
Abstract
Excitatory (E) and inhibitory (I) balance of neural network activity is essential for normal brain function and of particular importance to memory. Disturbance of E/I balance contributes to various neurological disorders. The appearance of neural hyperexcitability in Alzheimer’s disease (AD) is even suggested as one of predictors of accelerated cognitive decline. In this study, we found that GAD67+, Parvalbumin+, Calretinin+, and Neuropeptide Y+ interneurons were progressively lost in the brain of APP/PS1 mice. Transplanted embryonic medial ganglionic eminence derived interneuron progenitors (IPs) survived, migrated, and differentiated into GABAergic interneuron subtypes successfully at 2 months after transplantation. Transplantation of IPs hippocampally rescued impaired synaptic plasticity and cognitive deficits of APP/PS1 transgenic mice, concomitant with a suppression of neural hyperexcitability, whereas transplantation of IPs failed to attenuate amyloid-β accumulation, neuroinflammation, and synaptic loss of APP/PS1 transgenic mice. These observations indicate that transplantation of IPs improves learning and memory of APP/PS1 transgenic mice via suppressing neural hyperexcitability. This study highlights a causal contribution of GABAergic dysfunction to AD pathogenesis and the potentiality of IP transplantation in AD therapy.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative disease clinically manifested with cognitive deficits. AD is pathologically characterized with neurofibrillary tangles formed by hyperphosphorylated tau and amyloid plaques consisting of accumulated amyloid-β (Aβ) [1, 2]. The accumulated Aβ and hyperphosphorylated tau are toxic to neural cells, leading to impaired synaptic plasticity, neurodegeneration, and eventually cognitive deficits. Accumulation of oligomeric Aβ results in hyperphosphorylation of tau [3]. Thus, for a long time, prevention/reduction of Aβ accumulation is one of the prime strategies in AD therapy. However, the failures of the candidate drugs targeting Aβ in the late stage of clinical trials make us consider other strategies ameliorating cognitive deficits of AD.
The balance of excitatory (E)/inhibitory (I) neuronal network activity plays an essential role in maintaining cognitive function [4, 5]. E/I imbalance is associated with cognitive deficits in various neurological disorders [4–6]. Hyperexcitability of neural networks has been observed in AD patients and various AD model mice [4, 6]. Eliciting a long-lasting neuronal hyperexcitability accelerates Aβ pathology in APP transgenic mice [7]. Conversely, suppression of neural hyperexcitability in the brain network attenuates cognitive deficits of AD model mice [6, 9]. These lines of evidence point out neural hyperexcitability caused by E/I imbalance as one of causal factors in AD pathogenesis. Recent studies further highlight dysfunctional gamma-aminobutyric acid (GABA) signaling system, which plays an important role in maintaining the E/I balance of the nervous system [10, 11], and in regulating synaptic plasticity [12, 13], contributes to cognitive deficits of AD individuals [14–16]. Furthermore, manipulation of the GABAergic system can rescue the memory deficits of AD model mice [17, 18], emphasizing it as a potential therapeutic strategy in AD therapy.
Stem cell transplantation has shown promising achievements and good prospects in the treatment of various diseases including AD [19]. The GABAergic interneurons are differentiated from interneuron progenitors (IPs) enriched in the ganglionic eminences of the embryo consisted with medial ganglionic eminence (MGE), lateral ganglionic eminence (LGE), and caudal ganglionic eminence (CGE). Compared with CGE/LGE-derived IPs, MGE-derived IPs have better mobility and can successfully differentiate into GABAergic interneurons, thereby restoring the lost/impaired GABAergic signaling systems [20, 21]. Therefore, in this study, we transplanted MGE-derived IPs into the hippocampus of 8-month-old APP/PS1 transgenic mice, which overexpress mutant APP and PS1 and exhibit AD-like pathology at 7 months old [22]. Transplantation of MGE IPs rescues impaired synaptic plasticity and cognitive deficits of APP/PS1 transgenic mice, concomitant with a suppression of neural hyperexcitability, while showing no effect on Aβ load, neuroinflammation, and synaptic density. These studies highlight transplantation of IPs as a potential therapeutic strategy in AD therapy.
MATERIALS AND METHODS
Mice
APP/PS1 transgenic mice (stock number 004462) and EGFP transgenic mice (003291) were purchased from the Jackson Laboratory. APP/PS1 transgenic mice express mutant human APP and PS1 under prion promoter. EGFP transgenic mice with an “enhanced” GFP (EGFP) cDNA under the control of a chicken beta-actin promoter and cytomegalovirus enhancer. GAD67-GFP knock-in mice were described previously [23]. All mice were maintained in standard housing conditions on a 12-h light/dark cycle with food and water provided. Male APP/PS1 transgenic mice were used in all experiments. Animal care and surgical procedures were approved by the Institutional Animal Care and Use Committee of Soochow University in accordance with international laws. The number of animals used in each experiment was shown in Supplementary Table 1.
Antibodies
Mouse anti-GAD67 (1:1000; Abcam), goat anti-parvalbumin (1:1000; Abcam), rabbit anti-NPY (1:5000; Abcam), rabbit anti-calretinin (CR;1:500; Abcam), rabbit anti-LHX6 (1:200; Santa Cruz), goat anti-GFAP (1:1000; Abcam), goat anti-Iba1 (IBA1;1:500; Abcam), goat anti-PDGFα (1:300; R&D Systems), mouse anti-amyloid β (6E10;1:200; Convance), and rabbit anti- synaptophysin (SYN;1:500; Abcam). The fluorescence-conjugated secondary antibodies were purchased from Invitrogen.
Culturing of MGE progenitors
The MGE regions were dissociated from E13.5 EGFP transgenic mice as described [24]. The dissociated MGE were digested using Leibovitz L-15 medium containing 100 μg/ml DNAseI (Sigma) and centrifuged for 3 min at 1,000 g. Cell suspensions (∼105 cells/μl in L-15 medium containing 100 μg/ml DNase I) were kept on ice until used.
Cell transplantation
Injections were bilaterally made into dentate gyrus of the hippocampus at the following stereotaxic coordinates: anterior-posterior (AP) 2.0 mm, medial-lateral (ML) 1.5 mm, dorsal-ventral (DV) 2.0 mm and AP 3.2 mm, ML 3.0 mm, DV 3.0 mm. At each transplantation site, 2 μl cell suspensions (∼20,000 cells) were injected at a flow rate of 200 nl/min and allowed to diffuse for 5 min before it was slowly withdrawn. Sham-operated controls were injected with an equal volume of vehicle at each site. After the injection, mice received daily injections of cyclosporine A (10 mg/kg, intraperitoneally) for 4 weeks from the day before transplantation.
Immunostaining
Mice were transcardially perfused with 0.9% (w/v) saline solution then with 4% (w/v) paraformaldehyde (PFA) and the brains were removed and postfixed in 4% PFA for 24 h at 4°C. After washing with PBS, tissue was dehydrated in 30% (w/v) sucrose in PBS at 4°C. Coronal sections were cut with a frozen sliding microtome (Leica) for floating section immunohistochemistry. The sections were blocked for 1-2 h at room temperature (RT) with 5% bovine serum albumin (BSA) in PBS containing 0.3% Triton X-100 and then incubated with primary antibody diluted in PBS with 5% BSA for 24 h at RT at 4°C. The sections were incubated for 1-2 h at RT with a secondary antibody diluted in PBS. Images were captured under a laser confocal microscope.
Image analysis and quantification
Six brain slices with the same anatomical structure from each mouse and 3–6 mice from each group were selected for analysis. All images were analyzed and quantified by Image J (National Institutes of Health, Bethesda, MD, USA) as described previously [22, 25]. For quantification of the survived cells, all of the hippocampal sections were visualized and analyzed the total GFP+ cells. For differentiation of transplanted cells, the percentage of marker+GFP+/GFP+ was analyzed. The numbers of Aβ plaques were defined as the density of Aβ plaques per μm2. The size of Aβ plaques was quantified as the area that was occupied by Aβ plaques divided by total area of hippocampus. The relative numbers of microglia and astrocytes was quantified as the area that was occupied by GFAP+ or Iba1+ signals divided by total area of the hippocampus. SYN-immunoreactivity was assessed by mean fluorescence intensities (MFIs) using Image J.
Behavioral tests
Behavioral tests were performed at 60–70 day after transplantation. Novel object recognition (NOR) and Morris water maze (MWM) were conducted to evaluate the ability of animal cognition. 50 cm×50 cm×50 cm box was used for NOR test. After three days of habituation, the mice were exposed to 2 identical objects placed in two opposite corners of the apparatus 5 cm from the sidewall for 10 min. Ninety minutes after the training time, the mice were returned to the open field in the presence of one familiar and one novel object for 10 min. Preference index (PI) = time exploring one of the identical objects/time exploring the identical object pairs ×100%. Recognition index (RI) = time exploring novel object / (time exploring novel object + time exploring familiar object) ×100%.
The MWM test was conducted in a pool (120 cm in diameter) with water temperature at 22–23°C and with a 10 cm platform submerged 1 cm below the surface of opaque water during hidden trials. Mice were trained 4 trials per day with a different start location. Each trial lasted 90 s until the mice found the platform. If not, the mouse would be gently guided to the platform and allowed to stay on the platform for 10 s. Escape latencies (time spent swimming from start point to the target) and path length (the distance from start point to the platform) were recorded for 5 consecutive days. For probe trials, the platform was removed after the last trial of the acquisition period. The average speed and the numbers of platform-site crossing were recorded during 60 s on the sixth day.
Electrophysiological recordings for LTP
Transverse hippocampal slices (300 μm thick) were prepared from 10-month-old mice by a vibratome (LEICA VT1200 S). Slices were cut in cold artificial cerebrospinal fluid (ACSF, 4°C) bubbled with 95% O2 and 5% CO2 containing: 110 mM NaCl, 2.5 mM KCl, 1.5 mM, MgSO4·2H2O, 2.5 mM CaCl2, 1.25 mM NaH2PO4, 26 mM NaHCO3, and 10 mM D-glucose (pH 7.3). Field excitatory postsynaptic potentials (fEPSPs) were recorded from the stratum radiatum of CA1 using glass microelectrodes filled with 3 M NaCl solution (resistance 1–2 MΩ). Test stimuli were delivered at 0.033 Hz (0.2 ms duration) through concentric bipolar electrodes, placed in the CA1 area of the hippocampal slice to stimulate Schaffer collateral pathway (SC-CA1). After baseline was recorded for 10 min at an intensity that was set to 40% of the maximal response, LTP was induced using high frequency stimulation (four 100 Hz and 1 s trains delivered 20 s apart). The data were acquired with an Axon multiclamp 700 B amplifier, filtered at 0.1e5 KHz, and digitized at 10 KHz, and analyzed offline by pClamp10.3 software (Molecular Devices Corp, USA). Input-output (I/O) curves were established by single-pulse stimulation of the SCs region in order to evaluate synaptic efficacy by adjusting the stimulus intensity between by steps 0.05–1.0 mA. Stimulus pulses were delivered at 0.033 Hz and five responses at each current intensity were averaged. Presynaptic function was explored by using paired-pulse facilitation (PPF) paradigm with inters-stimulus intervals (ISIs) ranging from 25, 50, 75, 100, 125, 150, and 200 ms. Facilitation was measured as a ratio of the second pulse-evoked EPSP slope to the first evoked, averaged over five responses per pulse pair.
Electroencephalographic recordings
Mice were anesthetized and implanted with Teflon-coated silver wire electrodes (0.125 mm diameter) after finishing behavior tests. Chronic electroencephalographic (EEG) was monitored. The position of electrodes implanted were as follows (bregma as reference): left frontal cortex: anteroposterior (AP), ±1 mm; mediolateral (ML), ±1 mm); the left and right parietal cortices (AP, ±2 mm; ML, ±2 mm). For depth EEG recordings, electrodes were implanted bilaterally into the hippocampus followed by the coordinates: AP, ±2.46 mm; ML, ±1.66 mm; and dorsoventrally, –1.87 mm. Electrode implanted into the subdural space of the left frontal cortex was defined as reference. Electrodes were cemented by a fast-acting adhesive and dental acrylic. After 7 days of recovery period at least, the EEGs were recorded for up to 3 days and 3 h each day under condition that mice can freely move in the test cage. The data were acquired daily with the Powerlab 8/35 software (version 7, Labchart). Epileptic spikes were detected automatically by the Gotman spike and seizure detectors from Powerlab. All behavior was monitored by a video camera during EEG recording periods.
Statistical analysis
All statistical analysis was conducted using SPSS 20.0. Comparisons between two groups were made by independent samples t-test. Data between multiple groups were analyzed by ANOVA followed by Fischer protected least significant difference post hoc tests. Data are presented as the mean±SEM. p < 0.05 was considered statistically significant (*p < 0.05, **p < 0.01, ***p < 0.001).
RESULTS
APP/PS1 mice exhibit loss of subtypes of interneurons
We first examined the numbers of GABAergic interneurons in APP/PS1 model mice, which overexpress mutant human APP and PS1 and are model mice commonly used in AD research. GAD67-GFP-knock in mice, which specifically express GFP in GABAergic neurons under the control of endogenous GAD67 promoter [23], were cross-bred with APP/PS1 mice to visualize interneurons in APP/PS1 mouse brains (Fig. 1A). APP/PS1 mice exhibited comparable numbers of GFP+ cells at 2 months old (Supplementary Figure 3), but reduced numbers of GFP+ cells at 7 months of age compared to wild-type mice (Fig. 1B). We examined the subtypes of GABAergic interneuron, such as calretinin (CR)+, PV+ and NPY+ interneurons in both hippocampus and cortex of APP/PS1 mice at 2, 7, and 12 months of age (Fig. 1C and Supplementary Figure 1). Consistent with the observations using GAD67-GFP-knock in mice, CR+, PV+, and NPY+ interneurons in the hippocampal DG of 2-month-old APP/PS1 transgenic mice showed comparable numbers with those in WT mice (Fig. 1C, D and Supplementary Figure 1). However, both CR+ and NPY+ interneurons exhibited reduced numbers in the hippocampal DG of 7- and 12-month-old APP/PS1 transgenic mice (Fig. 1C, D), when these mice already exhibit AD-like pathology and cognitive deficits [22]. Decreased numbers of NPY+ interneurons were also observed in the other hippocampal subregions such as CA1 and CA3 of 7- and 12-month-old APP/PS1 transgenic mice (Supplementary Figure 1A, B, D). In contrast, in the CA1 and CA3 subregions, no differences in the numbers of CR+ interneurons have been observed between APP/PS1 transgenic mice and WT littermates at all age indicated in this study (Supplementary Figure 1A, B, E). Decreased numbers of PV+ interneurons were only observed in the hippocampus of APP/PS1 at 12 months of age, rather than younger age (Fig. 1C, D and Supplementary Figure 1A–C). These results indicate that APP/PS1 transgenic mice exhibit loss of GABAergic interneurons in an age-dependent manner.
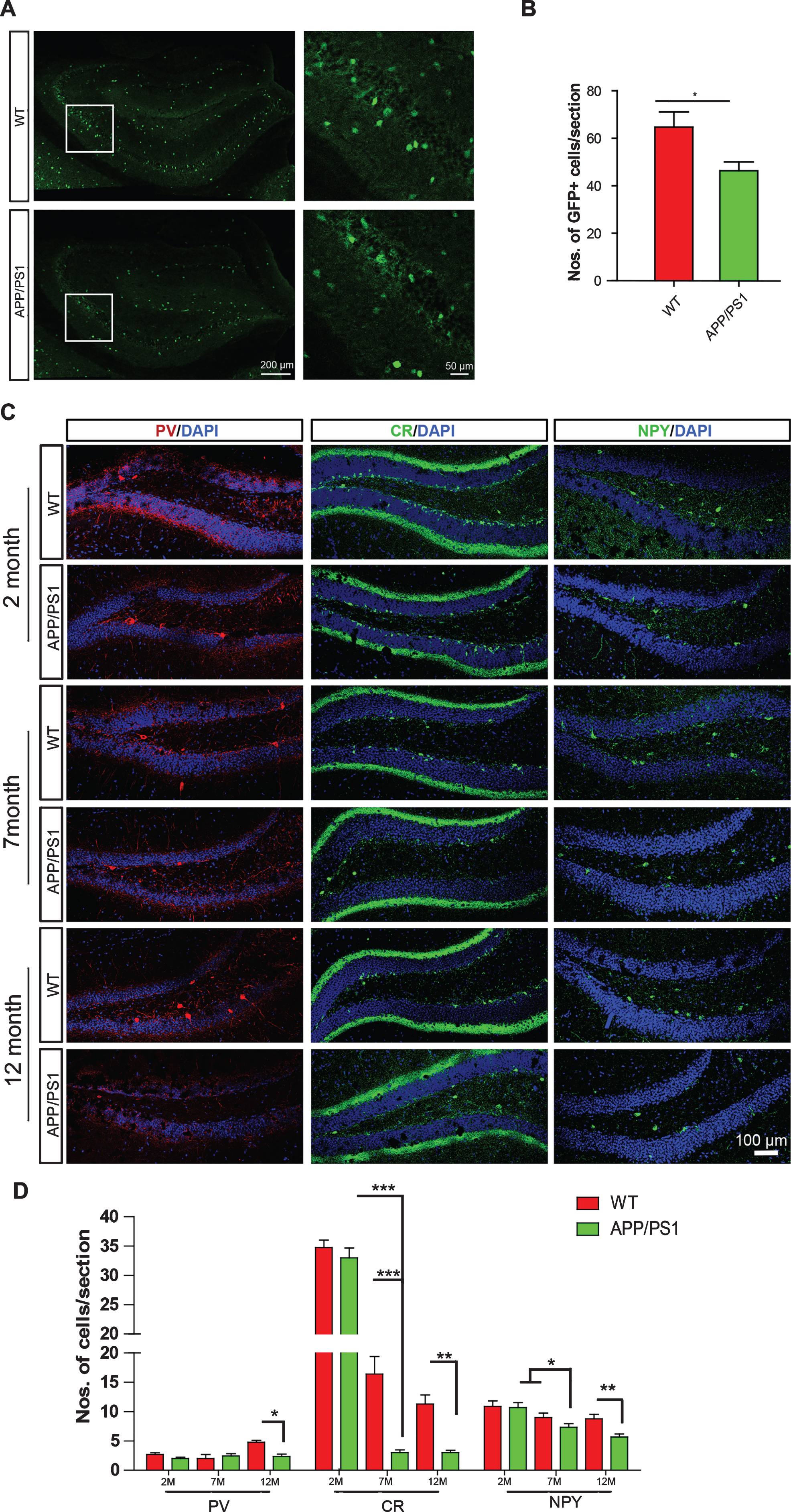
APP/PS1 transgenic mice exhibit reduced numbers of GABAergic interneurons in the hippocampus at 7 months of age. A) GAD67-GFP+ cells in the hippocampus of 7-month-old GAD67-GFP:APP/PS1 double transgenic and GAD67-GFP mice. B) Numbers of GAD67-GFP+ interneurons per hippocampal slice. C) The coronal sections of the DG regions of APP/PS1 transgenic mice at 2, 7, and 12 months of age were immunostained for PV, CR, NPY, and DAPI counterstaining. D) Numbers of marker+ cells per section. Scale bars: 200 μm (left panel in A), 50 μm (right panel in A), 100 μm. Data represent mean±SEM; n = 3 (B), 4–6 (D). *p < 0.05; **p < 0.01. ***p < 0.001. Student’s t-test (B). One-way ANOVA (D).
Transplanted IPs survive and differentiate to GABAergic interneurons in the hippocampus of APP/PS1 transgenic mice
Stem cell transplantation show promising potential in therapy of neurological disorders. Considering the fact that loss of GABAergic interneurons in the brains of APP/PS1 transgenic mice, we were wondering whether transplantation of IPs could rescue impaired learning and memory of APP/PS1 transgenic mice. IPs were isolated from the MGE of embryonic 13.5 EGFP transgenic mice and validated as described [24]. IPs were transplanted into the hippocampal DG of 7-month-old APP/PS1 transgenic mice, which already harbor extensive Aβ load, neuroinflammation, synaptic loss, and exhibiting cognitive deficits [9, 22]. Transplanted GFP+ IPs were further validated by immunostaining for Lhx6, a marker of interneuron progenitor [24]. To analyze the survival and migration of transplanted IP, we analyzed the distribution of EGFP+ IPs in the hippocampus of host mice at 7 days, 30 days, and 60 days upon transplantation, respectively. At 7 days upon transplantation, the transplanted EGFP+ cells were mainly distributed along the injection sites and did not migrate to a large extent, and only a small portion of the transplanted cells began to migrate. At the 30th day of transplantation, the transplanted EGFP+ IPs has migrated to the hippocampal DG subregion, and a small portion of cells have migrated outside the DG subregion. At the 60th day of transplantation, the transplanted EGFP+ IPs had a good migration in the hippocampus, and some EGFP+ IPs show branches (Supplementary Figure 2). The number of survived GFP+ IPs per unilateral hippocampus were 6336.667±477.2607 (mean + SEM) 7 days after transplantation, which decreased to 2867.333±401.571 (mean + SEM) 60 days after transplantation (Fig. 2D).
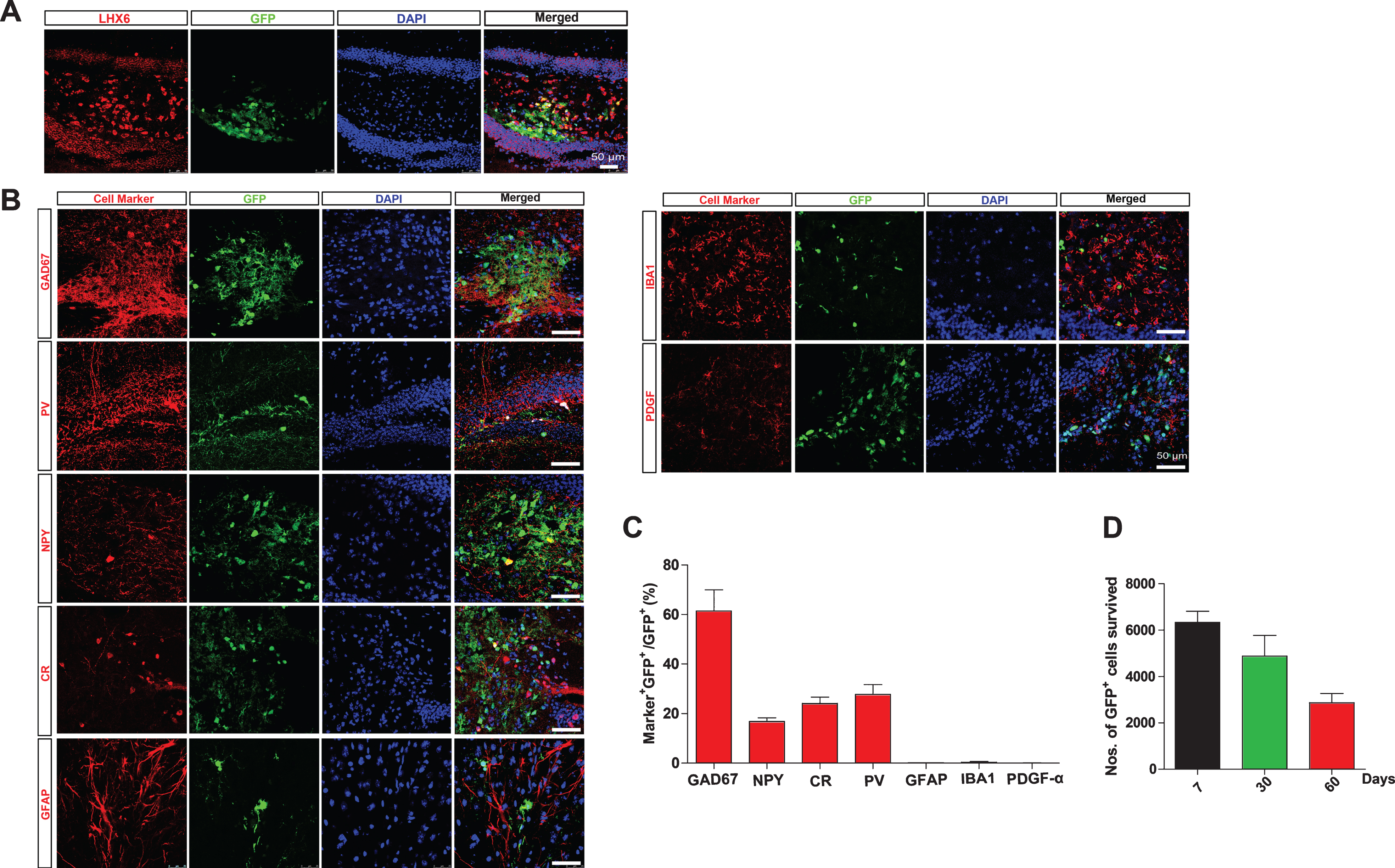
MGE-derived IPs differentiate into GABAergic interneurons. A,B) The coronal hippocampal sections of APP/PS1 transgenic mice transplanted with IPs for two months were immunostained for Lhx6 (A), GAD67, PV, NPY, CR, GFAP, Iba1, and PDGFRα (B), and DAPI counterstaining. D) Numbers of survival transplanted cells per unilateral hippocampus. C) The percentage of marker+GFP+/GFP+ (%) per section. Statistical analysis of differentiation into various types of cells after IP transplantation. Data represent mean±SEM; n = 3–6. Scale bars: 50 μm.
We further examined whether the transplanted GFP+ IPs could differentiate into GABAergic interneurons and their subtypes by immunostaining for GAD67, NPY, CR, and NPY in the hippocampal sections of the host mice transplanted with IPs for 2 months (Fig. 2B). The results revealed that nearly 61.450±8.486% transplanted GFP+ IPs differentiated into GAD67+ GABAergic interneurons. Among them, 16.850±1.438%, 24.117±2.541%, and 27.783±3.948% (mean + SEM) transplanted GFP+ IPs differentiated into NPY+, CR+, and PV+ interneurons respectively (Fig. 2C). In contrast, few transplanted GFP+ IPs differentiate into GFAP+ astrocytes, Iba1+ microglia or PDGFα+ oligodendrocytes, further confirming that transplanted EGFP+ IPs specifically differentiate into interneurons, rather than glial cells (Fig. 2B, C). These data indicate that transplanted IPs differentiate successfully into GABAergic interneurons, even under an “AD”-like hostile environment.
Transplantation of IPs shows no effect on Aβ-related pathology of APP/PS1 mice
Accumulation of Aβ is a key initiator of AD pathogenesis, which results in neuroinflammation, loss of synapses, and suppressed synaptic plasticity [3]. We thus first examined whether transplantation of IPs ameliorate accumulation of Aβ. APP/PS1 transgenic mice were transplanted GFP+ IPs at 7 months of age and subjected to analysis of Aβ plaque density 2 months after transplantation (Fig. 3A), when most of transplanted IPs differentiated into GABAergic interneurons (Fig. 2). The age- and gender-matched APP/PS1 transgenic mice and wildtype (WT) mice, which were stereotaxically injected with culture medium, were used as control mice. We immunostained hippocampal slices of each group of mice with anti-Aβ antibody (6E10) (Fig. 3B) and analyzed the densities of Aβ plaques. The results showed that neither the numbers nor the size of Aβ plaques in the IPs-transplanted APP/PS1 transgenic mice differed from those in control APP/PS1 transgenic mice (Fig. 3C, D).
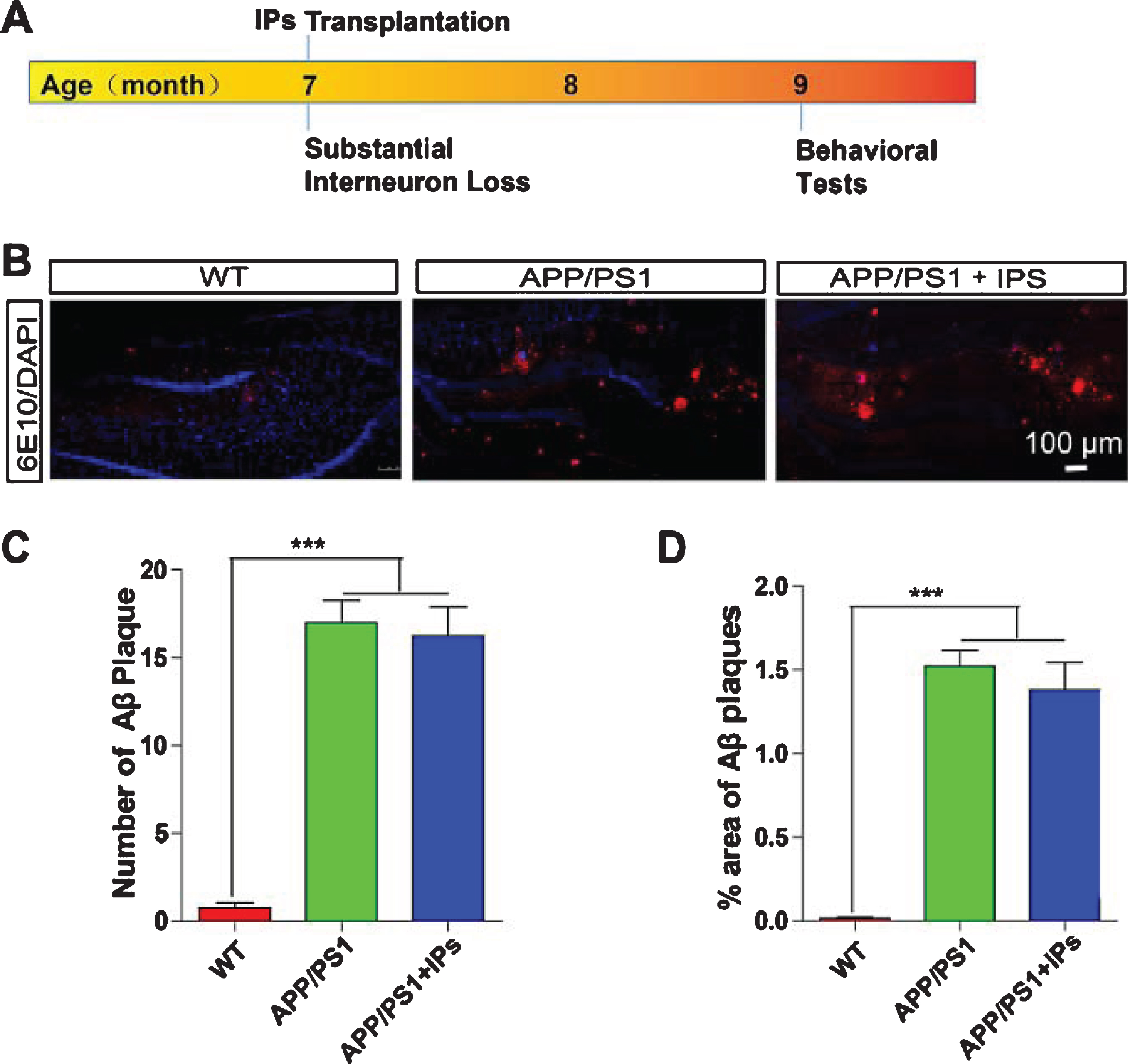
IPs transplantation exhibits no effect on Aβ load in APP/PS1 mice. A) Timeline for IPs transplantation and behavioral tests. B) The coronal hippocampal sections of APP/PS1 transgenic mice transplanted with IPs were immunostained for Aβ (6E10) and DAPI counterstaining. C) Numbers of Aβ plaques per hippocampal section. D) The percentage of the area occupied by Aβ plaques. n = 5 mice per group.
Neuroinflammation is an essential contributor to AD pathophysiology [26]. To investigate whether transplanted IPs ameliorates neuroinflammation, we analyzed the densities of astrocytes and microglia, two types of cells eliciting the inflammatory response in the brains of AD patients and AD model mice [26], by staining the hippocampal sections for GFAP and Iba1, which are markers for astrocytes and microglia respectively. Neither astrocytes (Fig. 4A, D) nor microglia (Fig. 4B, E) exhibited different densities in the hippocampus of the IPs-transplanted APP/PS1 transgenic mice comparing to those in control transgenic mice, indicating that transplantation of IPs failed to ameliorate neuroinflammation of APP/PS1 transgenic mice.
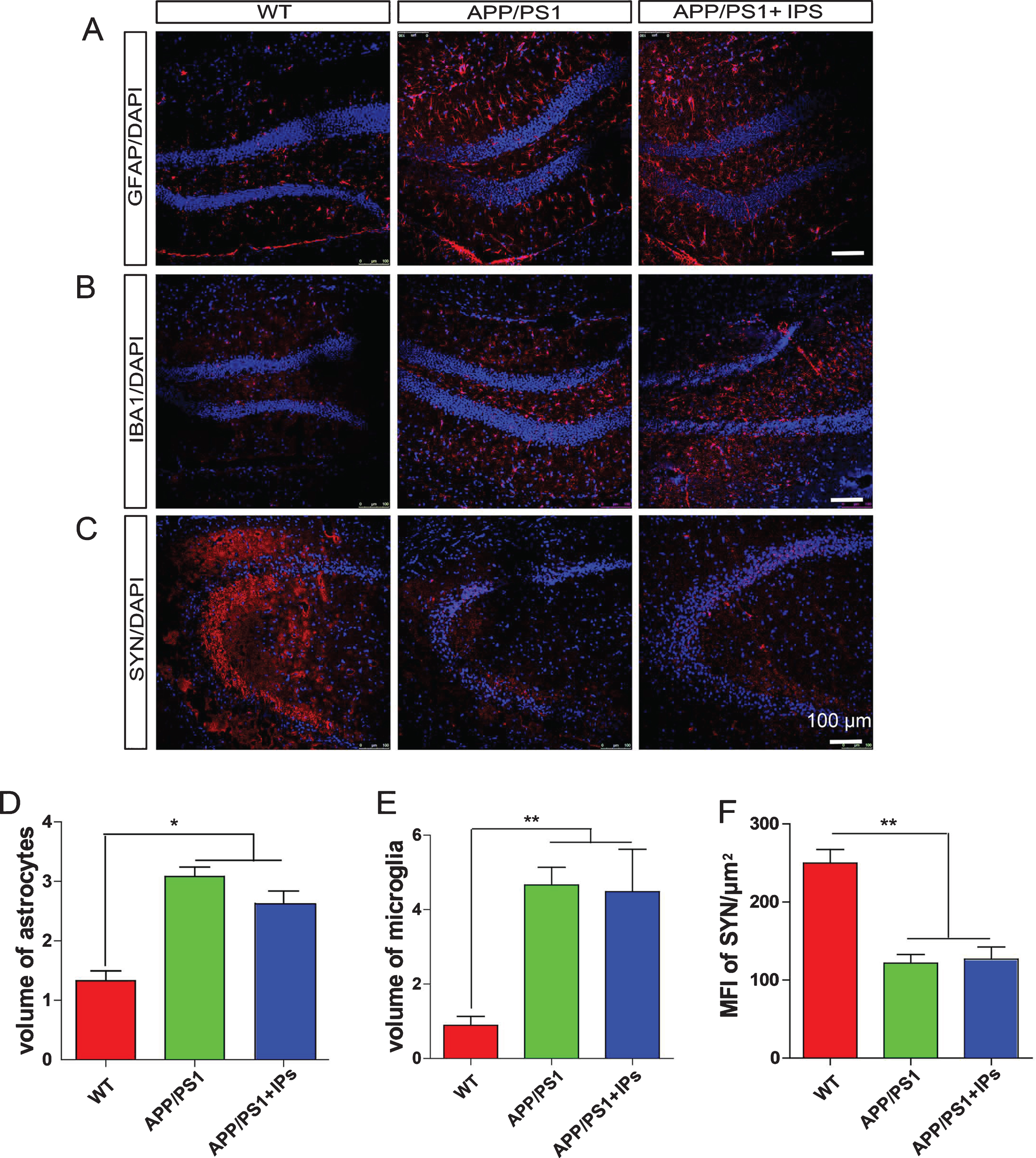
IPs transplantation exhibits no effect on neuroinflammation and synapse loss in APP/PS1 mice. A–C) The coronal hippocampal sections of APP/PS1 transgenic mice transplanted with IPs were immunostained for GFAP (A), IBA1 (B), synaptophysin (SYN, C), and DAPI counterstaining. D, E) The percentage of the area occupied by astrocytes (D) or microglia (E) in total hippocampal area. F) The mean fluorescent intensity (MFI) of SYN immunoreactivity in the hippocampus. Data represent mean±SEM. n = 5 mice per group. *p < 0.05, **p < 0.01, ***p < 0.001. One-way ANOVA. Scale bar: 100 μm.
Loss of synapses, which is strongly correlated with cognitive deficits, has been observed the brains of AD patients and model mice [27–30]. To investigate whether transplantation of IPs attenuates loss of synapses in APP/PS1 transgenic mice, we immunostained the hippocampal sections for synaptophysin (SYN), a protein specifically expressed in presynapses and representing synaptic function [31]. IPs-transplanted APP/PS1 mice contained identical levels of SYN+ immunoreactivity in the hippocampus as control APP/PS1 mice, both of which harbored less SYN+ immunoreactivity than WT mice (Fig. 4C, F). These results indicate that transplantation of IPs fails to attenuate synaptic loss in APP/PS1 transgenic mice.
Transplantation of IPs attenuates neural hyperexcitability in the hippocampus of APP/PS1 mice
Neural hyperexcitability characterized by frequent abnormal spikes and increasing numbers of epileptic activities is one of causal factors contributing to cognitive deficits of AD [15, 32]. Suppression of hyperexcitability in brain improves cognition in various AD model mice [6]. We further examined whether IP transplantation would suppress hyperexcitability in the brains of APP/PS1 transgenic mice by EEG monitoring. As described, APP/PS1 transgenic mice exhibited neural hyperexcitability in the hippocampus and cortex, which is characterized by increased appearance frequency of epileptic spikes. IPs-transplanted APP/PS1 transgenic mice exhibited decreased appearance frequency of epileptic spikes compared to control APP/PS1 transgenic mice in the hippocampus (Fig. 5A, B), but not in the cortex where no IPs being transplanted (Fig. 5A, C). Therefore, these results indicate transplantation of IPs attenuates neural hyperexcitability in APP/PS1 mouse brains.
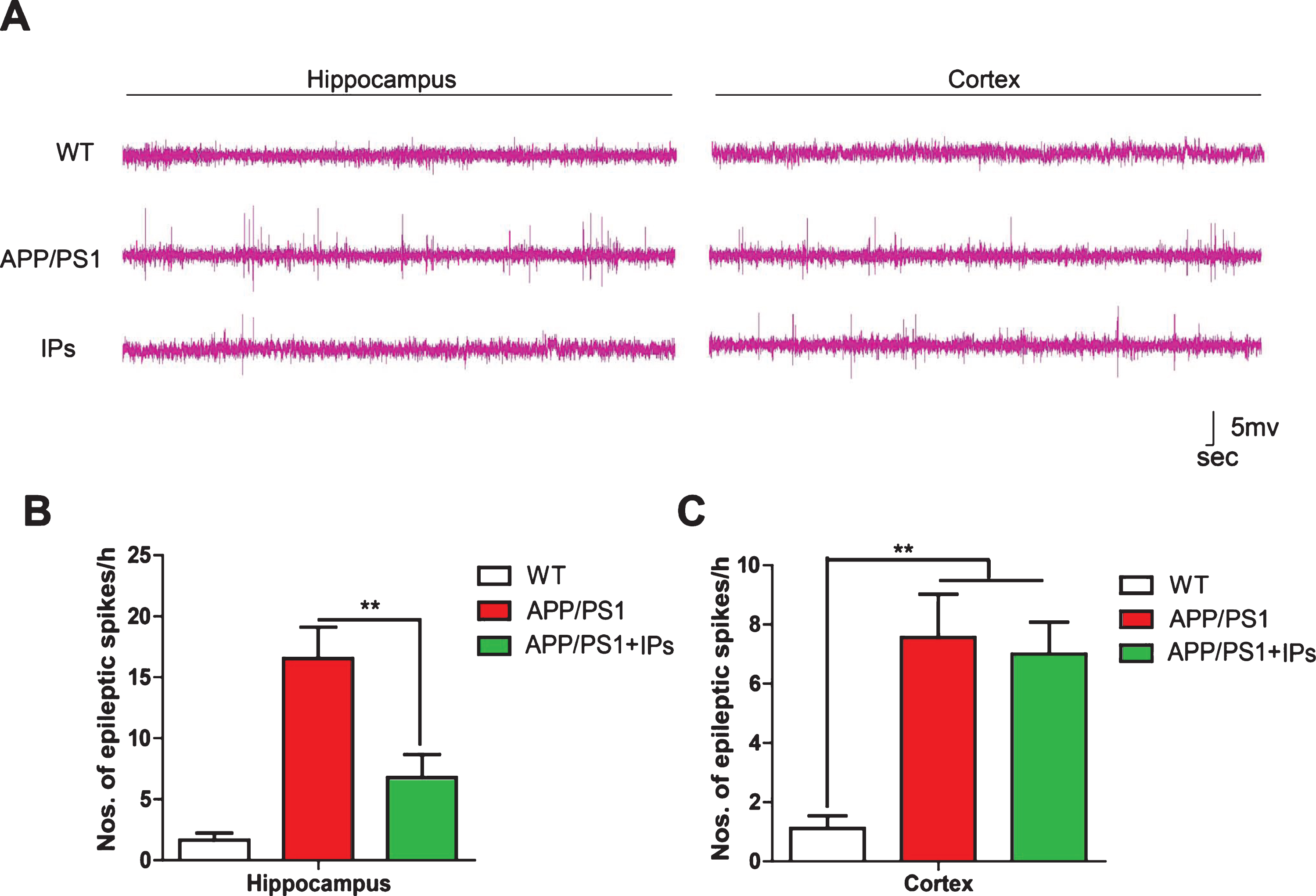
IPs transplantation attenuates hyperexcitability in the hippocampus of APP/PS1 mice. A) Representative images of brain EEG in hippocampus and cortex. B,C) the frequency of occurrence of epileptic spikes in the hippocampus (B) and cortex (C) of each group of mice. Data represent mean±SEM. n = 4–6 mice per group. **p < 0.01. One-way ANOVA.
Transplantation of IPs rescues suppressed long-term potentiation in APP/PS1 transgenic mice
To further investigate whether transplanted IPs-suppressed hyperexcitability benefits synaptic plasticity, which is impaired in the brains of AD patients and AD model mice [9, 33], we examined long-term potentiation (LTP) in the hippocampus of three groups of mice. As described, LTP at SC-CA1 synapse was impaired in control APP/PS1 transgenic mice. In contrast, IPs-transplanted APP/PS1 mice showed an enhanced LTP, even on a level comparable with WT mice (Fig. 6), indicating that transplantation of IPs rescues suppressed LTP in APP/PS1 transgenic mice.

IPs transplantation rescues suppressed LTP in APP/PS1 mice. A) Time course of the effects of High-Frequency-Stimulation (HFS) on the fEPSP initial slope. B) Normalized fEPSP slops. C) The I/O curve of each group. D) The PPF ratio of each group. Data represent mean±SEM. n = 5–7 mice per group. *p < 0.05.
To examine whether the properties of basic synaptic transmission at SC-CA1 synapses is altered in IPs-transplanted APP/PS1 mice, input-output curves were obtained by measuring the post-synaptic potential slope with varying stimulus intensities (0.05–1.0 mA). Data were normalized using 100% as the highest amplitude (average of five selected sweeps in each stimulation intensity) of the fEPSP. IPs-transplanted APP/PS1 mice showed no significant change in the input-output curve compared to APP/PS1 transgenic mice (Fig. 6C). This indicates that transplantation of IPs does not affect the basal synaptic response. PPF was obtained after the I/O curve measurements to determine the probability of synaptic vesicle release, and this was measured by paired pulses at intervals between 25 and 200 ms. As shown in Fig. 6D, there is no differences between IPs-transplanted APP/PS1 mice and Control APP/PS1 transgenic mice at any inter-stimulus interval. This finding suggests that transplantation of IPs may not influence the probability of synaptic vesicle release from the pre-synaptic terminals. Thus, the improvement of synaptic plasticity by transplanted IPs may be related to the reduction of Aβ toxicity for post-synaptic neuron.
Transplantation of IPs rescues cognitive deficits of APP/PS1 transgenic mice
We further analyzed whether IP transplantation improved cognitive impairment of APP/PS1 mice by performing behavioral tests. APP/PS1 transgenic mice were transplanted GFP+ IPs at 7 months of age and subjected to behavioral tests 2 months after transplantation (Fig. 3A), when most of transplanted IPs differentiated into GABAergic interneurons. We first performed new object recognition (NOR) experiment to analyze whether transplantation IPs improve short-term memory in APP/PS1 mice as described previously [9, 22]. As shown in Fig. 7A, three groups of mice did not show a preference for an object when exploring the same object. Control APP/PS1 transgenic mice failed to show a preference for new objects as WT did, indicating cognitive impairment in these mice. However, APP/PS1 transgenic mice transplanted with IPs showed a preference for new objects with a recognition index of 0.6, which is comparable with that in WT mice (Fig. 7B).

IPs transplantation rescues cognitive deficits of APP/PS1 mice. A,B) Novel object recognition. Preference index (A) and recognition index (B). D–H) Morris water maze. Escape latency (C). Representative images of the path that mice swam along to find the platform (D). The distance that the mice swam to find the platform (E). The times that mice swam across the target sites after retrieval of the platform (F). Swimming velocity (G). Data represent mean±SEM. n = 9–13 mice per group. *p < 0.05, * *p < 0.01. Two-way ANOVA.
Next, we conducted a water maze experiment to evaluate whether IP transplantation improved the spatial learning and memory of APP/PS1 mice. Compared with WT mice, the control APP/PS1 mice exhibited longer escape latency (Fig. 7C) and swimming distance (Fig. 7D, E) to locate the platform, indicating they have learning and memory impairment. In contrast, IPs-transplanted APP/PS1 transgenic mice showed decreased escape latency and swimming distance compared to the control APP/PS1 mice, even to a level comparable with WT mice (Fig. 7D, E). In the probe trial, APP/PS1 transgenic mice transplanted with IPs swam more times across the site where the platform originally located compared to control APP/PS1 mice (Fig. 7F). No differences in the swimming speed were observed among three groups of mice (Fig. 7G), indicating that the differences observed among three groups of mice were not due to swimming capability. In summary, there results indicate that transplantation of IPs rescues the cognitive deficits of APP/PS1 transgenic mice.
DISCUSSION
Network hyperexcitability has been observed at the early stage of AD as indicated by an increased incidence of seizures [34, 35] and detection by means of fMRI in individuals at risk of developing dementia [36], even in patients with mild cognitive impairment, which is considered to be a prodromal stage of AD [37]. Network hyperexcitability has also been detected in numerous mouse model of AD pathology including APP/PS1 transgenic mice used in the present study [9, 39]. The appearance of network hyperexcitability in AD is even suggested as predictors of accelerated cognitive decline [34, 35]. The aberrant network hyperexcitability contributes to AD pathology via accelerating neurodegeneration, enhancing Aβ and tau release, increasing regional vulnerability to Aβ and enhancing Aβ- and tau-induced pathology [25, 41]. Suppressing hyperexcitability has been shown to ameliorate cognitive deficits in both AD patients and animal models [6, 43]. Therefore, these lines of evidence highlight a therapeutic potential of suppression of neural hyperexcitability in AD therapy.
GABAergic transmission is a key regulator of hippocampus-dependent learning, as it associates with encoding and retrieval of memories [42–44]. Emerging literatures indicate that dysfunctional GABAergic transmission plays essential roles in AD pathogenesis. Hypofunction of Nav1.1, a voltage-gated sodium channel predominantly expressed in PV+ interneurons, has been observed in various AD model mice including hAPP-J20, Tg2576, and the transgenic mice overexpressing BACE1 [15, 45]. Dysfunction of SST+ interneurons, which is caused by decreased cholinergic innervation, also contributes to the cognitive deficits of APP/PS1 transgenic mice [16]. Moreover, accumulation of Aβ impairs interneuron function [15, 46]. Furthermore, restoring GABAergic interneuron function rescues cognitive deficits of AD model mice, highlight a causal contribution of GABAergic dysfunction to AD pathogenesis. We herein have observed that loss of GABAergic interneurons in the aged brains of APP/PS1 transgenic mice. This observation is consistent with previous reports that decreased GABA+ and SST+ immunoreactivity have been observed in the cerebral cortex of AD patients [47, 48] and in apoE4 knock-in mice [18].
Transplantation of interneuron progenitors exhibit great potentiality in the therapy of various neurological disorders such as epilepsy, schizophrenia, Parkinson’s disease, and autism spectrum disorders [49]. Transplanted IPs have been shown to increase both synaptic and extrasynaptic inhibition onto host pyramidal neurons [50] and granule cells [24, 51], activation of extrasynaptic GABA receptors [52], thus restoring the hyperexcitability of brain network [24]. Furthermore, transplanted IPs have been shown a capability of driving the endogenous neural plasticity and reshape the neural network [53, 54]. We herein have observed that the transplanted IPs can survive and differentiate into subtypes of GABAergic interneurons, even in a hostile environment enriched with soluble and insoluble Aβ, which impairs interneuron function [15, 46]. Transplanted IPs rescue impaired synaptic plasticity and cognitive deficits in APP/PS1 transgenic mice, concomitant with a suppression of neural hyperactivity. It is well known that increased neural activity increases Aβ releasing and regional vulnerability to Aβ [7, 41]. Consistently, suppression of hyperexcitability attenuates accumulation of Aβ [42]. However, we failed to observe any effect of transplantation of IPs on Aβ accumulation and Aβ-related pathology, despite the fact that transplantation of IPs successfully rescued hyperexcitability. We suspect the controversy observation may due to the fact that the transplantation was performed on 7-month-old APP/PS1 model mice, when these mice already harbored severe Aβ accumulation. Our observation is also consistent with the report that neural activity seems to affect the formation of Aβ plaques rather than the clearance of Aβ [42]. Prophylactic transplantation of IPs at the earlier stage may be more helpful in reducing Aβ accumulation. However, it is worth noting that merely inhibiting neural hyperexcitability is sufficient to restore cognitive impairment in APP/PS1 mice as shown in this study. This is consistent with the evidence that suppression of aberrant network hyperactivity using anti-epileptic drugs such as levetiracetam and lamotrigine, both of which are non-selective sodium channel blockers, attenuates the cognitive deficits of both AD patients and AD model mice [6, 9] and transplantation of IPs rescues cognitive deficits in apoE knock-in mice [18]. Therefore, these evidence indicate that transplantation of IPs attenuates cognitive dysfunction, at least partially through suppression of hyperexcitability.
In contrast to IP transplantation, transplantation of other types of stem cells also exhibit a therapeutic potential in AD pathology. For example, transplantation of neural stem cells decreases the accumulation of Aβ [55–57] and neuroinflammation [58, 59], which are not seen in AD transgenic mice transplanted with IP as shown in this study. Thus, our results suggest that different types of neural progenitors may display distinct behaviors upon transplantation. In addition, one of mechanisms underlying stem cell therapeutic strategy in AD is upregulating the levels of growth factors such as brain-derived neurotrophic factor (BDNF), insulin growth factor (IGF-1), nerve growth factor (NGF), and vascular endothelial growth factor (VEGF) [60]. Although we have not tested here, the growth factors secreted by the transplanted cells are another factor that cannot be ignored, and they may also be beneficial to the improvement of AD cognition by IP transplantation [61, 62].
The transplanted IPs differentiate into several subtypes of GABAergic interneurons, which regulate neural network via distinct fire patterns. For example, SST+ and NPY+ cells tend to fire tonically and independently of brain and behavioral states. Whereas PV+ cells fire predominantly during brain states that promote encoding [63]. The mechanisms by which interneuron subtypes deficits contribute to cognitive dysfunction in AD remain to be fully elucidated. Dysfunction of PV+ cells may contribute to the deficits in theta-gamma coupling in the brains of AD mice, which are associated cognitive deficits [64, 65]. Loss of SST+ cells is linked to cognitive deficits in APOE-KI mice via decreasing GABAergic inhibition [17]. However, another study indicates that decreasing cholinergic action on O-LM interneurons, one type of SST+ cells in the hippocampus, contributes to cognitive impairment in APP/PS1 mice [16]. Thus, these subtypes of interneurons may also attenuate the cognitive deficits of APP/PS1 transgenic mice via other different mechanisms, in addition to via suppressing neural hyperexcitability. It remains further investigation how these distinct subtypes of GABAergic interneurons differentiated from transplanted IPs contribute to the cognitive improvement in AD transgenic mice.
It is worth noting that only limited numbers of the survived transplanted IPs were survived, i.e., 2867.333±401.571 cells per unilateral hippocampus, at 2 months after transplantation. The number of transplanted IPs is enough to suppress neural hyperexcitability in APP/PS1 transgenic mice. Our observation is consistent with the reports that loss of about 2500–3000 GABAergic interneurons [66] would cause abnormal behaviors and one GABAergic interneuron regulates more than 1500 excitatory granule neuron in the dentate gyrus [67]. Thus, although transplanted IPs failed to ameliorate Aβ accumulation and Aβ-related pathology such as neuroinflammation and loss of synapses, transplanted IPs still rescued impaired synaptic plasticity and cognitive deficits of APP/PS1 transgenic mice. The present study further highlights a potential therapeutic effect of IP transplantation. In this context, due to the ethical limitation of human embryonic IPs, induced pluripotent stem cells, which could differentiate into IPs [68], is another option in AD therapy.
Footnotes
ACKNOWLEDGMENTS
This work was supported by National Natural Science Foundation of China (81870897, 81901296, 81601111, 81671111, 81571061), Natural Science Foundation of Jiangsu Province (BK20181436), Guangdong Key Project in “Development of New Tools for Diagnosis and Treatment of Autism” (2018B030335001), National Major Scientific and Technological Special Project for “Significant New Drugs Development” (2019zx09301102), the Suzhou Clinical Research Center of Neurological Disease (Szzx201503), translational medicine fund of WuXi municipal health commission (ZM010), Priority Academic Program Development of Jiangsu Higher Education Institution (PAPD), Jiangsu Provincial Medical Key Discipline Project (ZDXKB2016022) and Jiangsu Provincial Special Program of Medical Science (BL2014042), Jiangsu Key Laboratory of Translational Research and Therapy for Neuro-Psycho-Diseases (BM2013003).