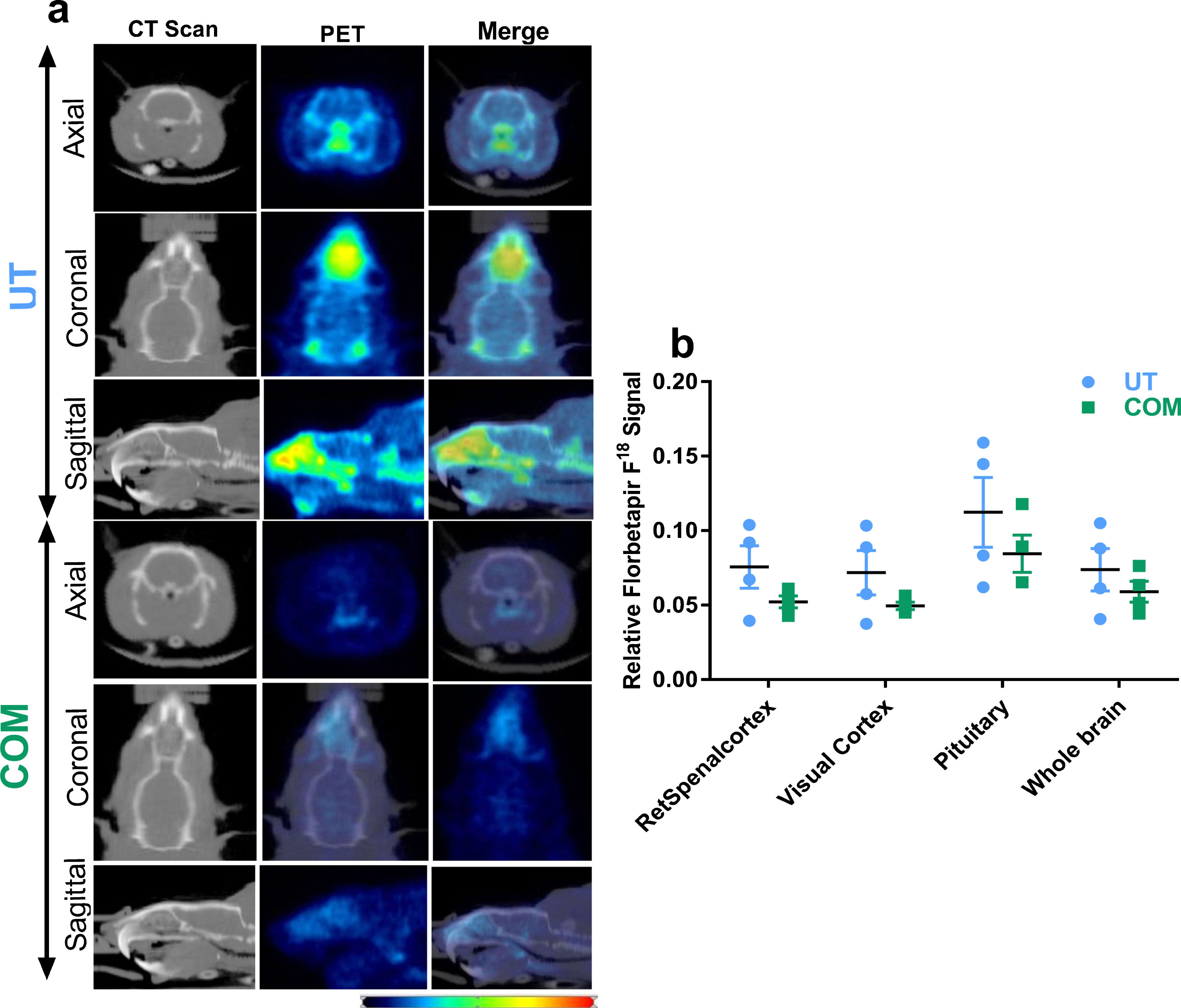
COM numerically reduces Aβ proteins from live rat brain. a) Images show axial, coronal, and sagittal view of CT scan and PET signal obtained from standard uptake curves. Pictures from age matching (11-month-old) male animals, one each from UT and COM group, have been displayed. CT images served as template to analyze PET signals and were used for attenuation correction of PET images. Based on the PET images, volume of interest was identified with the guidance of the Schiffer rat brain atlas provided in the PMOD software (V 3.8, PMOD Technologies LLC, Zurich Switzerland). Thirty-three different brain regions were included and Florbetapir F18 standard uptake values were calculated for each region. b) Histograms represent averaged and normalized signal from whole brain and three regions where the largest numeric reduction in Aβ signal was observed in COM treated rats. UT, n = 4 and COM, n = 4. Blue and red colors in heat map respectively indicate the lowest and highest Florbetapir F18 signal. Signals from non-brain regions (including nostrils and cranial bone) were excluded in the calculations.
To determine the biochemical changes associated with COM treatment, rat brain cerebellar tissue samples were studied for changes in expression of the substrates associated with metabolic waste clearance in UT and COM treated animals using western blots. Due to the close proximity of the cerebellum to the target of the COM procedure, the CV4 region, TgF344-AD rat brain cerebellum lysate was used for the western blot assay. In addition, recent findings suggest that cerebrospinal fluid drainage occurs through the meningeal lymphatic vessels at base of the skull [3]. It was hypothesized that a mechanical pressure applied on the CV4 region can be translated to the brain parenchyma and affect the fluid dynamics in, and upon, this brain tissue. Cerebellar tissue of UT and COM group rat brain were obtained using a brain matrix as reference. Tissue samples were lysed using RIPA buffer with a protease & phosphatase inhibitor cocktail (ThermoFisher Cat# 78440) and protein concentration was determined using Bradford assay. Equal amounts (30 μg) of protein samples from each group were added with appropriate volume of 4X Laemmli (Bio-Rad, cat#1610747). 100 μl of 2-mercaptoethanol was added to every 900 μl of 4X Laemmli used. Samples were boiled at 95–100°C for five minutes before electrophoresed in sodium dodecyl sulfate polyacrylamide gel (10%) and transferred to PVDF (pore size 0.2 μm) membrane. Blots were blocked with 5% nonfat dry milk in tris buffered saline with 0.075% tween 20 and probed with antibodies recognizing the target proteins.
Initially we have studied the changes in expression of aquaporin-4 (AQP4), a subtype of water channel predominantly expressed in the astrocyte end feet processes [15], and two markers for proteins associated with lymphatic vessels [lymphatic vessel endothelial hyaluronic acid receptor 1 (LYVE1) and vascular endothelial growth factor-C (VEGFC)]. Results obtained from these western blots reveal that AQP4, LYVE-1 and VEGFC are significantly elevated in the COM treated rat cerebellum (Fig. 3a, b).
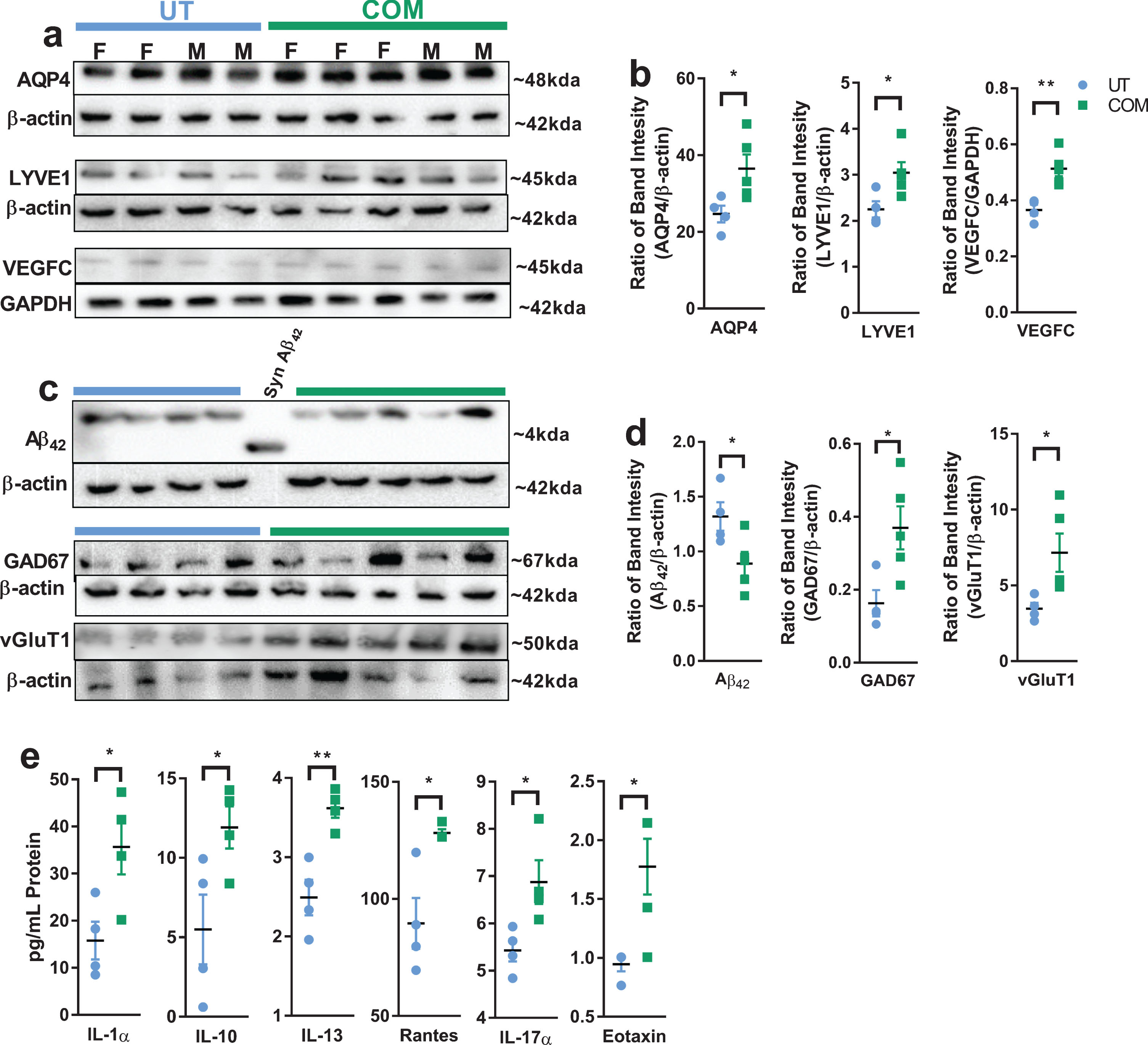
COM changes the expression of biomolecules that are associated with lymphatic drainage and AD phenotype. Each lane was loaded with 30 ug of cerebellar lysate from UT and COM treated animals (gender marked in 3a). The relative protein expression was determined by dividing individual sample band intensity with corresponding β-actin signal obtained from the same blot. Each dot in the figure represents this ratio. Representative western blot pictures (a) of AQP4 (abcam, ab46182), LYVE1 (Invitrogen, PA1-16635), VEGFC (abcam, ab9546) with loading controls (GAPDH or β-actin), and respective dot plots (b) indicate significant increases in expression of proteins associated with CNS fluid dynamics. (c) and (d) represent COM mediated changes in expression of Aβ42 (Cell Signaling, 14974), GAD67 (Millipore, MAB5406), vGluT1, (Synaptic Systems, 135302) that are relevant to AD related cognitive dysfunction. A synthetic rat Aβ42 peptide, labelled as SynAβ42 (Anaspec, AS-25381), was used to confirm Aβ42 antibody specificity. Manufacturer and catalog number of the antibodies used for western bots are italicized. Primary blots were stripped and re-probed with anti-β-actin (or GAPDH) antibody which was used as control. (e) IL-1a, IL-10, IL-13, RANTES, IL-17A, and EOTAXIN levels are elevated in COM treated rat cerebellum lysates. This experiment was performed using Cytokine & Chemokine 22-Plex Rat ProcartaPlex™ Panel assay kit (ThermoFisher, EPX220-30122-901) in MagPix magnetic bead-based multi-analyte panel platform. 20 μg of total protein loaded in each well. UT, n = 4, COM, n = 5, *p < 0.05 and **p < 0.01 of unpaired, two tailed, student t-test. Non-denatured samples were used to facilitate anti-vGluT1 antibody binding as directed by the manufacturer. Subsequently, β-actin was re-probed from the same blot. This might explain the inconsistent β-actin signal in vGluT1 assay.
This observation motivated us to study the changes in the Aβ42 peptide which is a major pathogenic metabolite of amyloid protein, and a reduction of which might indicate a beneficial effect of COM treatment. In addition to Aβ42, two other proteins that are associated with excitatory (vesicular glutamate transporter-1, vGlut1) and inhibitory (glutamate decarboxylate-67, GAD-67) neurotransmission, events relevant to cognitive function that declines in AD, were studied. Western blot results reveal a significant reduction in Aβ42 peptide levels (Fig. 3c, d) in COM treated rats. Previous studies associated the reduction in Aβ proteins with improvement in learning and memory [16, 17]. Increases in GAD-67 and vGluT1 expression suggest COM mediated changes in inhibitory and excitatory neurotransmission, respectively.
To assess the inflammatory mediators which are involved in the AD phenotypes [1], we have studied different cytokines or chemokines using a magnetic bead-based multi-analyte panel platform (MagPix system). Results from this assay revealed a moderate increase in IL-1α, IL-10, IL-13, RANTES, IL-17A, and EOTAXIN levels in COM treated rat cerebellum lysates (Fig. 3e). These changes indicate that the immune system recognized the pressure applied around the cerebellomedullary cistern region.
DISCUSSION
Overall, the results obtained from this pilot study reveal that COM treatment is transferable to TgF344-AD rats to produce changes in behavior and molecules associated with AD. Differences observed in various MWM assay parameters indicate COM mediated improvement in spatial learning and memory. While these observations need to be reconfirmed by more control groups with larger sample size, similarities in MWM data obtained from aged rats [14] and transgenic rats at different time points exhibit reproducibility. At the molecular level, PET imaging of Aβ plaques using florbetapir F18 tracer reveals COM induced reduction in Aβ proteins. Further, western blot results reconfirmed COM induced reduction in a more pathogenic metabolite of Aβ protein, Aβ42. While reporting this we do not ignore the ongoing debate among AD researchers that reduction in Aβ protein is not necessarily a direct indication of improvement in AD symptoms. However, the large number of publications that evolved from decades of intense investigation to identify the role of various Aβ fragments in AD pathogenesis cannot be overlooked. Nonetheless, animal models with mutations in amyloid precursor protein exhibit AD phenotypes [13]. Therefore, after identifying a reduction in Aβ fragments combined with the increased expression of GAD67 and vGluT1 that indicate changes in neurotransmission, it is fair to make a cautious interpretation that reduction in Aβ protein might have influenced the changes observed in spatial learning and memory in the transgenic rats. This leads to an immediate question on the potential mechanism of reduction in Aβ fragments from the rat brain. Although we could not effectively address this question in this study, an increased expression of AQP4, LYVE-1, and VEGFC in COM treated rat cerebellar tissue serve as a trailblazer for future investigations in this direction.
Previous studies reported COM induced skull deflections in live non-human primates, dogs, cats, rabbits, and unembalmed human cadaver heads [18, 19]. These studies reveal that a mechanical pressure can cause cranial deflections up to 910 μM based on the region of the skull [20]. These cranial deflections may influence brain fluid circulation through the glymphatic periarterial influx pathways and meningeal lymphatic perivenous efflux pathways that are running physically in close proximity to the cranial bones [21]. The function of glymphatic and meningeal lymphatic pathways is largely regulated by the water channels (AQP4) expressed in the perivascular astrocyte end-foot processes. Our data shows a significant increase in AQP4 protein expression (Fig. 3a) in the COM treated animals. This finding, together with other results, supports the hypothesis that COM, designed to induce cranial bone deflections, can potentially improve fluid circulation that plays a role in brain waste clearance. Not directly measuring the COM induced changes in CNS fluid dynamics is a major limitation of this pilot study. However, we are currently working to study the rate of fluid clearance using appropriate markers.
Most AD patients are more than 70 years old. This age group of patients may have minimally functioning liver and kidneys that are essential to metabolize and excrete the drugs running through them. In addition, AD patients also suffer from comorbidities including hyperglycemia, hypertension, and dyslipidemia, and these disorders require daily medications throughout life. Co-administration of an additional drug to treat the symptoms of dementia may induce adverse reactions that might outweigh the beneficial effects of the drugs. Therefore, a non-pharmacological adjunct therapy will be favorable to obtain more clinically desirable outcomes. In the current era of economic crisis associated with long-term healthcare for AD patients, we anticipate COM therapy will improve the symptoms with low cost and no adverse effects.
Footnotes
ACKNOWLEDGMENTS
This work was funded by the Research Eureka Accelerator Program (REAP, Fund # 10261) of the Edward Via Virginia College of Osteopathic Medicine, and American Osteopathic Association (grant#1915733). Authors acknowledge Virginia Tech Teaching & Research Animal Care Support Service (TRACSS) for taking care of TgF344-AD rats. The PET scanner at UVa was purchased with funds from NIH grant 1 S10 OD021672.