
Abstract
We have previously reported that low doses of external beam ionizing irradiation reduced amyloid-β (Aβ) plaques and improved cognition in APP/PS1 mice. In this study we investigated the effects of radiation in an age-matched series of 3xTg-AD mice. Mice were hemibrain-irradiated with 5 fractions of 2 Gy and sacrificed 8 weeks after the end of treatment. Aβ and tau were assessed using immunohistochemistry and quantified using image analysis with Definiens Tissue Studio. We observed a significant reduction in Aβ plaque burden and tau staining; these two parameters were significantly correlated. This preliminary data is further support that low doses of radiation may be beneficial in Alzheimer’s disease.
INTRODUCTION
Existing pharmaceuticals provide only short-term symptomatic relief of Alzheimer’s disease (AD) without affecting disease progression [1] and new drug development is time consuming and costly. To address the paucity of effective AD treatments, we reported the use of modest doses of ionizing radiation therapy as a novel treatment against amyloid pathologies associated with AD [2, 3]. This research was based on the observation that radiotherapy (RT) has been successfully used to treat extracranial amyloidosis [4–6]. Applying RT to the AD brain has not been considered due to the known side-effects on the brain but these effects are completely dose dependent where total doses of 30–60 Gy given in 2 Gy fractions can produce macroscopic tissue destruction [7] and impair cognition [8–12]. Prophylactic cranial irradiation [13], to a total dose of 18–35 Gy in 1.8–2.5 Gy fractions given daily, has become the standard-of-care for selected groups of adult patients with small cell lung cancer and in pediatric leukemia patients to decrease central nervous system relapse [14], with no [15, 16], mild [17, 18], or moderate [19] cognitive impairment. Our studies using a hemibrain irradiation technique demonstrated that a short course of 5×2 Gy X-ray doses significantly reduced the amyloid burden in the brain [3]. Further experiments, which used whole-brain irradiation, demonstrated that the reduction in amyloid plaques was associated with improvements in cognition using the Morris water maze [3].
These results have led to the development of a Phase I study in intermediate stage AD patients at our institution (NCT012359864) that has also been established at Virginia Commonwealth University (NCT02769000), University of Geneva (NCT03352258), and Kyung Hee University Hospital at Gangdong in South Korea (NCT04203121).
Our research to date has been predominantly carried out in male B6.Cg-Tg (APPswePSEN1dE9)85Dbo/J (005864) mice; these double transgenic mice express a chimeric mouse/human amyloid precursor protein (Mo/HuAPP695swe) and a mutant human presenilin 1 (PS1-dE9). In our previous studies, we used mice at 6 months of age for the majority of studies on the effect of radiation on Aβ plaques and 14-month-old animals for our studies on cognition [3]. In this preliminary study we report our initial experience with the B6;129-Tg(APPSwe,tauP301L)1Lfa Psen1tm1Mpm/Mmjax (3xTg-AD) model [20] which is homozygous for the Psen1 mutation and homozygous for the co-injected APPSwe and tauP301L transgenes. The mice in this study were aged to 14 months due to the later onset of tau pathology in this model [20].
MATERIALS AND METHODS
Animal studies
Six female B6;129-Tg(APPSwe, tauP301L)1Lfa Psen1tm1Mpm) (3xTg-AD) mice were purchased from The Jackson Laboratory (Bar Harbor, ME, USA). Mice were housed under standard conditions, maintained on a 12-h dark and 12-h light cycle with ad libitum access to rodent chow and water. The Beaumont Animal Care Committee approved all procedures. Mice were aged to 16 months before undergoing irradiation studies.
Irradiation procedure
Mice were anesthetized with 3% isoflurane inhalation followed by maintenance with 1–3% isoflurane while being irradiated using a Small Animal Radiation Research Platform (SARRP, Xstrahl, Suwanee, GA, USA). The right side of the brain was irradiated with a dose of 2 Gy per day for 5 consecutive days using a 2-beam treatment plan. Treatment beams were delivered using a variable collimator with size adjustments for each mouse. Study endpoint was 8 weeks post radiation treatment.
Tissue harvesting and slide preparation
Mice were euthanized by CO2 inhalation followed by cervical dislocation and decapitation. Whole brains were harvested and allowed to float in 10% Zinc Formalin for 24 h followed by another 24 h in 70% Ethanol. They were then cut with a 1 mm coronal brain matrix for a 40 to 75 g mouse (Ted Pella, Redding, CA). Sections were cut at the bregma –2.12 mm, interaural 1.68 mm at the hippocampus. Both the anterior and posterior brains were placed in cassettes for automated tissue processing starting with an exchange of fresh graded alcohols at 70% through Xylene. Tissue was paraffin embedded and a section was cut and stained with H&E to validate correct anatomical sectioning with equal proportions of hippocampus on the right and left side of the brain.
Immunohistochemistry
For Aβ staining, 4μm thick sections were heated at 65°C for 2 h, followed by deparaffination and rehydration. Slides were rinsed in Tris Buffer Saline followed by antigen retrieval in 3% Formic Acid for 3 min. After rinsing, slides were barcoded and placed onto a Ventana Discovery XT automated IHC (ISH) platform (Roche Diagnostics, Indianapolis, IN). The primary antibody was clone 6E10 (Biolegend, San Diego, CA) diluted to 1:15,000 and incubated for 60 min. Standard IHC followed with a Discovery ultra-map anti mouse HRP (Roche) and Discovery Chromo Map DAB detection kit (Roche).
Tau was stained with monoclonal antibody 5A6 directed against N-terminal Tau residues 16 to 46 [21] which targets microtubule associated protein tau (MAPT) obtained from the Developmental Studies Hybridoma Bank maintained by The University of Iowa. Phosphorylated tau was stained using a polyclonal antibody anti-tau (Thr205) (Santa Cruz Biotechnology, Dallas, TX-this antibody is no longer available). Sections were also stained with antibody AT8 (Invitrogen, Carlsbad, CA) which stains phosphorylated paired helical filament tau at both serine 202 and threonine 205. Antigen retrieval used a heated, high pH (9.0) Tris-based solution for 20 min (Vector Laboratories, Burlingame, CA). 3% hydrogen peroxide was used to terminate any endogenous peroxidase activity. After washing and blocking normal serum with a mouse-on-mouse kit (Vector Laboratories), slides were either incubated with a 1:200 dilution of 5A6 or 1:50 dilution of anti-p-tau or a 1:200 dilution of AT8 for 2 h in a Leica Autostainer XL © (Leica Biosystems, Nussloch, GmbH). Slides were incubated with the Vectastain Elite kit biotinylated secondary antibody anti-mouse IgG (Vector Laboratories) for 30 min, rinsed twice in PBS and Vectastain ABC reagent (Vector Laboratories) was applied for 30 min. The slides were incubated with ImmPACT® DAB Peroxidase (HRP) Substrate (Vector Laboratories) to achieve the desired stain intensity.
Image analysis
Slides were scanned using an Aperio ScanScope AT Turbo (Leica, Buffalo Grove, IL) at×20 magnification. For Aβ analysis, a manual and an automated analysis was carried out. Three blinded observers counted the number of plaques independently of each other in three sections from each mouse brain; these counts were later decoded and respectively averaged to give a single value for each brain. For automated analysis, images were imported into Definiens Tissue Studio software (Definiens Inc., Cambridge, MA). The software uses proprietary Definiens Composer Technology to train and then segment the images into regions of interest (ROIs) based on machine-learning. The software generates many variables and the two most informative are # marker areas which equates to the number of plaques and % IHC stain which represents the total area stained and is a surrogate for plaque burden.
Tau 5A6 stained hippocampal brain slices were analyzed to compare the number of neurofibrillary tangles present in the treated versus untreated hemispheres. Only the hippocampus was counted as it had the most consistent stain pattern between mice. The analysis was carried as described above for manual counting of Aβ plaques and the data averaged to give a single value for each hippocampus side for each mouse.
Statistics
Differences between the treated and untreated sides of the brain were assessed using a two-tailed paired Student’s T-test. The correlation between the tau manual count and the Aβ percent marker area within the same mouse was tested using the Pearson correlation coefficient. An alpha value of 0.05 was considered statistically significant.
RESULTS
Hemi-brain irradiation reduces Aβ plaques in the 3xTg-AD mouse
In some mice plaques were found predominantly in the entorhinal cortex (Fig. 1B) while in other mice the plaques were more prevalent in the hippocampus (Fig. 1A). This is in keeping with previous descriptions of the temporal and spatial description of Aβ plaques in this mouse model [22]. In both examples, the reduction in plaque number and burden on the irradiated side of the brain is clearly visible (Fig. 1). The number of plaques varied considerably with an average value of 10.9 to 68.0 per brain, However, there was a consistent and significant (p = 0.028) reduction in plaque number in the irradiated side of the brain with an average of 40.2±8.9 on the unirradiated side and 23.6±5.2 on the irradiated side (Fig. 1C). Analysis using Definiens software produced comparable results in terms of # marker area although the absolute count of plaques was higher than manual counting (Fig. 1D). The most significant change (p = 0.0022) was in % IHC stain (Fig. 1E) which takes into account both the number of plaques and the area they occupy in the section, i.e., plaque burden.
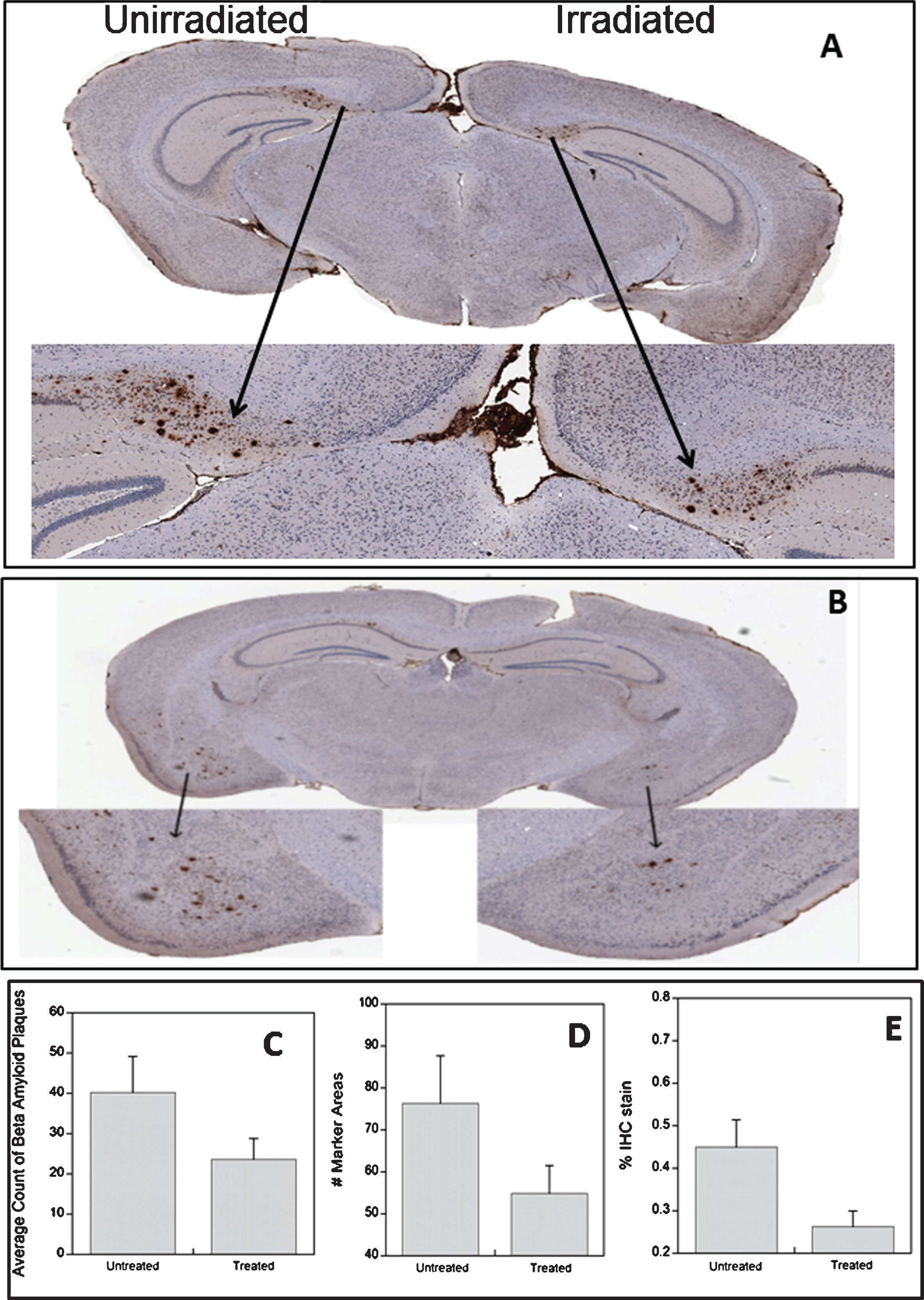
Examples of Aβ staining in the hippocampus (A) and the cortex (B) in different 3xTg-AD mice. The effect of radiation treatment on Aβ plaques asses by manual counting (C), automated counting of plaques (D), and assessment of Aβ plaque burden (E).
Hemi-brain irradiation reduces neurofibrillary tangles in the 3xTg-AD mouse
Figure 2A, B, and C shows typical examples of staining with the three different anti-tau antibodies. Tau staining was predominantly in the hippocampus in the pyramidal neurons of the C1 subfield as previously described [20, 22]. Staining with the 5A6 antibody, which represents total tau staining, produced the cleanest and most distinct staining pattern of pyramidal neurons in the CA1 of the hippocampus and this was selected for quantitation of tau in the hippocampus (Fig. 2A, D). The phospho tau antibody stained tau in neuronal cell bodies and also axonal projections extending into the stratum radiatum (Fig. 2B) while AT8 stained aggregations of paired helical filaments that formed neurofibrillary tangles (Fig. 2C). Again the average number of positive neurons varied between mice from as low as 50 to a maximum of 154. However, each mouse showed a reduction in tau on the irradiated side (Fig. 2C) with the average change being a 20% reduction (p = 0.0024).
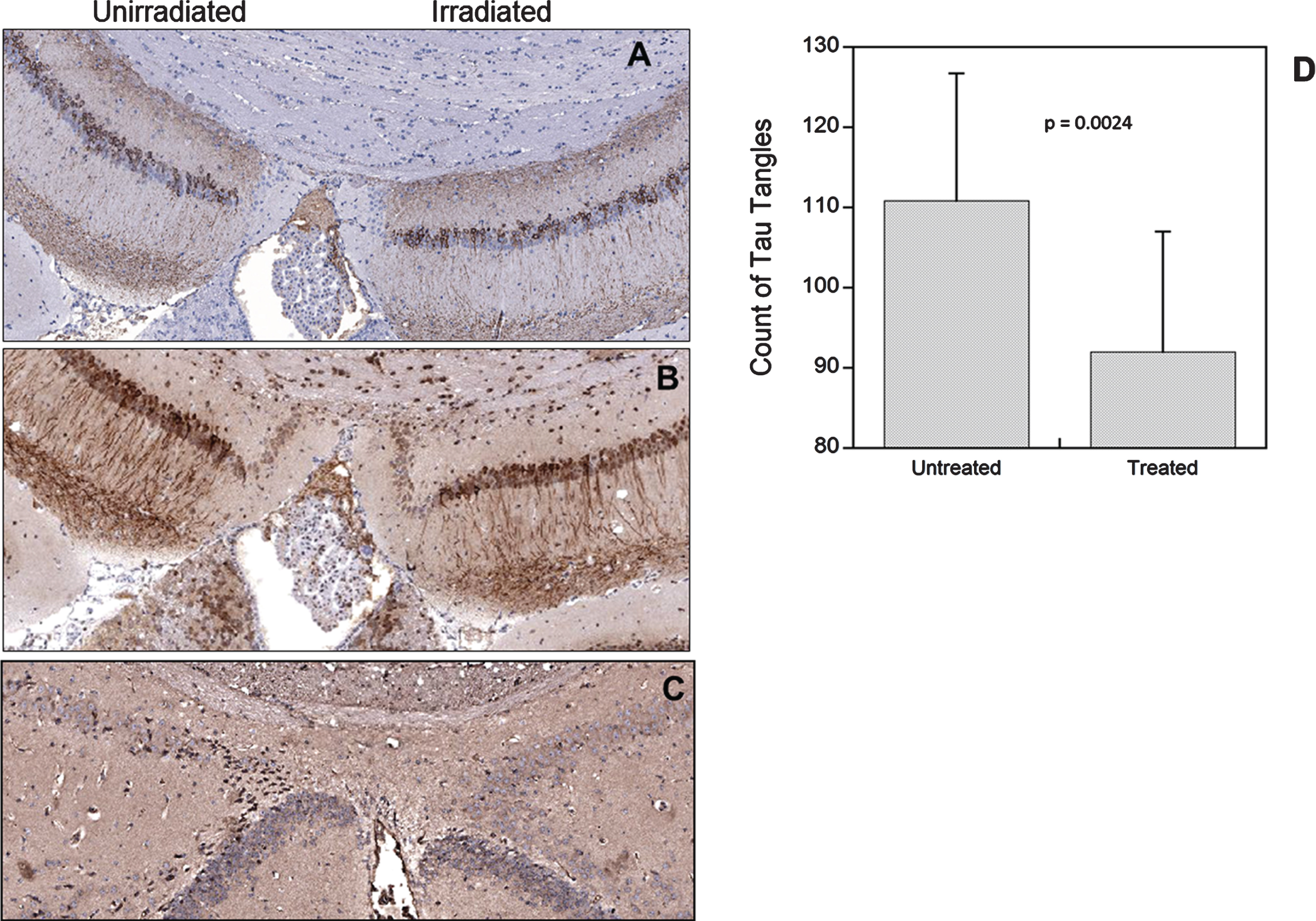
Examples of tau immunohistochemistry stained with the 5A6 antibody (A), the phospho-tau antibody (B), or the AT8 antibody (C). Analysis of 5A6 tau staining in the irradiated and non-irradiated sides of the brain (D).
DISCUSSION
This preliminary study provides evidence that similar low total doses of radiation can not only reduce Aβ plaques but also reduce tau-associated neurofibrillary tangles in the triple transgenic 3xTg-AD mouse model. In the time-scale of the experiment, the effect on Aβ plaques was more dramatic than the effect on tau but the reduction in both was highly correlated (p = 0.0368). There remains a constant debate on the sequence of events that leads to AD. Our data demonstrates a more significant reduction in Aβ plaques and a less pronounced effect on tau in keeping with the amyloid cascade model. However, several lines of evidence suggest that these two pathologies act synergistically. In particular, studies using immunotherapeutic approaches against either Aβ or tau have shown the ability to reduce both the hallmarks of AD and improve cognitive behavior [23–26]. Other non-immune approaches such as inhibition of HDAC3 decreased pathological tau phosphorylation and acetylation, decrease Aβ protein expression in the brain and periphery, increase Aβ degradation and improved learning and memory in the 3xTg-AD model [27]. In another study using mifepristone, a glucocorticoid receptor antagonist, the drug was able to rescue the pathologically-induced cognitive impairment and markedly reduce Aβ-load and tau pathologies [28].
The mechanism by which radiation affects Aβ and now tau remains elusive. The vast majority of literature on radiation effects on the brain comes from either exposure during clinical treatments for brain radiotherapy using high doses or from low doses or dose-rate in the context of environmental radiation exposure. Very few studies [29] specifically address the dose range used in our studies, and none (apart from our own) have studied the effect of radiation in brains affected by AD.
One of the hallmarks of radiation treatment to the brain is neuroinflammation caused by increased expression of cytokines and activated microglia in the neurogenic zone of the dentate gyrus of the hippocampus [30]. In normal brain, radiation induces a pro-inflammatory environment through the activation of inflammation-related genes including nuclear transcription factor kappa B (NF-κB), interleukin-1β (IL-1β), interferon-gamma (INF-γ), and tumor necrosis factor-alpha (TNF-α) [31]. In our previous study [3], we did not observe an increase in IL-1β in the irradiated versus non-irradiated AD brain but there was an increase in the anti-inflammatory cytokine IL-10 on the irradiated side of the brain possibly mitigating immunopathology. This anti- versus pro-inflammatory dichotomy was discussed in a letter in response to our previous article in the APP/PS1 model [2] and is a possible mechanism whereby radiation may be beneficial.
Another established effect of radiation concerns neural stem cells in the subgranular zone of the dentate gyrus which have been shown to undergo apoptosis after low doses of X-rays [32]. It has been shown that X-ray doses of 1 to 30 Gy induce significant apoptosis in the subgranular zone and substantially suppress proliferation [33]. However, there is a subsequent increase in the number of proliferating cells, at lower doses, seven to 14 days post-irradiation. This suggests that lower total doses cause a repopulation response resulting in enhanced neurogenesis which may be a compensatory response and represent an endogenous brain repair mechanism [34].
Another plausible explanation arises from the observations that bone marrow–derived cells of monocyte-macrophage origin effectively home to the central nervous system after irradiation injury [35]. Once in the brain these cells mature, and integrate into the existing cellular microenvironment in various brain regions where there is evidence for functionally relevant brain repair following the radiation injury. Interestingly, this effect is mediated by bone marrow stem cells which are G-CSF-responsive [35]. Recently, several articles have reported that the therapeutic potential of G-CSF in the CNS may ameliorate the neurological disorders including AD [36, 37].
Clearly there is much more research required to elucidate and optimize the potential of low doses of radiation as a treatment for AD. However, the attraction of radiotherapy is that it is a well-established and safe medical modality defined by decades of clinical application and is not hampered by the blood-brain barrier.
DISCLOSURE STATEMENT
Authors’ disclosures available online (https://www.j-alz.com/manuscript-disclosures/20-0030r2).