
Abstract
Background:
Accumulation of p25 is thought to be a causative risk factor for Alzheimer’s disease (AD). As a cleaved product of p35, p25 binds to cyclin-dependent kinase 5 (Cdk5) and leads to the hyperactivity of Cdk5. Then, Cdk5/p25 phosphorylates many pathological substrates related to neurodegenerative diseases. p25 transgenic (Tg) mouse model recaptures some pathological changes of AD, including tau hyperphosphorylation, neurofibrillary tangles, neuroinflammation, and neuronal death, which can be prevented by transgenic expression of Cdk5 inhibitory peptide (CIP) before the insult of p25.
Objective:
In the present study, we would like to know whether adeno-associated virus serotype-9 (AAV9)-mediated CIP can protect neurons after insult of p25 in p25Tg mice.
Methods:
Administration of AAV9-CIP or control virus were delivered in the brain of p25Tg mice via intracerebroventricular infusions following the induction of p25. Western blotting, immunohistochemistry and immunofluorescence assessment, and animal behavioral evaluation were performed.
Results:
Brain atrophy, neuronal death, tau phosphorylation and inflammation in the hippocampus, and cognitive decline were observed in p25Tg mice. Administration of CIP but not the control virus in p25Tg mice reduced levels of tau phosphorylation and inflammation in the hippocampus, which is correlated with inhibition of brain atrophy and neuronal apoptosis in the hippocampus, and improvement of cognitive decline.
Conclusion:
Our results provide further evidence that the neurotoxicity of p25 can be alleviated by CIP.
INTRODUCTION
Dysregulation of cyclin-dependent kinase 5 (Cdk5), a proline-directed serine/threonine protein kinase, plays a causative role in many neurodegenerative diseases, including Alzheimer’s disease (AD). Cdk5 is essential for the development of the mammalian central nervous system [1, 2]. The activity of Cdk5 is dependent on its binding to p35 or p39 [3–7]. p35 would be cleaved into p25 in some neurotoxic conditions, such as oxidative stress, amyloid-β (Aβ) peptide stimulation, and trauma [8–13]. As an additional activator of Cdk5, p25 also binds to Cdk5, forming the Cdk5/p25 complex with a wider subcellular localization and a longer half-life than Cdk5/p35, which causes hyperphosphorylation of many downstream substrates related to degenerative diseases [14, 15]. In postmortem examination of AD patients, accumulated p25 and aberrant activity of Cdk5 were found in the neurons containing neurofibrillary tangles (NFT) of AD brains [9]. Overexpression of p25 in the brain of mice recaptures some pathological characteristics of AD, including hyperphosphorylated tau, NFT, deposition of Aβ plaques, and neuroinflammation [16–18]. These studies suggest that p25 is one of the causative risk factors for pathological changes and loss of neurons related to neurodegenerative diseases [16, 19–21].
Targeting neurotoxicity of p25 by Cdk5 inhibitors in different neurodegenerative disease models has been a potential neuroprotection strategy. Inhibitory peptides derived from p35, such as p5 or Tat peptide modified p5 (TFP5, T, transactivator of transcription, F, fluorescein isothiocyanate) have been shown to inhibit the activity of the Cdk5/p25 complex in 5XFAD-Tg mice, which express high levels of mutant APP (KM670/671NL, I716V, V717I) and PSEN1 (M146L and PSEN1), MPTP-insulted mice, a Parkinson’s disease (PD) model, and p25Tg mice [22–24]. Transgenic expression of CIP before the induction of p25 in p25Tg mice have also been proved to be neuroprotective [17]. Previously, we have demonstrated that AAV9-mediated CIP protects neurons and alleviates behavioral abnormalities in APP/PS1 Tg mice, MPTP-insulted mice, and p25 overexpression mice delivered by AAV8-GFP-p25 [18, 25]. In the present study, we further investigated whether AAV9-mediated CIP exhibited neuroprotection following the induction of p25 in p25Tg mice. The results showed that intracerebroventricular (ICV) infusions of AAV9-CIP-T2A-mCherry reduced tau phosphorylation, prevented weight loss, inhibited neuronal death, and attenuated cognitive decline in p25Tg mice. These data provided evidence that the neurotoxicity of p25 can be alleviated by AAV9-mediated CIP, which may be a promising potential therapy for neurodegenerative diseases.
MATERIALS AND METHODS
Animals
Two Tg mouse lines, C57BL/6-Tg (tetO-CDK5R1/GFP) mice were purchased from Jackson Laboratory (CA, USA). B6;CBA-Tg (CamkII-tTA) mice were provided by Animal Core Facility of Nanjing Medical University (Nanjing, China). TetO-CDK5R1/GFP Tg mice were crossed with CamkII-tTA Tg mice, resulting in p25 double Tg mice (p25Tg), which express p25 in the forebrain [16]. Genotyping was performed for each mouse. In the p25Tg mice, inhibition of p25 expression was achieved by providing water containing the tetracycline derivative doxycycline (Dox). Once Dox was withdrawn, the p25Tg mice started expressing p25 in the forebrain [16]. Mice were derived from heterozygote crosses for all genotypes. Animals were housed in standard cages under 12 h light/dark cycle (light time for 8 am to 8 pm, dark time for 8pm to 8am), standard conditions of temperature (∼22°C) and humidity (30% ∼50%). All mice used in this study were conceived and raised in the presence of Dox (up to 200 μg/ml in water, changed thrice a week) for at least 8 weeks postnatally.
All animal experiments were performed according to the protocols approved by the University Committee on Animal Care of Southern Medical University, Guangzhou, China and conducted in accordance with the Guide for the Care and Use of Laboratory Animals.
AAV9 vectors preparation
AAV9 vectors were produced by Hanbio Biotechnology (Shanghai, China). Briefly, the 3′-flag-tagged CIP was generated by PCR with pcDNA-p35 as a template. Primers used included: 5′- GAGCGCAGTCGAGAAGCCACCATGTGCCTGGGTGAGTTTCT-3′ and 5′-TAGTGCTAGCGGATCCTTT
Administration of AAV virus
There were four experimental animal groups: 1) wild-type mice (WT group), 2) p25Tg mice (p25 group), 3) p25Tg mice infused by ICV with 2 μl AAV9-mCherry (CT group), 4) p25Tg mice infused by ICV with 2 μl AAV9-CIP (CIP group). Mice at the age of 8 weeks were anesthetized with 2% pentobarbital sodium (40 mg/kg; RWD, Shenzhen, China). The stereotaxic coordinates for ICV infusions were 1.0 mm lateral, 0.5 mm posterior to the bregma, and 2.5 mm deep from the dura [26]. Mice were deeply anesthetized by 2% pentobarbital sodium (120 mg/kg; RWD, Shenzhen, China) and euthanized at 5 weeks after ICV infusions (age of 13 week) or at the end point (age of 24 week). Brain samples were obtained for different assessments.
Western blot analysis
The hippocampus lysate was prepared routinely as described previously [27]. Primary antibodies (Abs) used were anti-GFP Ab (Aves Labs, Stuttgart, Germany), anti-mCherry Ab (HanBio, Shanghai, China), anti-β-actin Ab (Proteintech, Chicago, IL), anti-GFAP Ab (Millipore, Billerica, MA), anti-cleaved Caspase 3 Ab (Asp175, Cell Signaling Technology, MA, USA), and several tau Abs from Thermo Fisher Scientific Life Sciences (Waltham, MA, USA), including total tau (TAU-5) and phospho-tau (Thr231), AT180, phospho-paired helical filament-tau (Ser202/Thr205) AT8, S199 (Ser199). All primary Abs and horseradish-peroxidase-labeled secondary Abs (Boster, Wuhan, China) were diluted at 1 : 1000. Signals were detected by Kodak In-vivo Imaging System FX Pro suite (Kodak, Rochester, MN, USA).
Immunohistochemistry and immunofluorescence
Immunohistochemistry and immunofluorescence were performed as described previously [27, 28]. Briefly, each hippocampus was dissected from the anesthetized mouse and fixed in 4% paraformaldehyde for paraffin section. Paraffin coronal brain sections for immunohistochemistry and immunofluorescence were incubated with different Abs: anti-NeuN (Cell Signaling Technology), anti-GFP (Aves Labs, Davis), anti-flag (Proteintech, Chicago, IL, USA), anti-GFAP (Proteintech), and anti-Cleaved Caspase 3 (Asp175, Cell Signaling Technology). Secondary Abs were goat against rabbit IgG (ZSGB-BIO Beijing, China), goat anti-chicken IgG/Alexa Fluor 488 (Bioss, Boston, MA, USA) and goat anti-mouse IgG/Alexa Fluor 594 (Bioss). Cell nuclei were stained by DAPI (Boster, China). The number of neurons in CA1 region of hippocampus of three sections were counted blindly by independent investigators. Images were taken under an inverted system microscope (Olympus, Tokyo, Japan).
Behavioral studies
Mice of 12 weeks were housed in the test room before testing. All tests were performed consecutively as described previously.
Morris water maze
The Morris water maze test (MWM) was conducted as reported previously [29] to evaluate visual-spatial learning and memory. The test pool was circular, 200 cm in diameter, 75 cm in height and contained 25 cm in depth of water. A circular platform with a diameter of 10 cm was set in the middle of the goal quadrant. The swimming activity of each mouse was recorded using a camera mounted above the pool. On day 1, each mouse was trained to reach the visible platform three times. On days 2–4, during the hidden platform test, we introduced the intensive training protocol, which included 12 trials per day in 3 blocks of 4 trials each for 3 days. On day 5, each animal was given a 90 s probe trial to evaluate its ability of memory retention.
Elevated plus maze
The elevated plus maze consists of two open arms (35×5 cm), two enclosed arms (35×5×20 cm), and one open square (5×5 cm) in the center. The maze was 50 cm above the floor. Each mouse was placed in the central area with the head facing the closed arm and allowed to explore the maze for 10 min, during which the total time spent in the open arms was recorded [29].
Statistics
All data were presented as means±SD. The results of hidden platform test were analyzed by repeated measures ANOVA. Number of neurons was analyzed by nonparametric test. All other data were carried out using t-test using GraphPad 5 (GraphPad, La Jolla, CA). A p-value <0.05 was considered to be statistically significant.
RESULTS
AAV9 vectors delivered CIP in the mouse brain via ICV injections
We firstly assured to induce expression of p25 in p25Tg mice. As described, expression of the p25-GFP fused protein was controlled by tetracycline transactivator (tTA), which was driven under the CamkII promotor [16]. In the presence of the tetracycline derivative Dox, the expression of p25 was inhibited. In this study, all mice were raised in the presence of Dox for at least 8 weeks postnatally. As shown in Fig. 1A, when Dox was added to the water, only a trace amount of p25-GFP (50 kDa) was found in the brains of p25Tg (p25Tg off). High levels of p25-GFP expression were observed in the cortex and hippocampus after induction only by replacing the Dox-containing water with regular water for 2 weeks (p25Tg on), which demonstrated that we were able to successfully control the inducible system. We then assessed the expression of AAV9 vectors after ICV injections. As illustrated in Fig. 1B, injection of virus was performed at week 8 following induction of p25, and different assays were analyzed at week 12, 13, or 24. Both AAV9-mCherry and AAV9-CIP-T2A-mCherry expressed mCherry in the brain of mice as indicated by western blot analysis five weeks after injection of virus (Fig. 1C). The expression of flag-tagged CIP was detected in the hippocampus by immunofluorescence using anti-flag antibody (Fig. 1C). The data indicate that AAV9 vectors can deliver CIP in the mouse brain.
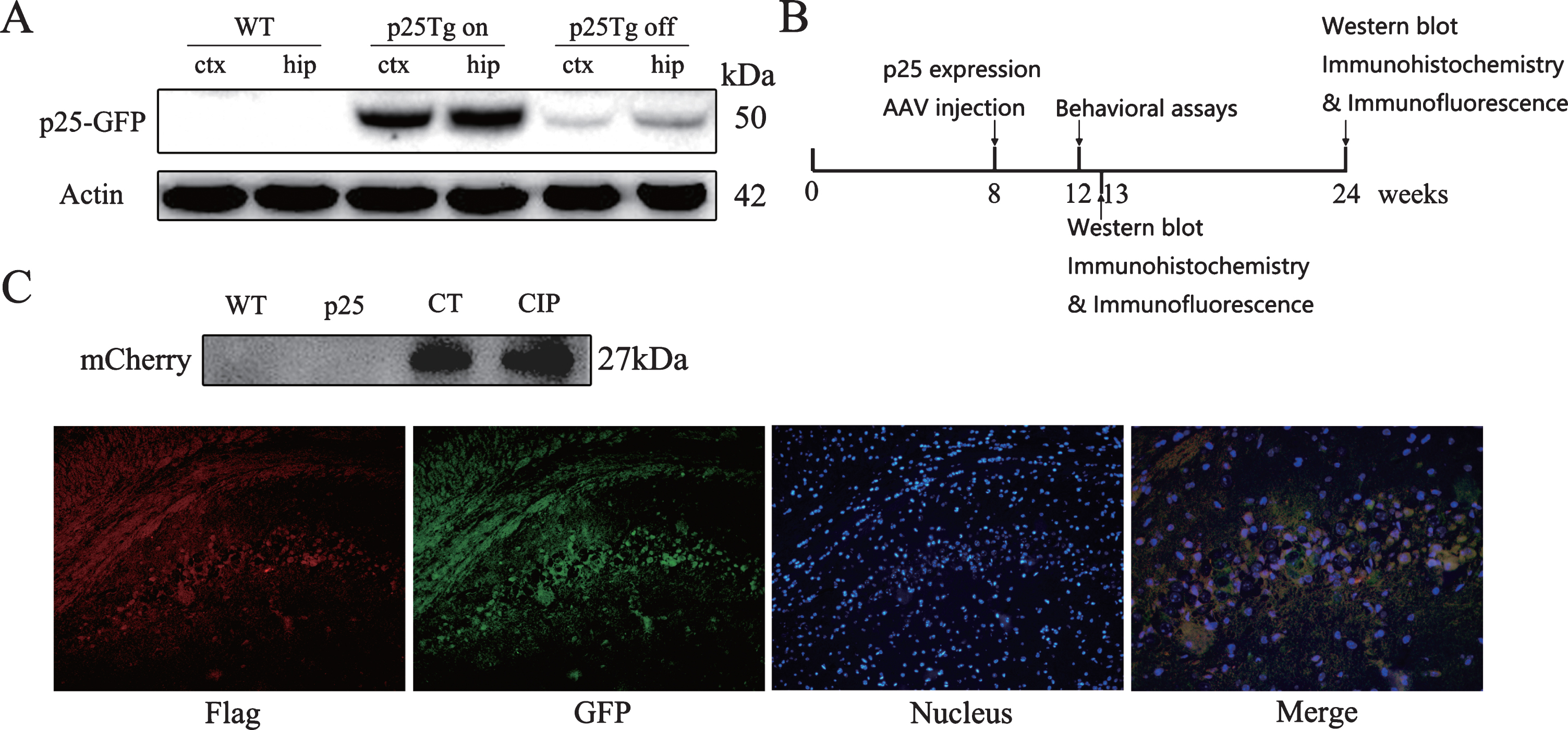
Expression of p25-GFP and CIP in the brain of p25Tg mice. A) Induction of p25 expression in p25Tg mice. In the presence of the tetracycline derivative doxycycline (Dox), the expression of p25-GFP in p25Tg was inhibited (p25Tg off). Two weeks after the induction by replace Dox with water (p25Tg on), a high level of p25-GFP fusion protein were detected in the cortex (ctx) and hippocampus (hip). B) The graph shows the time points of p25 expression, treatment of AAV9 vectors, and analysis. The expression of p25 was started at week 8. CIP group mice and CT group mice received AAV9-CIP or AAV9-mCherry at week 8. Behavioral tests were assessed at week 12. Five weeks after ICV injection (week 13) or at the end point (week 24), mice were deeply anesthetized and euthanized. Brain samples were obtained for different assessments. C) Western blot analysis of mCherry indicated that AAV9-mCherry and AAV9-CIP expressed mCherry in the brain of mice. Immunofluorescence of flag-tagged CIP or p25-GFP in CA1 of the hippocampus was detected by using anti-flag or anti-GFP Abs, respectively. Red: flag-tagged CIP, green: p25-GFP, blue: DAPI staining of nucleus.
AAV9-CIP prevented weight loss and neuronal death in p25Tg mice
As shown in Table 1, the body weights of male and female p25Tg mice decreased significantly when compared with the WT mice, while AAV9-CIP treatment significantly increased the body weights of p25Tg mice when compared with that in the CT group. Control virus had no effect on the weight of p25Tg mice.
The body weight of mice in different groups
Note. X, XX: Means of mice weight in the same row with different superscripts differ significantly at p < 0.05 (in the comparison of the same gender between WT group and p25 group). Y, YY: Means of mice weight in the same row with different superscripts differ significantly at p < 0.05 (in the comparison of the same gender between CT group and CIP group).
In order to evaluate whether AAV9-CIP would protect neurons from the neurotoxicity induced by p25, we firstly measured the brain weights of mice at weeks 13 and 24. As showed in Fig. 2B, the brain weights of p25Tg mice gradually decreased, compared with WT mice. AAV9-CIP treatment prevented brain atrophy at week 13 and alleviated the brain atrophy at week 24 when compared with AAV9-mCherry treatment. We then used anti-NeuN Ab for immunostaining of the neurons in the CA1 region of the hippocampus at week 13 and 24. As indicated in Fig. 2C, the number of neurons of the p25Tg mice decreased significantly at week 13 and 24, compared with that in the WT mice. AAV9-CIP treatment prevented neuronal loss in p25Tg mice at week 13 and 24 when compared with AAV9-mCherry treatment. These results suggest that ICV injection of AAV9-CIP prevents weight loss, brain atrophy, and neuronal loss in p25Tg mice.

AAV9-CIP prevented loss of body weight, brain atrophy, and neuronal loss in p25Tg mice. A) Comparison of the body weight of male and female mice in different groups from week 8 to week 24. B) Compared with WT mice, brain atrophy was obvious in p25Tg mice. The brains of CIP group mice were larger than that in CT group mice at week 24. The brain weight of p25Tg mice (week 13 : 0.3934±0.0612 g, n = 8; week 24 : 0.3337±0.0218 g, n = 6) decreased significantly when compared with WT mice (week 13 : 0.4833±0.0250 g, n = 8, **p < 0.01; week 24 : 0.4888±0.0194 g, n = 6, **p < 0.01). The brain weight of CIP group mice (week 13 : 0.4431±0.0441 g, n = 8; week 24 : 0.3932±0.0227 g, n = 6) were heavier than CT group mice (week 13 : 0.3861±0.0601 g, n = 8, *p = 0.0484; week 24 : 0.3395±0.0261 g, n = 6, *p < 0.01). Scale bars, 100 μm. C) Immunohistochemistry of hippocampus with anti-NeuN Ab for each group at week 13 and week 24. Number of neurons was calculated by counting NeuN-positive cells in the CA1 region of hippocampus. Number of neurons decreased significantly in the p25Tg mice (week 13 : 297.8±40.42, n = 3; week 24 : 193.2±34.67, n = 3) when compared with that in the WT mice (week 13 : 484.6±35.47 g, n = 3, **p < 0.01; week 24 : 489.6±43.57, n = 3, **p < 0.01). AAV9-CIP treatment (week 13 : 407.6±46.68, n = 3; week 24 : 350.7±48.43, n = 3) inhibited neuronal loss when compared with AAV9-mCherry treatment (week 13 : 273.3±39.71, n = 3, *p < 0.01; week 24 : 190.9±33.49, n = 3, *p < 0.01).
AAV9-CIP inhibited tau hyperphosphorylation in p25Tg mice
Tau hyperphosphorylation triggers formation of NFTs, which is one of the hallmarks of pathological changes in AD. Tau hyperphosphorylation is also one of the pathological changes in p25Tg mice, which is inhibited by transgenic expression of CIP before the overexpression of p25 [16, 17]. In the present study, western blot analysis showed that the level of tau phosphorylation in p25Tg mice increased significantly, as shown in staining with AT180 (Thr231), AT8 (Ser202/Thr205), S199 (Ser199) Abs at week 13 and 24, which are among some of the most active phosphorylation sites of tau. At week 13, tau phosphorylation detected by AT8, AT180, and S199 Abs were all reduced in mice of CIP group compared to mice in CT group (Fig. 3A). At week 24, tau phosphorylation at Thr231, but not at Ser199 and Ser202/Thr205, was still significantly lower in the CIP group compared with that in the CT group (Fig. 3B). These data show that AAV9-CIP treatment inhibits tau hyperphosphorylation in p25Tg mice.

ICV injection of AAV9-CIP inhibited tau phosphorylation, reduced the level of GFAP and Cleaved Caspase3 in p25Tg mice. A) Western blot analysis of tau phosphorylation from 4 groups at week 13. The level of tau phosphorylation in p25Tg group increased significantly when compared with WT group. ICV injection of AAV9-CIP reduced the tau phosphorylation in CIP group (week 13: S199 : 0.3226±0.1549, n = 3; AT8 : 0.5355±0.1079, n = 3; AT180 : 0.5084±0.0658, n = 3) at Thr231, Ser202/Thr205, and Ser199 when compared with CT group (week 13: S199 : 0.8406±0.2618, n = 3, *p < 0.01; AT8 : 1.0400±0.0942, n = 3, *p < 0.01; AT180 : 0.8296±0.1131, n = 3, *p = 0.013). B) Western blot analysis of the tau phosphorylation from 4 groups at week 24. ICV injection of AAV9-CIP (week 24: S199 : 0.7768±0.0938, n = 3; AT8 : 0.9842±0.1788, n = 3; AT180 : 0.4496±0.1204, n = 3) reduce the tau phosphorylation at Thr231 when compared with CT group (week 24: S199 : 0.9114±0.0839, n = 3, NSp = 0.138; AT8 : 1.1770±0.2208, n = 3, NSp = 0.304; AT180 : 0.8467±0.1244, n = 3, *p = 0.017). C) Immunofluorescence images of CA1 region of hippocampus and western blot analysis of GFAP and Cleaved Caspase 3 at week 13. The level of GFAP and Cleaved Caspase 3 were increased in p25Tg mice compared to WT mice. CIP group mice (week 13: GFAP: 0.4333±0.0518, n = 3; Cleaved Caspase 3 : 0.5436±0.1338, n = 3) had lower level of GFAP and Cleaved Caspase 3, when compared with CT group mice (week 13: GFAP: 0.6823±0.0575, n = 3, *p < 0.01; Cleaved Caspase 3 : 0.7844±0.0552, n = 3, *p = 0.045). D) Immunofluorescence images of CA1 region of hippocampus and western blot analysis of GFAP and Cleaved Caspase 3 at week 24. CIP group mice (week 24: GFAP: 0.3818±0.0677, n = 3) had lower level of GFAP compared to CT group mice (week 24: GFAP: 0.5733±0.0892, n = 3, *p = 0.042), while the level of Cleaved Caspase 3 had no difference between CIP group mice (week 24: Cleaved Caspase 3 : 0.5179±0.0716, n = 3) and CT group mice (week 24: Cleaved Caspase 3 : 0.6328±0.0559, n = 3, NSp = 0.093).
AAV9-CIP reduced the level of GFAP and cleaved caspase3 in p25Tg mice
Previous studies have shown that the levels of GFAP and cleaved caspase 3 (CC3), markers for astrocytosis and apoptosis, respectively, are increased in p25Tg mice, which are reduced by pre-expression of CIP in p25Tg mice [16, 30]. To evaluate whether AAV9-CIP can reduce levels of GFAP and CC3 after p25 induction in p25Tg mice, we used immunofluorescence and western blot analysis to detect the levels of GFAP and CC3 at week 13 and week 24. At week 13, the levels of GFAP and CC3 in p25Tg were higher than that in WT mice, while the levels of GFAP and CC3 in CIP group were lower than that in CT group (Fig. 3C). At week 24, AAV9-CIP treatment still reduced the level of GFAP in CIP group when compared with CT group (Fig. 3D). In addition, we detected microglia by immunohistochemistry using anti-Iba1 Abs, and there was no difference between WT mice and p25Tg mice (data not shown). These results suggest that ICV injection of AAV9-CIP reduces astrocytosis and prevents apoptosis caused by p25 expression.
AAV9-CIP reversed behavioral changes in p25Tg mice
To assess whether AAV9-CIP treatment would alleviate the behavioral changes in p25Tg mice, we performed the elevated plus maze test and Morris water maze test at week 12. Elevated plus maze is based on the aversion of mice to open spaces and used to evaluate the anxiety-like behavior of mice. p25Tg mice spent more time in the open arms compared to WT mice, while mice in the CIP group spent less time in the open arms than mice in the CT group (Fig. 4B). There was no difference in time spent in the close arms between CIP group and CT group (Fig. 4B). These results imply that p25 expression leads to anxiety-like or aversion changes in p25Tg mice and the administration of AAV9-CIP alleviates these changes.

ICV injection of AAV9-CIP prevented the behavioral changes and alleviated cognitive deficits in p25Tg mice. A) The track of mice in elevated plus maze of WT mice, p25Tg mice, CT mice, and CIP mice at week 12. B) Graph shows that p25Tg mice (241.3±126.5 s, n = 9) spent more time in open arms when compared with WT mice (69.4±57.5 s, n = 14, **p < 0.01), while in CIP mice (84.7±66.2 s, n = 8), the time spent in open arms was decreased when compared with CT mice (216.0±126.1 s, n = 8, *p = 0.021). There was no difference between CIP group mice (428.5±75.4 s, n = 8) and CT group mice (328.0±126.9 s, n = 8, NSp = 0.075) on the time spent in close arms. C) The track of mice in explore test of Morris water maze test from 4 groups at week 12. D) In hidden platform test, at trial 9, p25Tg mice (27.00±14.7 s, n = 8) had longer latency to find hidden platform compared to WT mice (9.66±4.11 s, n = 14, **p < 0.01), while CIP mice (14.83±7.45 s, n = 8) had shorter latency compared to CT mice (36.68±14.69 s, n = 8, *p < 0.01). E) In explore test, p25Tg mice (22.15±7.09 s, n = 8) spent less time in the goal quadrant when compared with WT mice (38.53±8.05 s, n = 14, **p < 0.01). AAV9-CIP treatment (39.65±6.86 s, n = 8) increased the time p25Tg mice spent in goal quadrant when compared with AAV9-mCherry treatment (22.67±12.55 s, n = 8, *p < 0.01). The times of p25Tg mice crossed platform area (3.00±1.31, n = 8) were lower than WT mice (7.14±2.57, n = 14, **p < 0.01). AAV9-CIP treatment (5.88±1.64, n = 8) increased the times p25Tg mice crossed platform area when compared with AAV9-mCherry treatment (2.63±2.07, n = 8, *p < 0.01). The total swimming distance had no difference between WT mice and p25 mice, while CIP mice had no difference with CT mice (WT: 2302±1012 cm, n = 14; p25 : 2062±1075 cm, n = 8, NSp = 0.606; CT: 2165±899.3 cm, n = 8; CIP: 2226±760.9 cm, n = 8, NSp = 0.885).
Morris water maze was used to evaluate the spatial memory of mice. As shown in Fig. 4D, in the hidden platform test, p25Tg mice took more time to find the invisible platform compared with WT mice. AAV9-CIP treated p25Tg mice spent less time to find the invisible platform than AAV9-mCherry treated p25Tg mice. In the explore test, the total swimming distance of mice in p25 group had no difference compared with mice in the WT group, while p25Tg mice spent less time in the target area than the WT mice (Fig. 4E). AAV9-CIP treated p25Tg mice spent more time in the target area, when compared with AAV9-mCherry treated p25Tg mice (Fig. 4E). On the other hand, the number of times p25 mice crossed the platform area were lower than that of WT mice, while the number of times of the CIP mice were higher than that of CT mice. These results indicate that the spatial memory impairment of p25Tg mice is alleviated by the ICV injection of AAV9-CIP.
DISCUSSION
Dysregulation of Cdk5 activity plays an important role in the pathological changes of AD [31, 32]. Aberrant Cdk5/p25 hyperactivity appears in the NFTs of the postmortem brains of AD patients [9]. Inducible expression of p25 in adult mice results in the aberrant activity of proline directed serine/threonine kinases, such as Cdk5 and GSK3β, mimics many pathological changes of AD, and leads to the impairment of memory [16, 31]. Transgenic expression of CIP before the insult of p25 alleviates tauopathy [17]. Here, we further provide evidence that the administration of CIP following induction of p25 in p25Tg mice prevented the weight loss and brain atrophy in p25Tg mice. Tau phosphorylation, neuronal loss, and astrocytosis were inhibited by AAV9-CIP treatment, accompanied by the prevention of cognitive decline. The present study suggests AAV-mediated CIP is a practical therapeutic strategy for the p25 intervention in neurodegenerative diseases.
Although several groups have observed various phenotypes from p25 overexpression Tg mice, most studies support that the aberrant activity of p25 is related to the pathologic changes observed in neurodegeneration [16, 33–35]. In p25Tg mice using rat neuron-specific enolase (NSE) as the promoter, hyperphosphorylation of tau and neurofilaments occurs mainly in the amygdala, hypothalamus, and cortex [35]. When the expression of p25 is under CMV promoter, the features observed in p25Tg mice include neuronal necrosis and the enlargement of the pituitary gland rather than the hyperphosphorylation of tau and neuronal death [20]. Four independent groups have used this inducible strategy to express p25 driven by CamkII-tTA-promoter. Two groups have similar observations in the hippocampus and cortex, reminiscent many features of AD-like pathological changes, including phosphorylation of tau, NFTs, Aβ deposition, neuroinflammation, and neuronal loss [16, 27]. One group found no tau phosphorylation and neuronal apoptosis, but did observe hippocampal sclerosis, neocortical degeneration, and activated microglia [34]. In agreement with the studies conducted by Cruz, Sundaram, and their colleagues [16, 17], our p25Tg mouse line clearly showed significantly increased p25 after induction, tau hyperphosphorylation, astrocytosis, loss of neurons, and shrinkage of brain. However, we did not observe microgliosis after induction of p25 (data not shown). Previously, we found that p25 overexpression mice via AAV8-GFP-p25 infection had tau hyperphosphorylation and neuroinflammation, but no neuronal apoptosis [18]. In the present study, we induced p25 expression 8 weeks postnatally, two weeks later than Sundaram et al. reported. Reasons for the difference among p25Tg mice remained mostly unknown. However, the location and level of p25 expression and the duration for p25 exposure affect the phenotype of p25Tg mice [17]. Transient p25 expression also exhibited a different effect compared to long term expression of p25. Short term expression of p25 enhanced hippocampus-dependent memory, while prolonged p25 expression resulted in cognitive deficits [36]. AAV-mediated p25 overexpression in adult mouse via tail vain injection or ICV injection also displayed a hyperphosphorylation of tau and memory impairment [18, 37].
Targeting the neurotoxicity of p25 in different diseases mouse models has been conducted by pharmacological, transgenic, and AAV mediated interventions [17, 39]. Gene therapy is believed to be a potential treatment for multiple diseases [40]. AAV vectors have been used as delivery systems for gene therapy. Delivery of therapeutic AAV vectors resulted in clinical improvement in patients [41–44]. AAV9 vectors showed long-term effects in studies and clinical trials [45–48]. CIP is believed to have a higher affinity with Cdk5 than p25, which results in its inhibition to the activity of Cdk5/p25 [49]. In the present study, we delivered AAV9-mediated CIP into the brain of p25Tg mice following induction of p25, in which treatment of CIP inhibited the neurotoxicity of p25, successfully prevented neuronal death, inhibited tau hyperphosphorylation, and alleviated cognitive decline. Previously, we have successfully reversed the pathological and behavioral changes via ICV injection of AAV9-CIP in APP/PS1 mice, MPTP/p induced PD mice and p25 overexpression mice delivered by AAV8-GFP-p25 [18, 25]. The practical application of AAV9-CIP is an extension to the study of transgenic expression of CIP in p25Tg mice [17]. Collectively, AAV-mediated CIP is a potential strategy for the therapy of neurodegenerative diseases. Besides CIP, selective inhibit peptides, such as p3, p5, and p10, also have an inhibitory effect on neurotoxicity of p25 [23, 50]. Among these, TP5 or TFP5, shows neuroprotection in p25Tg mice, as well as AD and PD mouse models [22–24]. TFP5 can be delivered via intraperitoneal injection. However, repeated injections of TFP5 are inconvenient and may induce immunoreaction [41]. Administration of AAV vectors may resolve these problems and provide long-term neuroprotection. The feasibility of AAV vectors has also been demonstrated in a study with shCDK5miR (a microRNA-based short hairpin (sh-miR) against Cdk5). AAV delivered shCDK5miR in the hippocampus successfully reversed tau aggregation and alleviated spatial memory in 3×Tg-AD mice [39]. In brief, targeting neurotoxicity of p25 is effective to alleviate the pathological and behavioral changes in some neurodegenerative mouse models, and the application of AAV vectors as delivery system is practical.
Some limitations still exist in the present study. The activity of Cdk5 or other kinases, like GSK3β, had not been assessed due to the application of isotopic material restriction in our institute. Thus, we cannot conclude that CIP inhibition is directly target on Cdk5 hyperactivity, although this inhibition is indirectly suggested by the inhibition on several tau hyperphosphorylation. On the other hand, the effect of AAV9-CIP on WT mice would be further studied. Chemical inhibitors, such as roscovitine and olomoucine, have already profiled some preclinical toxicology and clinical trials. Studies showed that such inhibitors affected the eye and the pancreas islet, which may be used for helping detect the safety issues of selective inhibit peptides [30, 52]. Lastly, as we mentioned before, short term expression of p25 may enhance hippocampus-dependent memory [36]. We would like to know if transient p25 expression in our p25Tg mice or p25 overexpression mice delivered by AAV8-GFP-p25, we used before, has the same effect on spatial memory of mice and whether AAV9-CIP prevented the enhance of hippocampus-dependent memory related to short term expression of p25.
Conclusions
The present study provides further evidence that overexpression of p25 leads to tau hyperphosphorylation, weight loss, neuronal death, and behavioral changes in mice, which can be alleviated by administration of AAV9-CIP. These results provide evidence that the neurotoxicity of p25 can be alleviated by AAV9-mediated CIP.