
Abstract
This article reviews recent advances in the study of microRNAs (miRNAs), long non-coding RNAs (lncRNAs), and their functions in type 2 diabetes mellitus (T2DM), ischemic stroke (IS), and vascular dementia (VaD). miRNAs and lncRNAs are gene regulation markers that both regulate translational aspects of a wide range of proteins and biological processes in healthy and disease states. Recent studies from our laboratory and others have revealed that miRNAs and lncRNAs expressed differently are potential therapeutic targets for neurological diseases, especially T2DM, IS, VaD, and Alzheimer’s disease (AD). Currently, the effect of aging in T2DM, IS, and VaD and the cellular and molecular pathways are largely unknown. In this article, we highlight results from the works on the molecular connections between T2DM and IS, and IS and VaD. In each disease, we also summarize the pathophysiology and the differential expressions of miRNAs and lncRNAs. Based on current research findings, we hypothesize that 1) T2DM bi-directionally and age-dependently induces IS and VaD, and 2) these changes are precursors to the onset of dementia in elderly people. Research into these hypotheses is required to examine further whether research efforts on reducing T2DM, IS, and VaD may affect dementia and/or delay the AD disease process in the aged population.
Keywords
INTRODUCTION
Type 2 diabetes mellitus (T2DM)
T2DM is a frequent type of diabetes, comprising about 90% of all diabetes cases. T2DM is a chronic disease considered by progressive insulin resistance (IR) and hyperglycemia with major complications such as gum disease, nerve damage, foot problems, kidney disease, dental problems, eye disease, sexual and bladder complications, ischemic stroke (IS), and vascular dementia (VaD) [1, 2]. T2DM is the result of a combination of genetic (inheritance), lifestyle (diet and physical activity), and environmental (exposure to toxins) factors. T2DM is a polygenic condition with multiple genes found on different chromosomes, and several environmental factors contribute to the clinical presence of the disorder in people with a genetic predisposition [3]. T2DM is a major risk factor for stroke and related dementia. Persons with diabetes have a stroke that is much worse than people without diabetes. Nevertheless, many of the bypass arteries in people with diabetes are also compromised by atherosclerosis and reduced blood flow to the brain [4]. T2DM is a metabolic disorder and one of the most challenging issues for public health [5].
In 2015, the prevalence of diabetes mellitus was projected at around 415 million people, and this figure is anticipated to rise to 642 million by 2040 (http://www.idf.org/idf-diabetes-atlas-seventh-edition). There are several risk factors that are associated with T2DM, such as increased age, race, lack of body exercise, family history of diabetes, history of gestational diabetes, overweight, unhealthy diet, high blood pressure, poor nutrition throughout pregnancy, and decreased glucose tolerance [6]. Tingling or numbness in the hands and feet, lack of energy, fatigue, excessive thirst and dry mouth, frequent urination, blurred vision, slow healing wounds, and recurrent skin infections are common T2DM-related symptoms [7]. Molecular links between T2DM, IS, and VaD are not completely understood. In the current article, we extensively discussed this issue.
Ischemic stroke
A stroke occurs when the blood supply to part of the brain is interrupted and the subsequent lack of oxygen and nutrients causes brain tissue damage. In a matter of minutes, brain cells begin to die. There would be a lack of memory and muscle function as brain cells die during a stroke [8]. Early action can mitigate brain damage and possible weaknesses. A stroke occurs in two forms: ischemic stroke and a hemorrhagic stroke. An artery that supplies blood to the brain is blocked by a blood clot is known as IS. A broken vessel causes blood to leak into the brain, known as a hemorrhagic stroke. Stroke can cause permanent damage, including facial numbness, impaired vision and speech, partial paralysis, memory, and comprehension. If symptoms last less than one or two hours, it is known as a transient ischemic attack or mini-stroke. The magnitude of the stroke is determined by the degree and location of the injury, varying from mild to catastrophic [9].
Stroke is the second leading cause of death and the third leading cause of disability-adjusted life years worldwide [10, 11]. The risk of having a stroke varies with race and ethnicity. Over the last four decades, the stroke incidence in low- and middle-income countries has more than doubled. Globally, 70% of strokes and 87% of both stroke-related deaths and disability-adjusted life years occur in low- and middle-income countries [12]. According to the World Health Organization, one in 6 people worldwide will have a stroke in their lifetime. Every year, 15 million people worldwide suffer a stroke, and 5.8 million people die from it [13]. Every year, more than 795,000 people in the United States have a stroke, and it kills about 140,000 Americans [14]. Stroke mortality rates are higher for African-Americans than for whites [15].
Stroke represents a clinical syndrome rather than a specific disease since patients with the same clinical demonstration can have unrelated etiopathology, and patients who experience the same type of stroke can have different clinical signs. Stroke can be caused by a number of different pathologies, which all result in a common sudden-onset focal cerebral damage. Current advances in imaging methods, such as computed tomography or magnetic resonance imaging, have become the primary imaging modalities in the initial assessment of acute stroke for several reasons [16]. Stroke clinical features can be grouped into syndromes, which are related to the vascular territories. Most strokes affect the anterior circulation, namely the middle cerebral artery territory [17].
The stroke is a heterogeneous disease, and the risk factor profiles leading to different types and subtypes of stroke vary [18]. There are two types of risk factors associated with stroke: modifiable and non-modifiable. Age, sex, ethnicity, and heredity are considered as non-modifiable risk factors. Hypertension, smoking, cardiovascular disease, diabetes, asymptomatic carotid stenosis, atrial fibrillation, dyslipidemia, unhealthy diet, obesity, physical inactivity, and post-menopausal hormone therapy are established modifiable risk factors for stroke [19]. The estimated stroke risk also increases with the number of modifiable risk factors that co-occur in an individual [20].
Stroke is a multifactorial disease in which heritable, lifestyle and environmental factors equally contribute. In recent years, several genetic studies were conducted to identify genes and/or single nucleotide polymorphisms that were associated with stroke. Further, multiple studies conducted to identify risk factors such as hypertension, diabetes, hyperlipidemia, and VaD that are linked to stroke. But only a few of them were fully proven to influence the susceptibility to the disease, and many of them have generated negative or conflicting results [21].
Vascular dementia
VaD is the second most common form of dementia caused by brain damage from impaired blood flow to your brain [22]. VaD may result from brain damage caused by numerous strokes or minored blood clots in the heart or neck arteries that block a branch of a blood vessel in the brain [23]. VaD is a pathogenetically heterogeneous neuropsychiatric syndrome, mainly characterized by cognitive impairment. Among dementias, it is second by incidence after Alzheimer’s disease (AD). The prevalence of dementia rapidly increases from about 2-3% among those aged 70–75 years to 20–25% among those aged 85 years or more. The prevalence of VaD is higher in men than in women [24]. Confusion, trouble paying attention and concentrating, reduced ability to organize thoughts or actions, a decline in the ability to analyze a situation, problems with memory, restlessness, and agitation, unsteady gait, sudden or frequent urge to urinate, depression, or apathy are the common sign and symptoms associated with VaD.
Many subtypes of VaD have been described. The spectrum includes 1) mild vascular cognitive impairment, 2) multi-infarct dementia, 3) VaD due to a strategic single infarct, 4) VaD due to lacunar lesions, 5) VaD due to hemorrhagic lesions, 6) Binswanger disease, 7) subcortical VaD, and 8) mixed dementia (combination of AD and VaD) [25].
MOLECULAR LINKS: T2DM, IS, AND VaD
Patients with T2DM have up to the threefold increased risk of recurrent stroke [26]. The association between T2DM and IS is bidirectional, and it is not limited to acute IS since T2DM may contribute to more insidious brain damage represented by lacunar infarcts, increasing the risk of dementia and leading to a steeper decline in cognitive function [27]. T2DM adversely impacts neuronal integrity and function after IS and associated dementia. T2DM and IS are common diseases that frequently occur together. Among patients with T2DM, several factors contribute in varying degrees to the overall cerebrovascular risk factors, and several studies have shown that people with diabetes have approximately twice the risk of IS compared with those without diabetes [27, 28].
T2DM is selectively related to the different subtypes of dementia. There is a twofold increased risk of VaD in persons with diabetes [29]. It has become increasingly clear that people with diabetes are more likely to develop dementia in old age than those without the disease. It is also known that adults with T2DM score lower on certain cognitive tests and subtests than adults without diabetes. The prevalence rate of dementia is nine times higher in patients who have had a stroke than in controls [30]. More than 25% of patients have stroke-developed VaD within 3 months of acute IS. Within 4 years following a stroke, the relative risk of incident dementia is 5.5% [31].
The impact of cellular and molecular mechanisms between T2DM and IS and T2DM and VaD remains unknown. There are many questions raised in research linking T2DM, IS, and VaD, which are largely unanswered. Hence, it is important to understand the early events and molecular links among T2DM, IS, and VaD.
We surveyed a large number of published in vitro and in vivo studies on stroke and summarized here, these include 1) the role of miRNAs in T2DM, 2) the role of miRNAs in IS, 3) the role of miRNAs in VaD, 4) the role of lncRNAs in T2DM, 5) the role of lncRNAs in IS, 6) the role of lncRNAs in VaD, and 7) interaction and crosstalk between miRNAs and lncRNAs in T2DM, IS, and VaD.
Pathophysiology of type 2 diabetes
Over the last decade, major advances have been made in our understanding of the pathophysiology of T2DM. Impaired insulin secretion through a dysfunction of the pancreatic β-cell and impaired insulin action through insulin resistance are the two main pathological defects in T2DM [32]. Insulin resistance will result in impaired 1) insulin mediated glucose uptake in the periphery, 2) suppression of hepatic glucose output, and 3) triglyceride uptake by fat. Islet cells will increase the amount of insulin secreted to overcome insulin resistance. Endogenous glucose production is accelerated in patients with T2DM or reduced fasting glucose [33]. Hyperglycemia can impair pancreatic β-cell function and worsen insulin resistance, leading to a spiteful cycle of hyperglycemia, causing a deteriorating metabolic state [34]. Insulin resistance and the associated increase in glucose, causes the pancreas to release more amount of insulin known as hyperinsulinemia. This condition deteriorates pancreatic β-cell function and leads to a reduction in insulin output [35]. Hyperglycemia and the associated inflammatory processes lead to the micro and macro-vascular changes that are seen as complications of T2DM [36]. Increased breakdown of lipids within fat cells, resistance to and lack of incretin, high glucagon levels in the blood, increased retention of salt and water by the kidneys, and inappropriate regulation of metabolism by the central nervous system are the other potentially important mechanisms associated with T2DM and insulin resistance. However, not all people with insulin resistance develop diabetes since an impairment of insulin secretion by pancreatic β-cells is also required [33]. Frequent urination and increased thirst, increased hunger, weight loss, fatigue, blurred vision, and infections are the known symptoms associated with T2DM.
Pathophysiology of ischemic stroke
Ischemic injury includes edema, microvascular thrombosis, programmed cell death (apoptosis), and infarction with cell necrosis. IS causes a hypoxic condition in the cell and depletion of cellular adenosine triphosphate (ATP). ATP is essential to maintain ionic gradients across the cell membrane and cell depolarization. At a cellular level, neurons present in the ischemic region become depolarized as ATP is depleted, and membrane ion-transport systems are unsuccessful. Interruption of cellular metabolism also damages normal sodium-potassium in the plasma membrane pumps, producing an intracellular increase in sodium, which, in turn, upturns intracellular water content. This cellular swelling is referred to as cytotoxic edema and occurs very early in cerebral ischemia [37–39].
Cerebral ischemia condition damages the normal sodium-calcium exchange proteins found on cell plasma membranes. The resulting influx of calcium leads to the release of glutamate (neurotransmitters), which, in turn, activates N-methyl-D-aspartate and other excitatory receptors on other neurons. These particular neurons then become depolarized, causing excess calcium influx, glutamate release, and strengthening the initial ischemic insult. This immense calcium-sodium inflow also activates various degradative enzymes, which leads to the destruction of the cell membrane and other essential neuronal structures [40]. In addition to the neurotransmitters, free radicals, nitric oxide, and arachidonic acid are also generated, which further damages the neurons. The excess level of calcium into cells can initiate cell death signaling cascades, through activation of several degradative enzymes such as proteases, endonuclease, phospholipase, and hydrolytic enzymes [15].
An acute vascular occlusion produces heterogeneous regions of ischemia in the affected vascular territory. This allows clinicians to be able to predict the area of the brain vasculature that can be affected. Middle cerebral artery, anterior cerebral artery, posterior cerebral artery, vertebrobasilar, cerebellar, and lacunar are the arteries commonly affected by the ischemic attack [41]. Cells are present in the affected region, presumed to die within minutes of stroke onset. Decreased or marginal perfusion zones are known as the ischemic penumbra. Tissues in the penumbra can remain viable for several hours because of marginal tissue perfusion [42]. Ischemia also directly results in dysfunction of the cerebral vasculature, with the breakdown of the blood-brain barrier (BBB) occurring within a short period after infarction. After the breakdown of the BBB, proteins and water abundance into the extracellular space cause vasogenic edema [43, 44]. Infarction results in the death of astrocytes as well as the supporting oligodendroglial and microglial cells. Within hours to days after a stroke, specific genes are activated, leading to the formation of cytokines and other factors that, in turn, cause further inflammation and microcirculatory compromise [40].
After a few minutes or even a few seconds following IS, the IS cascade begins and can continue for hours until the disease ceases. A brain ischemia leads to ischemic cell damage and stimulation of the inflammatory process, free radical production, excitotoxicity, disruption of sodium and calcium influx, enzymatic changes, endothelin release, delayed coagulation, activation of platelets and leukocytes and endothelial dysfunction, which are the pathophysiological reactions that separately and/or together contribute to the brain injury resulting in the onset of stroke [15].
Pathophysiologically, IS can be broadly classified as 1) primary vascular pathologies, such as atherosclerosis, aortic arch atherosclerosis, arterial dissection, migraine, or vasculitis, 2) cardiac pathologies, such as atrial fibrillation, myocardial ischemia/infarction, and patent foramen ovale, and 3) hematologic pathologies, such as prothrombotic hypercoagulable or hyperaggregable states [45].
Pathophysiology of vascular dementia
VaD can cause different symptoms depending on where the blood vessels are damaged in the brain. Many people develop VaD after suffering a stroke or multiple small strokes. VaD is sometimes called multi-infarct dementia [15]. The stroke causes parts of the brain to die, which further leads to dementia. Strokes often cause physical symptoms, depending upon what part of the brain is affected, but mini-strokes may happen without obvious external symptoms. VaD most commonly develops when the stroke occurs on the left side of the brain or when it involves the memory center of the brain, the hippocampus. About one-third of people who had no dementia symptoms before the stroke will develop VaD afterwards, usually within a year [46]. High blood pressure, diabetes, high cholesterol, family history of heart problems, heart rhythm abnormalities, overweight, smoking, unhealthy diet, high alcohol consumption, and increasing age are the risk factors associated with VaD.
BLOOD-BASED BIOMARKERS
Blood-based entities will be beneficial for the diagnosis of metabolic syndrome and will help reduce the risk of T2DM, IS, and VaD, contributing to both societal and economic benefits [47]. Clinical approaches accessible for the diagnosis and prognosis of stroke were restricted to radiological imaging, such as computerized tomography and magnetic resonance imaging, which had limited availability and higher cost [48]. There is only one drug available to date, and it predominately targets to eliminate the physical occlusion within the blood vessel. To explore potential therapeutic targets to improve recovery following IS, researchers sought to study the role of miRNAs as peripheral biomarkers for IS [49]. A single biomarker might not be adequate to identify the underlying complexities known to underlie the cellular changes linked to disease and to discriminate diseased from healthy individuals [50]. A biomarker panel that reflects diverse pathophysiological characteristics of a disease or syndrome might be needed to capture the complexities of a particular disease. Biomarker panels have been sought to improve the diagnosis of T2DM, stroke, VaD, and their causes. A biomarker needs to identify a particular feature of disease state as accurately and specifically as possible and to be presented as clearly as possible for use by clinicians [50].
microRNAs
microRNAs are short (17–25 nt) non-coding RNAs that regulate gene translation and primarily have a role in mediating a huge range of biological functions (Fig. 1). The miRNA gene is transcribed to generate a primary microRNA, a precursor molecule that undergoes nuclear cleavage to form a precursor microRNA (pre-miRNA). The pre-miRNA is cleaved in the cytoplasm to create a microRNA duplex containing the mature miRNA. The duplex unwinds, and the mature miRNA assembles into the RNA-induced silencing complex. The miRNA-Ago2 complex targets the complementary seed sequence at the 3’ untranslated region of messenger RNA (mRNA) target genes and leads to either translation inhibition or mRNA degradation [51]. miRNA-mediated gene modulation basically depends on the seed sequence complementarity with the target mRNA [52]. miRNAs are expressed in the cytoplasm and also abundantly in the nucleus. miRNAs regulate transcription and RNA splicing via diverse mechanisms [53].
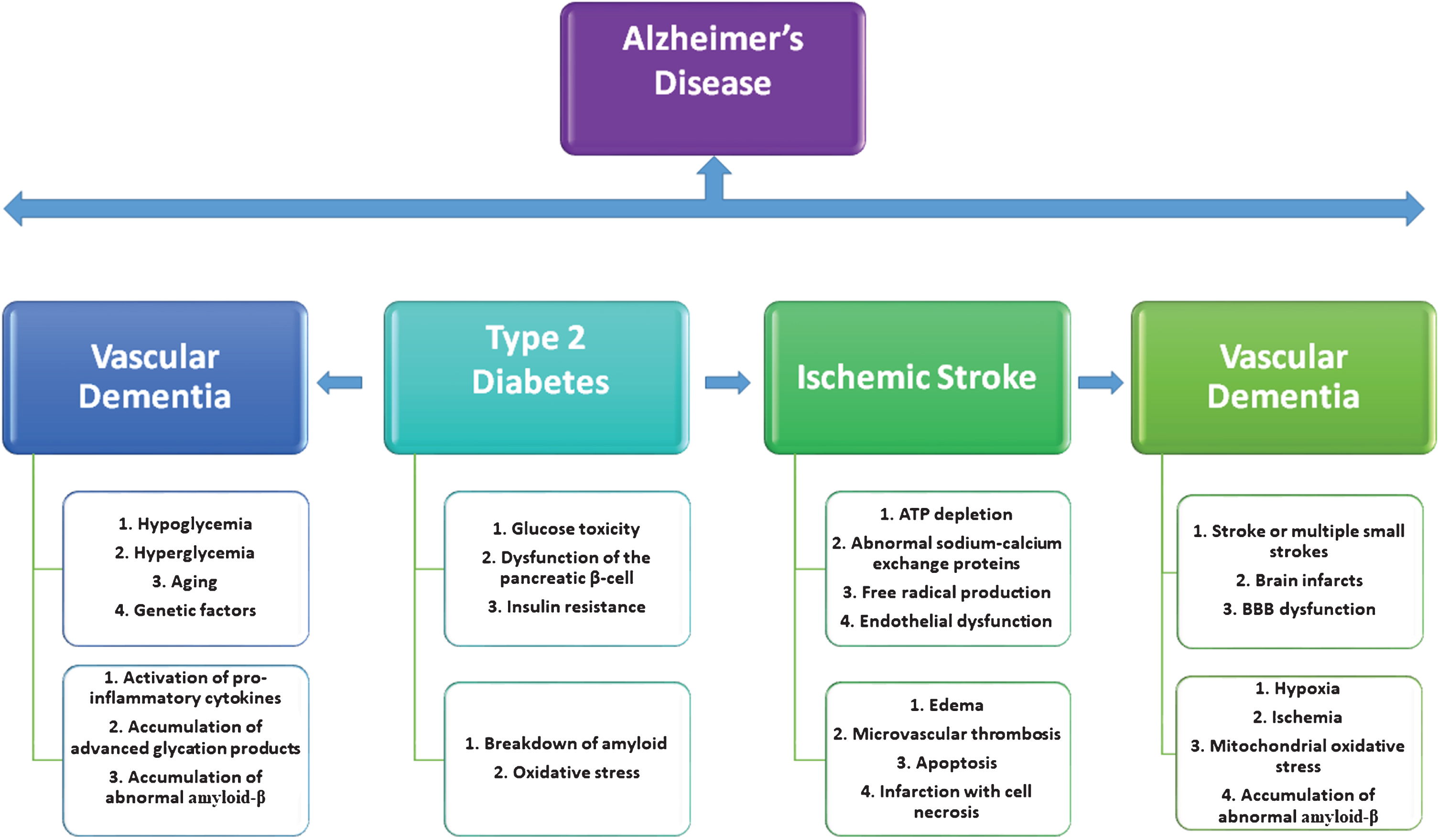
Interactions between type 2 diabetes mellitus, ischemic stroke, and vascular dementia. Type 2 diabetes mellitus increases the risk of ischemic stroke and vascular dementia in the aging population, and all these factors are precursors for the development of Alzheimer’s disease in elderly individuals.
Most mammalian mRNAs are conserved targets of miRNAs, and it can potentially target hundreds of mRNAs [52]. miRNA-mediated reduction of their target mRNA levels is proposed as a major mechanism of miRNA-mediated repression of a target gene expression [54]. miRNAs are composed of a group of endogenous and small non-protein coding genes present in virtually all animals, plants, and some viruses. miRNAs are important regulators of several biological processes, such as cell growth, cell proliferation, embryonic development, tissue differentiation, and apoptosis [50]. miRNAs have also been found to improve the gene-regulatory processes in cerebrovascular diseases, and miRNA-mediated gene silencing controls a diversity of important cellular processes, such as cellular senescence and neuro-regeneration [55–57]. miRNAs have also been found to be important regulators of many diseases, such as diabetes, stroke, cancer, and AD [58–65]. miRNAs might serve as valuable biomarkers due to their easy detection and stability in blood samples [66].
Long non-coding RNAs
Long non-coding RNAs are a relatively abundant component of the mammalian transcriptome and are a large and diverse class of non-coding RNAs with a length of more than 200 nucleotides that do not encode proteins [67]. lncRNAs have been implicated in the regulation of gene transcription, and these can be transcribed as whole or partial natural antisense transcripts to coding genes, or located between genes or within introns [68]. In humans, lncRNAs are thought to incorporate nearly 30,000 different transcripts; hence, its transcripts account for the major part of the non-coding transcriptome. lncRNAs may be classified into different subtypes based on the position and direction of transcription in relation to other genes [69, 70]. Currently, there are very few studies that have been characterized by lncRNAs in detail. However, these lncRNAs are dynamic that have a wide range of roles in cellular and developmental processes and regulating gene expression. Through a range of diverse mechanisms, lncRNAs may carry out both gene inhibition and gene activation. lncRNAs are very useful in understanding the complexity of genomic regulation.
A few lncRNAs have had their functions experimentally defined and have been shown to be involved in fundamental processes of gene regulation [71]. lncRNAs are important regulators of several biological processes, such as chromatin structure and function, direct transcriptional regulation, regulation of RNA processing (splicing, editing, localization, translation, and turnover/degradation), post-translational regulation of protein activity and localization, facilitation of ribonucleoprotein complex formation, modulation of miRNA regulation, gene silencing through production of endogenous siRNA, and regulation of genomic imprinting. lncRNAs can be divided into five different categories: 1) sense lncRNAs, 2) antisense lncRNAs, 3) bidirectional lncRNAs, 4) intronic lncRNAs, and 5) intergenic lncRNAs [72]. The lncRNAs can implement its function in four different ways: 1) signals, 2) decoys, 3) guides, and 4) scaffolds [73]. lncRNAs are predominantly present in nuclear, others nuclear, and/or cytoplasm. The main challenges of working with lncRNAs are low abundantly present (typically lower than mRNA levels) in cells and can be overlapped with coding transcripts on both strands and are often restricted to the nucleus. lncRNAs have also played an important role in many diseases, such as diabetes, cancer, neurodegenerative disease, and respiratory disease [74–76].
Crosstalk and interaction between microRNAs and long non-coding RNAs
To understand the role of miRNA and lncRNAs in 1) neurodegenerative disease, 2) disease developmental stage, and 3) tissue specific mechanisms, it is important to understand the interactions between miRNA and lncRNAs. The number of studies that describe miRNA-lncRNAs regulation is increasing. Recent studies suggest that lncRNAs could play a “sponge/decoy” role, competing for other genes for miRNA binding and, therefore, reduce the regulatory effect of miRNAs on targeted mRNAs [77]. Genomic regions where miRNA transcripts and lncRNAs overlap have dual/multiple functionalities. lncRNAs may act as endo-siRNAs or encode small non-coding RNAs [78]. Different biological processes can either promote the activation of the miRNA biogenesis pathway or trigger lncRNAs function. lncRNAs have been shown to function as “sponges” coordinating the miRNA function, and it takes place in the cytoplasm, while there are some miRNA targeting lncRNAs in the nucleus too [79]. Several studies have identified lncRNAs–miRNAs indirect interactions by advanced experiments that quantify the miRNA effect on mRNA levels or the mRNA effect on miRNA levels [80].
Crosslinking and immunoprecipitation techniques, high-throughput sequencing of RNA isolated by crosslinking immunoprecipitation, photoactivatable-ribonucleoside-enhanced crosslinking and immunoprecipitation, crosslinking, ligation, and sequencing of hybrids have been implemented to determine the complex network of miRNA-lncRNAs regulatory mechanisms [81–83].
Role of microRNAs in type 2 diabetes
Recent studies have highlighted the roles of miRNAs in pancreatic islet development and the execution of specialized β-cell functions, including insulin production, insulin secretion, and the establishment of T2DM [84]. A large number of miRNAs have been implicated in normal pancreatic development and function. Moreover, given the complex interplay between many miRNAs in normal pancreatic cells, it is expected that aberrant miRNA expression or mutations could result in β-cell pathology [85].
Most recently, Wu et al. (2019) [86] investigated the effect of IL6R on the viability and apoptosis of INS-1E cells regulated by miR-22 in T2DM and the underlying mechanism involved. The study indicated that miR-22 strengthened the viability and suppressed apoptosis of INS-1E cells, partly by downregulation of IL6R through the activation of the JAK/STAT signaling pathway [86].
Garcia-Jacobo evaluated the levels of miR-146a, miR-34a, and miR-375 in normal-glycemic individuals, pre-diabetic, and T2D patients in relation to β-cell functionality, insulin resistance, and metabolic parameters. They found that the relative expression of circulating miR-375, miR-34a, and miR-146a were not associated with β-cell function, and their expression was affected by other variables such as glycaemia, obesity, insulin treatment, and the presence of diabetic nephropathy or diabetic foot [87].
miR-3188, one of the earliest discovered microRNAs, is involved in regulating the mTOR-p-PI3K/AKT pathway, thus affecting the progression of diabetic complications. A population-based study from China indicated that the miR-3188 (rs7247237-C>T) polymorphism is associated with the incidence of vascular complications in Chinese patients with T2DM, likely due to its remarkable effect on miR-3188 expression. Further, it is stated that miR-3188 expression upregulated the expression of gstm1 and trib3 proteins, which impairs NO production in human endothelial cells through inactivating the AKT/eNOS signal transduction pathway [88].
Another study explored the role of miR-3666 in T2DM. miR-3666 was significantly downregulated in the serum of T2DM patients when compared to healthy volunteers, and the miR-3666 expression level was negatively correlated with blood glucose levels of T2DM patients. Overexpression of miR-3666 inhibited cell proliferation, reduced insulin secretion, and promoted cell apoptosis of pancreatic β-cell line (INS-1 cells). On the other hand, knockdown of miR-3666 had the opposite effects in INS-1 cells. In addition, miR-3666 negatively regulated the mRNA and protein expression of ADIPOQ. Overexpression of ADIPOQ promoted insulin secretion after glucose stimulation, promoted cell proliferation, inhibited cell apoptosis, and partially abolished the effects of miR-3666 overexpression on insulin secretion, cell proliferation, and cell apoptosis of INS-1 cells. They concluded that miR-3666 inhibited pancreatic cell proliferation, reduced insulin sensitivity, and promoted apoptosis by targeting ADIPOQ [89].
Dong et al. (2019) [90] investigated how miRNA-122-5p regulates the hepatic IR in vitro. They first found that the miR-122 level was upregulated in the liver of rats fed with a high-fat diet and injected with streptozotocin (T2DM rats), while the expression level of insulin-like growth factor 1 receptor (IGF-1R), a potential target of miR-122, was downregulated in the diabetic liver. An increase of miR-122 and a decrease of IGF-IR levels were observed in IR hepatic cells in vitro, which was the same as that in the diabetic liver. In addition, the PI3K/Akt pathway was deactivated in IR cells and then reactivated with miR-122 inhibitor transfection. They concluded that miR-122 is able to regulate IR in hepatic cells by targeting IGF-1R [90].
Another study evaluated miRNA-21 as a potential biomarker for the risk of developing diabetes in adults with impaired glucose tolerance (IGT) and to investigate its downstream effects. They observed significant upregulation of miRNA-21 in IGT and in T2DM subjects, and miRNA-21 was positively correlated with glycemic parameters. These data demonstrated that miRNA-21 is associated with prediabetic status and exhibits a predictive value for early detection of glucose imbalances. Further, these data could provide novel clues for miR-based biomarkers to evaluate diabetes [91].
Sucharita and the research team aimed to compare the circulating levels of the miRNA (miR9, miR30d, miR1, miR133a, miR29a, and miR143) between T2DM and gender-matched controls and also to evaluate the strength of association between circulating miRNAs and β-cell function/IR among Indians with T2DM. They have used 30 T2DM (25–60 years), and their gender-matched controls (n = 30) and estimated plasma glucose and insulin, HbA1c, lipid profile, and miRNA levels. They have demonstrated that miR30d (insulin gene transcription in the pancreatic β-cell and regulator of insulin sensitivity in skeletal muscle) was overexpressed among T2DM [92].
The dysregulation of alpha-cells is associated with transcriptome changes. Dusaulcy et al. (2018) hypothesized that miRNAs, which are negative regulators of mRNA stability and translation, could be involved in alpha-cell alterations or adaptations during T2DM [93]. They have performed miRNA microarray analyses on pure alpha- and β-cells from high-fat diet fed obese hyperglycemic mice and low-fat diet fed controls. From the microarray results, 16 miRNAs were significantly regulated in alpha-cells of obese hyperglycemic mice and 28 in β-cells. From these findings, they have concluded that the downregulation of miR-132-3p in alpha-cells of obese diabetic mice may constitute a compensatory mechanism contributing to keeping glucagon-producing cell number constant in diabetes [93].
Another in vivo study aimed to investigate the role of miR-125b-5p in pancreatic β-cell function and insulin sensitivity of mice with T2DM with the involvement of Dishevelled antagonist Dapper1 (DACT1) and the c-Jun NH2-terminal kinases (JNK) signaling pathway [94]. They demonstrated that mice with T2DM had lower pancreatic β-cell function and insulin sensitivity, as well as diminished expression of miR-125b-5p but enhanced expressions of DACT1, JNK, and c-Jun. miR-125b-5p inhibited DACT1 expression and the activation of the JNK signaling pathway, as well as restrained cell proliferation and promoted cell apoptosis. They further suggested that upregulated miR-125b-5p promotes insulin sensitivity and enhances pancreatic β-cell function through inhibiting the JNK signaling pathway by negatively mediating DACT1 [94].
Pastukh et al. (2019) [95] recruited 40 human subjects: 10 healthy subjects, 10 with diabetes but without retinopathy (NDR), 10 with diabetic non-proliferative retinopathy (NPDR), and 10 with proliferative diabetic retinopathy (PDR). They examined seven specific miRNA candidates (miR-126-3p, miR-130a-3p, miR-21-1, let-7f-5p, miR-122, miR-30c, and miR-451a) whether these serum miRNAs show altered levels at different stages of diabetic retinopathy. From these, a significant increase in miR-122 was found in patients with diabetic retinopathy that may be related to its role in preventing angiogenesis and proliferation [95]. Another study from China stated that lower serum miR-122 is a unique feature of HNF1A-DM patients and might partially explain the increased risk for liver neoplasm and abnormal lipid metabolism in HNF1A-DM patients [96].
Liang and the research group used RNA-seq and qRT-PCR to identify differential serum miRNAs in patients with T2DM with diabetic retinopathy (DR) [97]. The results suggested that a profile of five serum miRNAs (hsa-let-7a-5p, hsa-miR-novel-chr5_15976, hsa-miR-28-3p, hsa-miR-151a-5p, hsa-miR-148a-3p) was significantly associated with T2DM-DR [97].
Overall, findings from the above studies provide information about miRNAs and their mRNA targets. These studies also suggested that miRNAs are novel therapeutic drug targets for T2DM.
Role of microRNAs in ischemic stroke
Mounting evidence has shown that altered miRNAs are implicated in the pathologies of IS.
Recently, our laboratory conducted a preliminary global miRNA analysis using a large number of serum samples from IS patients and control subjects and identified the significant number of differentially expressed miRNAs in IS patients compared to controls. We used the Illumina deep sequencing method, and we obtained a total of 484,651,777 raw RNA reads. Among these, 341,678,616 (70.5%) were mappable reads for miRNAs, 39,890,853 reads were mapped to mRNA, and 24,723,087 reads were mapped to other RNAs (RFam: rRNA, tRNA, snRNA, snoRNA, and others). miRNAs sequencing analysis revealed/detected a total of 4,656 miRNA serum samples of IS patients versus healthy controls. Among them, 272 miRNAs were significantly deregulated. Interestingly, we found 173 miRNAs were significantly upregulated, while 76 were found to be significantly downregulated in IS patients. Based on the miRNAs reads and their fold change (±2-fold) values, 16 most significant deregulated miRNAs were selected for further validation. Interestingly, our study identified two potential and previously unreported miRNA candidates (PC-3p-57664, PC-5p-12969) its and two previously reported miRNAs (hsa-miR-122-5p_R-1, hsa-miR-211-5p_1ss18CT) were significantly upregulated in patients compared to controls, while seven miRNAs (hsa-miR-22-3p, PC-3p-32463, hsa-miR-30d-5p, hsa-let-7f-5p, hsa-miR-186-5p, hsa-miR-23a-3p, mmu-miR-5124a) were downregulated in patients compared to controls. Receiver operating characteristic curve analysis revealed a significant area under curve (AUC) value for PC-3p-57664, PC-5p-12969, hsa-miR-122-5p, and hsa-miR-211–5p [58].
Based on a global miRNA sequencing analysis, we found four previously unreported and novel miRNAs in IS patients [59]. Among these, miRNA PC-5P-12969 was exclusively expressed in the IS condition; otherwise, it was not expressed in normal condition and, therefore, we focused on miRNA PC-5P-12969 for further studies. We investigated novel miRNA PC-5P-12969 for its expression levels using quantitative real-time PCR assay (qRT-PCR) in an in vitro, oxygen-glucose deprivation/reoxygenation (OGD/R) treated mouse primary hippocampal neuronal cells (HT22) and in an in vivo using a photothrombotic stroke model [59]. In an in vitro study of stroke-induced HT22 cells, we found a 2-fold increase of PC-5P-12969 expression levels, in agreement with our original global miRNA study. In the cerebral cortex of photothrombotic stroke mice, we found significantly upregulated levels of PC-5P-12969 in 4-hour and 1-day post-stroke relative to the control mice. However, we did not find any change in the expression of PC-5P-12969 in the cerebellum (unaffected in IS) of both stroke and control mice. Based on findings from this study, together with our earlier original global microRNA study results, we concluded that PC-5P-12969 is a potential therapeutic target for IS. This was the first study, which validated that the miRNA PC-5P-12969 might be a potential biomarker for IS. Moreover, target genes prediction of the co-expressed miRNAs and further Gene Ontology and Kyoto Encyclopedia of Genes and Genomes analyses were performed, which revealed that PC-5P-12969 and its mRNA targets were involved in the regulation of the stroke event [59].
Most recently, Ge et al. (2019) [99] investigated the functional role of miR-19a-3p in cerebral ischemic injury and explored its underlying regulatory mechanism. The expression level of miR-19a was significantly downregulated and upregulated in rat neurons and astrocytes, respectively. A significantly elevated level of miR-19a-3p was found in ischemic/reperfusion (I/R) and OGD models in comparison to control groups. Expression of the glycolysis enzyme markers, apoptosis-related factors levels, apoptosis, glucose uptake, and lactate production was significantly repressed by both I/R and OGD. Further, they predicted and confirmed ADIPOR2 to be a direct target of miR-19a-3p. Collectively, their results indicated that elevated miR-19a-3p mediates cerebral ischemic injury by targeting ADIPOR2 [99].
Another study from China investigated the function of miR-145 in an in vitro OGD model [100]. In their initial studies, they have found that miR-145 and its predicted target, erythropoietin-producing hepatoma receptor A4 (EPHA4), was closely associated with IS. Further, miR-145 regulates the expression of EPHA4 at both the mRNA and protein levels. In an in vitro OGD model, miR-145 employed a protective role by downregulating EPHA4, which suggests their potential roles in IS [100].
miRNAs can simultaneously control many mRNA targets. Likewise, miR-145-5p directly inhibited dual specificity phosphatase 6 (DUSP6). Wu et al. (2019) investigated the effects of miR-145-5p in regulating myocardial ischemic injury. The results indicated that DUSP6 was beneficial against I/R injury through inhibiting c-Jun N-terminal kinase pathways. In conclusion, the essential roles of miR-145-5p and DUSP6 in I/R provide a novel therapeutic target to develop future intervention strategies for cerebral injury [101]. miR-145-5p is known as a putative regulator of Nuclear Receptor Subfamily 4 Group A Member 2, and in a middle cerebral artery occlusion/reperfusion model, anti-miR-145-5p administration promoted neurological outcomes in the rat [102].
In many previous in silico and miRNA microarray studies [103, 104], differentially expressed miRNAs were identified in cerebral I/R injury. However, the relationship between miRNA and cerebral I/R injury remains largely unknown.
Deng and the research team investigated the function and explored the possible mechanism of miR-224-3p in cerebral I/R injury [103]. Overexpression of miR-224-3p alleviated the apoptosis induced by OGD, and cleaved caspase-3 was significantly reduced. They further provided the possible mechanism that miR-224-3p may protect cells from cerebral I/R injury by targeting FAK family-interacting protein (FIP200). Further rescue experiments proved that overexpression of FIP200 partially blocked the effect of miR-224-3p. They concluded that miR-224-3p might serve as a potential target for cerebral ischemia [103].
Burek et al. (2019) [104] analyzed the expression of miR-212/132 in hypoxic mouse and human brain microvascular endothelial cells (BMEC) as well as in posttraumatic mouse and human brain tissue and serum exosomes. miR-212/132 expression was detected in brain capillaries by in situ hybridization and was increased up to ten times in hypoxic BMEC. They identified and validated tight junction proteins claudin-1 (Cldn1), junctional adhesion molecule 3 (Jam3), and tight junction-associated protein 1 (Tjap1) as potential miR-212/132 targets. Overexpression of miRs led to a decrease in mRNA and protein expression of Cldn1, Jam3, and Tjap1, which could be rescued by a respective anti-miRNA [104].
It is known that cyclic adenosine monophosphate-degrading enzyme phosphodiesterase 3 (PDE3) is an important treatment target, which may be modulated by miRNAs important for regulating gene expression [105]. HCMEC/D3 cells transfected with miR-27a-3p and miR-222-3p mimics had reduced relative expression of PDE3A protein. PDE3A-related miRNAs miR-221/miR-222 and miR-27a/miR-27b/miR-128 were potentially linked to pathways essential for immune regulation as well as cerebral and vascular integrity/function. Furthermore, relative PDE3A protein expression was reduced by miR27a-3p and miR-222-3p [105].
Another study intended to investigate the effects of miR-212 on the recovery function and vascular regeneration of endothelial progenitor cells by inactivation of the Notch signaling pathway by binding to matrix metallopeptidase 9 (MMP9) in mice with IS [106]. MMP9 was predicted as a gene related to IS, and miR-212 is a potential regulating mRNA of MMP9 [106]. Another miRNA, miR-539, targeted MMP-9, and further regulated the BBB permeability in cerebral I/R injury [107]. miR-7 mimics may therapeutically minimize stroke-induced brain damage and disability by repressing α-synuclein [108]. Another important miRNA in post-stroke angiogenesis is miRNA-27b. Yuan and the research team investigated the regulatory effects of miRNA-27b on post-stroke angiogenesis. In primary cultured BMECs, the inhibition of miRNA-27b induced the activation of adenosine monophosphate-activated protein kinase (AMPK), which increased tube formation and migration. This action was attenuated when AMPKα2 was knocked down [109].
Progesterone (P4) is a potent neuroprotectant and a promising therapeutic for stroke treatment. Singh and his colleagues recently reported that the miRNA let-7i is a negative regulator of membrane-associated progesterone receptor (Pgrmc1) and brain-derived neurotrophic factor (BDNF) in glia and that let-7i disrupts P4-induced BDNF release and P4’s beneficial effects on cell viability and markers of synaptogenesis [110]. Further, they used the in vivo model of ischemia, demonstrating that inhibiting let-7i enhances P4-induced neuroprotection and facilitates functional recovery following stroke [110].
Gamdyzk and the team investigated the role of PPAR-β/δ as well as its molecular mechanisms in neonatal hypoxia-ischemia (HI). They hypothesized that PPAR-β/δ agonist GW0742 could activate miR-17-5p, consequently inhibiting the TXNIP and ASK1/p38 pathway leading to attenuation of apoptosis. In their results, they found that intranasal administration of PPAR-β/δ agonist GW0742 attenuated neuronal apoptosis at least in part via PPAR-β/δ/miR-17/TXNIP pathway [111].
Recent research from Verma and his team used whole microRNAome analysis and identified miR-141-3p as a unique miRNA that was significantly upregulated in isolated mice in a time-dependent manner up to 2 weeks after stroke [112]. There was a significant upregulation of Tgfβr1 (transforming growth factor beta receptor 1, a direct target of miR-141-3p) and Igf-1 (insulin-like growth factor 1) mRNA after treatment with miR-141-3p antagomiR. Further, they have concluded that miR-141-3p is increased with post-stroke isolation. Inhibition of miR-141-3p improved mortality, neurological deficits, and decreased infarct volumes. Importantly, these therapeutic effects occurred in aged animals [112].
Janus tyrosine kinase (JAK) and signal transducer and activator of transcription (STAT) of the JAK2/STAT3 pathway play important roles in the downstream signal pathway regulation of IS-related inflammatory neuronal damage. A recent study suggested that the upregulation of miR-216a, which targets JAK2, could induce neuroprotection against ischemic injury in an in vitro and in vivo, which provides a potential therapeutic target for IS [113].
BBB disruption caused by reperfusion injury after IS is an intractable event conducive to further injury [114]. Brain pericytes play a vital role in maintaining BBB integrity by interacting with other components of the BBB. Recent studies have investigated the effect of miRNA in BBB permeability, neuronal apoptosis, and microglia activation. Wan et al. (2018) [115] described that miR-149-5p could regulate ischemia-induced BBB permeability by directly targeting Sphingosine-1-Phosphate Receptor 2. Upregulation of miR-149-5p significantly decreased BBB leakage and improved the outcome after IS. These findings suggest that miR-149-5p may be a potential therapeutic target in the treatment of BBB disruption after stroke [115].
Overall, findings from the above studies are useful and provide new information about miRNAs (overexpression or inhibition) and its mRNA targets. These studies suggested that miRNAs may be developed as a novel therapeutic strategy to treat cerebral ischemia injury and important in the treatment of IS.
Role of miRNAs in vascular dementia
VaD is the second most common form of dementia in elderly people. However, little is understood about the role of miRNAs involved in cognitive impairment in early VaD. miRNAs are versatile regulatory molecules that influence almost every aspect of BBB structure and function. miRNAs are involved in regulating biological processes such as apoptosis and inflammation, which can exacerbate BBB dysfunction. Although much progress has been made, the role of miRNAs in BBB function remains poorly characterized. Understanding the mechanisms of miRNAs in regulating BBB structure and function may lead us to discover novel pharmaceutical targets for the development of effective microRNA-based therapies against VaD and other CNS disorders [116].
Ren et al. (2018) [117] established a VaD model induced by chronic cerebral ischemia and fetal bovine serum (FBS)-free cell model that detects synapse formation to investigate the function of miRNAs in early VaD. The microarray analysis showed that miR-210-5p increased significantly in the hippocampus of rats with 4 weeks of ischemia. The overexpression of miR-210-5p decreased the synaptic number in primary hippocampal neurons, whereas specific suppression of miR-210-5p resulted in the formation of more synapses. Further, miR-210-5p decreased the luciferase activities of 3’UTR of Snap25 mRNA. They have concluded that miR-210-5p contributes to cognitive impairment in chronic ischemia-induced VaD model through the regulation of Snap25 expression, which potentially provides an opportunity to develop a new therapeutic strategy for VaD [117].
Chronic cerebral hypoperfusion (CCH) is a high-risk factor for VaD. miR-96 expression was assessed in a CCH rat model was established by two-vessel occlusion (2VO). The level of miR-96 was significantly increased in 2VO rats, and inhibition of miR-96 ameliorated the cognitive impairment induced by 2VO. miR-96 may play a key role in autophagy under CCH by regulating mTOR; therefore, miR-96 may represent a potential therapeutic target for CCH [118]. Another study clearly stated that miRNA 933 expression by endothelial cells was increased by 27-Hydroxycholesterol and was more prevalent in plasma from dementia patients [119].
Toyama and research team sought key TNFα-responsive miRNAs that might influence BBB breakdown via cerebral tight junction disruption in vascular cognitive impairment [120]. Using a mouse model of vascular cognitive impairment, chronic cerebral hypoperfusion within the white matter was induced with bilateral common carotid artery stenosis surgery. miR-501-3p was upregulated by TNFα in vitro and was upregulated in the white matter after BCAS surgery. Further, miR-501-3p directly bound to the 3’-untranslated region of human ZO-1 and downregulated transendothelial electric resistance. For the first time, they provided evidence that the TNFα-miR-501-3p-ZO-1 axis plays an important role in the pathogenesis of cerebral hypoperfusion-induced working memory deficits and white matter lesions, as a result of BBB breakdown via tight junction disruption [120].
miR-132 might participate in the downregulation of MeCP2 after CCH, and MeCP2 downregulation was possibly involved in the cognitive deficit through regulation of BDNF and its downstream pathways after 2VO [121]. Plasma miR-409-3p, miR-502-3p, miR-486-5p, and miR-451a could be used to differentiate small vessel VaD patients from healthy controls [122]. miR-195 was involved in CBH-induced dendritic degeneration and neuron death through activation of the N-APP/DR6/caspase pathway [123]. miR-195 downregulation in chronic brain hypoperfusion involved PP2A inactivity, which was mediated by the post-transcriptional regulation PME-1, AβPP, and β-site AβPP cleaving enzyme 1 expression [124]. Downregulation of miR-195 plays a key role in the increased vulnerability to dementia via the regulation of multiple targets following chronic brain hypoperfusion [125].
Ragusa et al. (2016) [126] hypothesized that differentially expressed circulating miRNAs in plasma from VaD patients could be applied to diagnose VaD through liquid biopsies. miR-10b, miR29a-3p, and miR-130b-3p were discovered and validated as significantly downregulated in circulating miRNAs in VaD patients compared to unaffected controls. They concluded that these reported miRNAs pave the way to translational applications to molecular VaD diagnosis, but they also should allow to further investigate its molecular bases [126].
Overall, the findings from the above studies indicated that miRNAs are potential targets for the treatment of VaD.
Role of long non-coding RNAs in type 2 diabetes
A large number of studies have revealed that lncRNAs play a vital role in the development of T2DM [127]. Ge et al. (2019) [128] designed a study to investigate the underlying function and mechanism of a novel lncRNA GAS5 in the progression of diabetic nephropathy (DN). They found that lncRNA GSA5 suppressed the development of DN in vivo [128].
Another study was designed to investigate the expression levels of SNHG17 and TTC28-AS1 in T2DM patients. They found that SNHG17 and TTC28-AS1 were downregulated in the T2DM group compared to the healthy control group. The logistic regression revealed that body mass index, systolic blood pressure, fasting blood glucose, and TTC28-AS1 expression substantially affect T2DM susceptibility. Furthermore, expression of SNHG17 was negatively correlated with high-density lipoprotein cholesterol, and expression of TTC28-AS1 was positively correlated with low-density lipoprotein cholesterol. They have concluded that decreased expressions of lncRNAs TTC28-AS1 and SNHG17 in T2DM are possibly associated with the development of T2DM [219].
lncRNA ANRIL has an influence on T2DM complicated with acute myocardial infarction (MI). To check this, Zhang and Wang. (2019) [130] used 1) peripheral blood from patients, and 2) a streptozocin-induced T2DM mouse model. They found that ANRIL was upregulated in peripheral venous blood from patients with T2DM-MI and in myocardial tissues from the established T2DM-MI model mice. Moreover, ANRIL overexpression caused myocardial fibrosis and myocardial cell apoptosis, and it increased the expression of the myocardial fibrosis-related proteins TGF-β1, collagen I, and collagen III in the T2DM-MI mice. However, ANRIL knockdown exerted the opposite effects [130].
A recent meta-analysis study was conducted to elucidate the diagnostic values of lncRNAs in patients with T2DM. They have included the studies concerning the association between lncRNAs expression and diagnostic outcomes in T2DM patients. The outcome of this study indicated that lncRNAs might serve as promising indicators for the diagnostic evaluation of T2DM patients [131].
A population-based study from Egypt aimed to evaluate a panel of circulating hyperglycemia-related lncRNAs: RNCR2, NEAT2, CDKN2B-AS1, and PVT1 in T2DM patients with/without DR. They used 130 T2DM patients and 108 controls. The expression pattern of lncRNAs demonstrated good diagnostic performance in differentiating T2DM from controls and DR [132].
Li et al. (2018) [133] designed a study to investigate the effect of lncRNA HOTAIR on hepatic IR and to explore the possible underlying mechanism. HOTAIR was significantly upregulated in T2D patients, C57BL/6J mice fed with a high-fat diet and db/db mice. However, SIRT1 expression presented an opposite changing trend. Moreover, upregulated HOTAIR remarkably promoted hepatic IR via the AKT/GSK pathway, which could be reversed by SIRT1 overexpression [133].
Erfanian Omidvar and the research team used 100 healthy controls and 100 T2DM patients peripheral blood mononuclear cells to investigate the association between the expression levels of lncRNAs VIM-AS1, CTBP1-AS2, and T2DM susceptibility. The results displayed that decreased levels of VIM-AS1 and CTBP1-AS2 in PBMC were associated with diabetes in the Iranian population [134].
Decreased expressions of lncRNAs LINC00523 and LINC00994 in T2DM are possibly associated with the pathogenicity of T2DM in the Iranian population [135]. Another study provided evidence of the significant association between H19 rs217727-TT and MEG3 rs7158663-AA genotypes with T2D susceptibility [136]. lncRNA MALAT1 was implicated in the improvement of renal function after duodenal-jejunal bypass through regulation of its downstream targets SAA3, IL-6, and TNF-α [137]. lncRNA NONRATT021972 increased and was positively associated with neuropathic pain scoring in T2DM. lncRNA NONRATT021972 exacerbated neuropathic pain via TNF-α related pathways [138].
Overall, the findings from the above studies described the differential expression profiles of lncRNAs in T2DM and also indicated that lncRNAs are potential targets for the treatment of T2DM.
Role of long non-coding RNAs in ischemic stroke
Long non-coding RNAs have been proven to be critical gene regulators of development and disease [139]. lncRNAs are endogenous molecules that play key roles in the pathophysiology of cerebral ischemia and are involved in neuronal cell death [140]. Stroke-induced lncRNAs may also interact with chromatin-modifying proteins and modulate genes associated with ischemic brain damage [67]. lncRNAs and it targets could affect the genetic machinery at molecular levels, and exploring their functions and mechanisms may be a favorable preference for IS treatment. The regulatory role of lncRNAs in different brain cell types, such as neuronal cells, vascular endothelial cells, and microglial cells involved in IS [141]. However, whether lncRNAs are involved in the pathogenesis of IS still remains largely unknown. This part of the review will elucidate the role of lncRNAs in IS.
For the first time, Deng et al. (2019) [142] investigated the role of lncRNA Nespas in IS middle cerebral artery occlusion (MCAO) and OGD. They found expression of Nespas was significantly increased in ischemic cerebral tissues, and OGD treated BV2 cells in a time-dependent manner. Interestingly, the silencing of Nespas aggravated MCAO induced IR injury and cell death. In addition, NF-κB (nuclear factor-κB) signaling activation was inhibited, and transforming growth factor β–activated kinase 1 (TAK1) activation was significantly suppressed by Nespas overexpression. They have concluded that Nespas played an anti-inflammatory and antiapoptotic role in cultured microglial cells after OGD stimulation and in mice after IS by inhibiting TRIM8-related K63-linked polyubiquitination of TAK1 [142].
lncRNA H19 (H19) is one of the earliest discovered long non-coding RNAs. H19 induced the onset of IS through regulating neuronal autophagy and microglial polarization. The relationship between lncRNA and neurogenesis after IS is unclear. To answer this, a research group from China aimed to study whether H19 participated in the neurogenesis process after IS. They have demonstrated that H19 prevents the process of neurogenesis after IS through the p53/Notch1 pathway and strengthens the novel role of H19-based therapy for IS [143]. Huang et al. (2019) [144] attempted to delineate the association of lncRNA H19 polymorphisms with the susceptibility and clinical features of IS in the southern Chinese Han population. Their findings suggested that the lncRNA H19 level may affect the occurrence of IS, and lncRNA H19 variants may influence blood pressure, coagulation function, and homocysteine metabolism of patients with IS in the southern Chinese Han population. lncRNA-H19 promoting Acid phosphatase 5 protein expression contributed to atherosclerosis and increased the risk of IS [145].
Another study suggested that H19 rs217727 gene polymorphism contributes to small vessel IS susceptibility in the Chinese Han population and may serve as a potential indicator for IS susceptibility [146]. H19 promotes neuroinflammation by driving HDAC1-dependent M1 microglial polarization, suggesting a novel H19-based diagnosis and therapy for IS [147].
Small nucleolar RNA host gene 12 (SNHG12) is one type of lncRNA, and it was reported to be highly upregulated in brain microvascular endothelium after cerebral ischemia. Molecular mechanisms of SNHG12 in regulating autophagy during cerebral (I/R) injury remain unclear. SNHG12 expression was upregulated by cerebral I/R in mice and SH-SY5Y cell model after OGD/R, and it alleviated OGD/R-induced SH-SY5Y cell injury, autophagy activation. On the contrary, the downregulation of SNHG12 inhibited autophagy. These findings suggested that SNHG12 as an autophagy inducer alleviates cerebral I/R injury, which might be a new therapeutic target of IS [148].
lncRNA expression profiles and their roles in specific cell types after stroke have not been studied well. The activation of microglial cells plays an important role in neuroinflammation after IS. Inhibiting the activation of microglial cells has been suggested as a potential therapeutic approach in the treatment of IS. lncRNA-1810034E14Rik plays an anti-inflammatory role in IS and regulates p65 phosphorylation, making it a potential target for stroke treatment [149].
lncRNA, GM15628, is highly expressed in pro-inflammatory monocyte-derived macrophages (MoDMs) but not in microglia-derived macrophages (MiDMs), and are functionally related to its neighbor gene, lymphocyte cytosolic protein 1 (LCP1), which plays a role in maintaining cell shape and cell migration. This lncRNA is otherwise called as Macrophage contained LCP1 related pro-inflammatory lncRNA (Maclpil). Maclpil inhibition reduced the migration and phagocytosis ability of MoDMs by inhibiting LCP1. Furthermore, the adoptive transfer of Maclpil silenced M (LPS) reduced ischemic brain infarction, improved behavioral performance, and attenuated penetration of MoDMs in the ischemic hemisphere [150].
Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) is the lncRNA contributing to protecting the BBB after stroke. Polydatin (PD) is a small molecule and natural product that may upregulate MALAT1. PD enhanced MALAT1 gene expression in rat brain microvascular endothelial cells, reducing cell toxicity, and apoptosis after OGD. C/EBPβ is a key transcription factor for PD-mediated MALAT1 expression. PPARγ activity is involved in MALAT1 protective effects through its coactivator PGC-1α and the transcription factor CREB. The beneficial effects of PD involve the C/EBPβ/MALAT1/CREB/PGC-1α/PPARγ pathway, which may provide a novel therapeutic strategy for IS [151].
Another research group focused on evaluating the effect of MALAT1 and MDM2 on IS through the regulation of the p53 signaling pathway. Their results indicated that the MALAT1/MDM2/p53 signaling pathway axis might provide a more effective clinical therapeutic strategy for patients with IS [152]. Cerebral microvascular endothelial cells have spontaneous Kruppel-like family of transcription factor 4 (KLF4) upregulation after ischemic insult, which acts as a protective mechanism to reduce cell apoptosis via transcriptionally activating MALAT1 [153].
Zinc finger antisense 1 (ZFAS1), a newly identified lncRNA, is transcript antisense to the 5’ end of the gene Znfx1. Wang et al. (2018) [154] performed a study to detect the expression of ZFAS1 in 176 IS patients and 111 healthy controls. The results showed that the level of ZFAS1 in IS patients was significantly lower than controls. Furthermore, we found that the ZFAS1 levels in large-artery atherosclerosis (LAA) strokes were significantly downregulated than those in non-LAA strokes and controls. These results suggested that ZFAS1 might be used as a potential noninvasive biomarker for the diagnosis of LAA stroke [154].
SETD5-AS1 is another lncRNA, which is mainly regulated stroke development and its underlying mechanism. SETD5-AS1 was highly expressed in the ischemia-reperfusion injury model. SETD5-AS1 participates in the development of IS by activating PTEN and inhibiting PI3K/AKT pathway [155]. The functional intergenic repeating RNA element was associated with OGD/R injury, and it could activate the NF-kB signal pathway [156].
There were very few studies carried out to delineate the link between long non-coding RNAs and their role in angiogenesis after stroke. Maternally expressed gene 3 (Meg3) is a type of lncRNA, which has a role in angiogenesis, and it was significantly decreased after IS. Overexpression of Meg3 suppressed functional recovery and decreased capillary density after IS, whereas downregulation of Meg3 ameliorated brain lesion and increased angiogenesis after IS [157]. Another investigator demonstrated that Meg3 functions as a cell death promoter in ischemia and physically and functionally interacts with p53 to mediate ischemic damage [158].
lncRNA-N1LR promoted neuroprotection, probably through the inhibition of p53 phosphorylation on serine 15 in a manner that was independent of its location-associated gene Nck1 [159]. Fos downstream transcript (FosDT) is a lncRNA, and that is cogenic with the Fos gene. During the ischemic condition, the expression level of FosDT and Fos was induced. Knockdown of FosDT significantly ameliorated the post-ischemic motor deficits and reduced the infarct volume, and it also activated the REST-downstream genes GRIA2, NFκB2, and GRIN1 in the post-ischemic brain [160].
In summary, these studies are conducted to profile lncRNA regulatory transcripts in ischemic conditions using different technologies. The functional importance and molecular regulatory mechanisms of lncRNAs and links between lncRNA-mRNA in IS conditions are unknown. The identification of new lncRNA transcripts will provide a novel opening to study RNA directed epigenetic regulators in IS condition, as well as likely reveal novel targets for a promising translational future of lncRNA-based diagnostics and therapeutics in IS (Fig. 2).
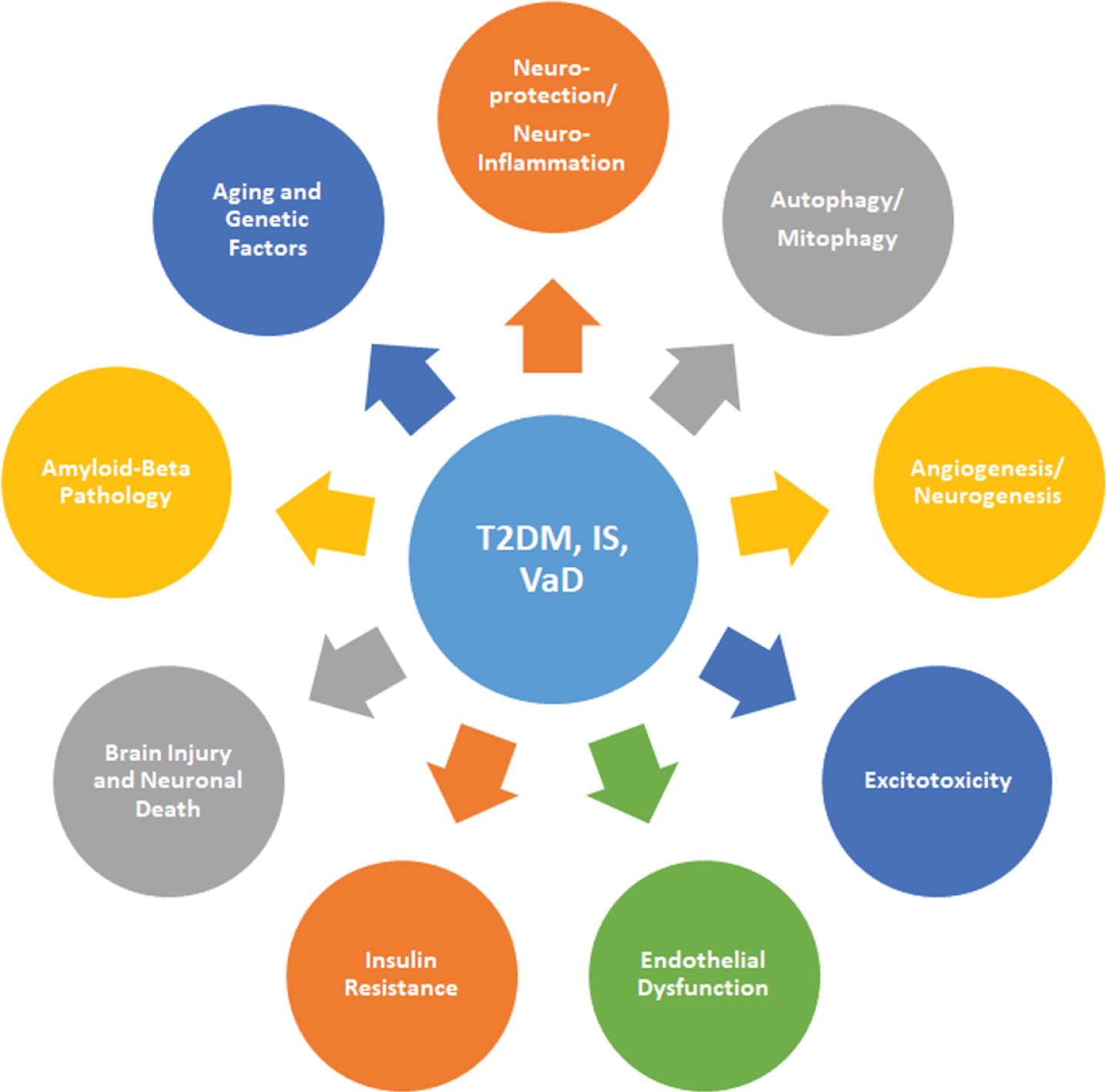
A pictorial diagram illustrating multiple pathological mechanisms linked to type 2 diabetes (T2DM), ischemic stroke (IS), and vascular dementia (VaD). Insulin resistance, excitotoxicity, endothelial dysfunction, amyloid-β pathology, brain injury, neuroinflammation, neuronal death, autophagy, and mitophagy, resulting in the pathogenesis of T2DM, IS, and VaD.
Role of long non-coding RNAs in vascular dementia
An in-depth analysis of lncRNAs can provide comprehensive, complete, and specific information relating to a given tissue at a specific time, which brings many advantages, including the discovery of novel genomic sequences and the accurate quantification of expression levels. These advantages allow us to understand the cellular and molecular mechanisms and therefore make it possible to identify innovative therapeutic targets [161]. However, only one study was conducted so far to delineate the link between lncRNAs and VaD. Li et al. (2019) [162] studied the comprehensive evaluation of white matter damage and neuron death and whole-transcriptome analysis of rats with CCH. CCH results in the development and progression of cognitive impairment, such as VaD. The whole-transcriptome microarray was performed to assess mRNA, circRNA, and lncRNA expression profiles at 4th and 8th weeks after CCH. The microarray analysis identified 491 lncRNAs were shown to be differentially expressed, 196 lncRNAs were upregulated, and 295 lncRNAs were downregulated. lncOlr1718 was the most upregulated lncRNA, and lncStyxl1 was the most downregulated lncRNA [162].
Unexpectedly, the level of research on lncRNA association with VaD is still in a vacuum. More research is urgently needed to define the link between lncRNAs and VaD.
INTERACTION AND CROSSTALK BETWEEN miRNAs AND lncRNAs IN T2DM AND IS
Non-coding RNAs are functional RNA molecules that are not translated into proteins but can regulate the expression and function of many protein-coding genes [69]. In this part of the review, we have shown that the T2DM and IS condition intensely affects the expression profile of ncRNAs such as miRNA and lncRNA. lncRNAs may also perform functions through competitively binding to miRNAs known as competitive endogenous RNAs (ceRNAs) that regulate specific RNA transcripts through competing for shared miRNAs [163]. lncRNA–miRNA interaction and crosstalk might be a prominent mechanism controlling T2DM and IS events.
miR-214 has been demonstrated to suppress gluconeogenesis by targeting activating transcription factor 4 (ATF4), which regulates gluconeogenesis by affecting the transcriptional activity of forkhead box protein O1 (FoxO1). Zhu and the research team aimed to explore whether miR-214 and ATF4 were involved in the lncRNA MEG3-mediated increase of FoxO1 expression [164]. The study stated that MEG3 promoted hepatic IR by serving as a ceRNA of miR-214 to facilitate ATF4 expression. Further, these data provide insight into the molecular mechanism of MEG3 involvement in the development of T2DM [164].
Platelet hyper aggregation and hyper coagulation are associated with an increase of thrombogenic risk, especially in patients with T2DM. lncRNA metallothionein 1 pseudogene 3 (MT1P3) was significantly upregulated in megakaryocytes from T2D patients than in healthy controls. MT1P3 as a key regulator in platelet activation by increasing p2y12 expression through sponging miR-126 under T2DM condition. These findings may provide a new insight for managing platelet hyperactivity-related diseases [165].
lncRNA Gm10768 sequestrated miR-214 to relieve its suppression on ATF4, a positive regulator of hepatic gluconeogenesis. In other words, Gm10768 as a new lncRNA activating hepatic gluconeogenesis through antagonizing miR-214 in mice [166]. LINC01619 functions as a competing endogenous RNA and regulates miR-27a/FOXO1-mediated ER stress and podocyte injury in DN [167]. An imprinted MEG3-DLK1 miRNA cluster is downregulated in human T2DM islets [168].
lncRNA AK038897 was reported to be upregulated in the ischemic condition in the rat. Wei et al. (2019) [169] detected increased AK038897 and decreased miR-26a-5p levels in mouse brains following MCAO/R and in mouse neuroblastoma cells following OGD/R. They identified AK038897 directly binds to miR-26a-5p and functions as a ceRNA for miR-26a-5p to regulate death-associated protein kinase 1 (DAPK1), which is a central mediator of ischemic neuronal death. Further, they concluded that AK038897/miR-26a-5p/DAPK1 was signaling cascade as a key mechanism controlling cerebral ischemia/reperfusion injury [169].
Many lncRNAs are related to angiogenesis following IS, and SNHG12 was one among them. The role and expression of SNHG12 and miR-150 were determined in OGD-stimulated mouse brain microvascular endothelial (bEnd.3) cells and in MCAO mouse models. SNHG12 expression was elevated, whereas miR-150 reduced in OGD-exposed bEnd.3 cells. Anti-miR-150 strengthened the effect of SNHG12 upregulation on the angiogenesis in bEnd.3 cells. Furthermore, SNHG12 functioned as a ceRNA for miR-150 to regulate vascular endothelial growth factor (VEGF) expression. Additionally, overexpression of SNHG12 improved the recovery of neurological function, reduced infarct volume and miR-150 expression, increased vascular density, and VEGF expression in the infarct border zone of MCAO mice [170]. lncRNAs HIF1A-AS2 by sponging to miR-153-3p might regulate the expression of HIF-1α and its downstream targets, thereby influencing angiogenesis in hypoxia [171].
Another study demonstrated that SNHG12 targets miR-199a to upregulate sirtuin 1 (SIRT1) expression, which attenuates cerebral ischemia/reperfusion injury through AMPK pathway activation [172]. SNHG12 suppresses endothelial cell injury induced by OGD/R by targeting miR-199a [173]. miR-199a was identified and validated to be a direct target of Snhg1, and the function effects of Snhg1 on BMECs survival and angiogenesis depended on miR-199a, which was involved in the regulation of HIF and VEGF expression [174]. SNHG14 increased the expression of Phospholipase A2 Group 4A by inhibition of miR-145-5p, which resulted in the activation of microglial cells in cerebral infarction [175].
As we know, lncRNA meg3 has an important role in the IS event, and it was acting as a ceRNA for miR-181b to regulate ischemic brain injury. To evaluate the association of SNPs in MEG3 and miR-181b rs322931 with IS risk, Han and research group extracted genomic DNA from blood samples of 509 patients with IS and 668 healthy controls. These findings indicated that miR-181b rs322931 might singly or jointly contribute to the risk of IS [176]. MEG3 functions as a ceRNAs and competes with programmed cell death 4 mRNA for directly binding to miR-21, which mediates ischemic neuronal death. The knockdown of MEG3 protects against ischemic damage and improves overall neurological functions in vivo [177].
lncRNA growth arrest-specific 5 (GAS5) was known to regulate cell survival negatively and plays a key role in the pathogenesis of several neurological diseases. However, the function and molecular mechanism of lncRNA GAS5 in IS have not been reported. Chen et al. (2018) [178] attempted to detect GAS5 and miR-137 expression in MCAO and OGD-treated models. Furthermore, they also found that GAS5 functioned as a ceRNA for miR-137 to regulate the de-repression of its endogenous target Notch1 and decrease neuron survival through inactivation of the Notch1 signaling pathway [178].
lncRNA Metastasis Associated Lung Adenocarcinoma Transcript 1 (Malat1) is one of the most highly upregulated OGD/R-responsive endothelial lncRNA and plays a protective role in BMECs against cerebral ischemic insults. Malat1 exerted regulatory effects and found that Malat1 served as an endogenous sponge to downregulate miR-26b expression by binding directly to miR-26b. Further, Malat1 overturned the inhibitory effect of miR-26b, which targets Unc-51 like autophagy activating kinase 2 (ULK2) [179].
Inhibitory functions of miR-145 on angiogenesis through direct targeting on VEGF-A and ANGPT2 and proved the protective role of lncRNA-MALAT1 for BMECs under OGD conditions through the direct regulation of miR-145 [180]. Another study revealed that downregulation of MALAT1 attenuated neuronal cell death through suppressing Beclin1-dependent autophagy by regulating miR-30a expression in cerebral IS [181].
lncRNAs TUG1 is known for its critical roles in multiple biologic processes. However, the lncRNA TUG1 expression and function in IS have not been reported yet. In order to know the link between TUG1 and IS, Chen et al. (2017) [182] conducted a study. They found that lncRNA TUG1 expression was significantly upregulated in brain ischemic penumbra from the rat MCAO model, while similar results were also observed in cultured neurons under OGD insult. The knockdown of TUG1 decreased the ratio of apoptotic cells and promoted cell survival in vitro, which may be regulated by the elevated miR-9 expression and decreased Bcl2l11 protein. Furthermore, TUG1 could directly interact with miR-9, and downregulating miR-9 could efficiently reverse the function of TUG1 on the Bcl2l11 expression [182].
Another study stated that the lncRNA Oprm1/miR-155/GATA3 axis plays an important role in cerebral I/R injury. lncRNA Oprm1 may attenuate cerebral injury through the NF-κB pathway [183]. New molecular mechanism of lncRNA CHRF through targeting miR-126/SOX6 signaling to modulate ischemic neuronal injury [184].
H19-miR-19a-Id2 axis plays a critical role in hypoxia induced neuronal apoptosis, and blocking this axis may serve as a novel therapeutic strategy for ischemic brain injury [185]. The HOTTIP overexpression attenuated OGD-induced neuronal injury and imbalanced glycolytic metabolism by sponging miR-143, resulting in the de-repression of its endogenous target HK-2 [186].
In recent years, research studies have highlighted the role of lncRNAs functioning as ceRNAs in the pathogenesis of T2DM and IS. Overall, the findings from the above studies indicated that lncRNAs, miRNAs, and mRNAs are potential targets for the treatment of T2DM and IS (Fig. 3).
Schematic diagram of microRNAs and lncRNAs role in type 2 diabetes, ischemic stroke, and vascular dementia. microRNAs and lncRNAs are involved in pathogenesis and are considered as biomarkers and therapeutic targets.
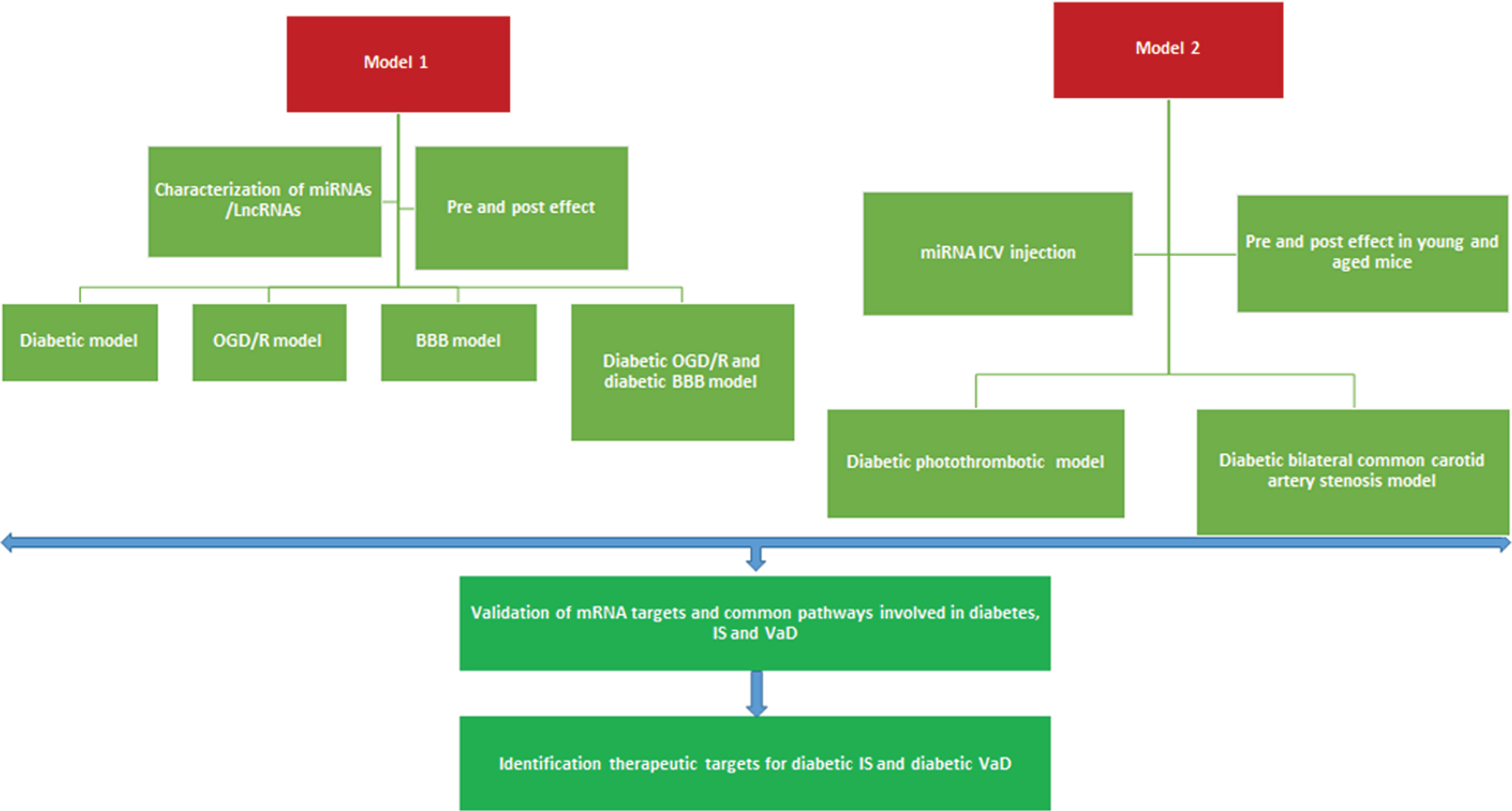
Hypothetical model to study the link between type 2 diabetes, ischemic stroke, and vascular dementia. Model 1) Characterizing the miRNAs and lncRNAs in diabetic, ischemic, and vascular dementia model in an in vitro condition. Model 2) In vivo characterization of miRNAs and lncRNAs using diabetic stroke and diabetic vascular dementia animal models. BBB, blood-brain barrier; OGD, oxygen-glucose deprivation/reoxygenation; ICV, intracerebroventricular.
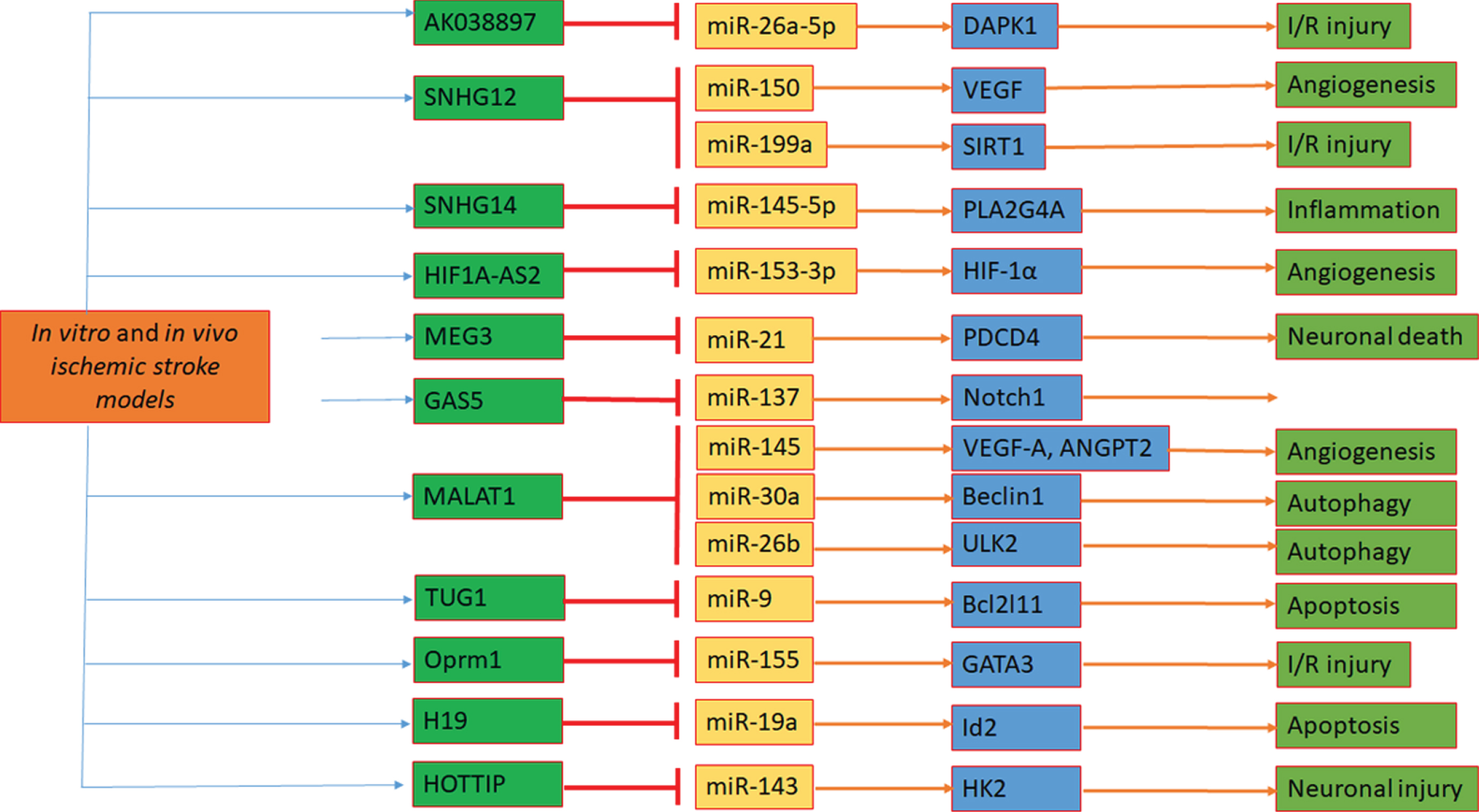
Schematic of functional crosstalk between lncRNAs and miRNAs in ischemic stroke. Ischemia-mediated alterations in lncRNAs, miRNAs, and mRNAs have key roles in angiogenesis, autophagy, apoptosis, neuronal injury, cell death, and survival. I/R, ischemic/reperfusion.
ROLE OF AGING IN T2DM, IS, AND VaD
Aging is defined by the loss of functional reserve over time, leading to decreased tissue homeostasis and increased age-related pathology [187]. Aging takes place in a cell, an organ, or the total organism with the passage of time. It is a process that goes on over the entire adult life span of any living thing. Aging is associated with changes in dynamic biological, physiological, environmental, psychological, behavioral, and social processes. Aging also declines in function of the senses and activities of daily life and increased susceptibility to and frequency of disease, frailty, or disability. In fact, advancing age is the major risk factor for a number of chronic diseases such as T2DM, IS, VaD, and neurological disorders in humans [188].
Aging is the impact of time on the human body, and it occurs on multiple levels: 1) cellular aging, 2) hormonal aging, 3) accumulated damage, and 4) metabolic aging. A healthy diet, regular exercise, regular health check-ups, socialization, and stress-free living are some of the useful suggestions for extending life. By 2030, the number of Americans age 65 and over is projected to reach some 71.5 million people, and nearly 10 million of them will be age 85 and older (U.S. Census Bureau, Population Projections) [188].
Summary of key functions of the microRNAs, long noncoding RNAs, and the interaction of lncRNAs and miRNAs
The prevalence of T2DM is expected to increase gradually with the prolongation of population aging and life expectancy [189]. An increase in the aging of the world’s population is a major contributor to the diabetes epidemic, and older adults represent one of the fastest growing sections of the diabetes population [190]. Older adults are at higher risk for developing T2DM because of the combined effects of increased IR and pancreatic β-cell dysfunction. Aged patients with a family history of diabetes are more likely to develop the disease as they age [191–194]. Carbohydrate metabolism, lifestyle factors, obese, and testosterone levels are the key factors that contribute to the high prevalence of diabetes in the aged population [195]. Aged populations with T2DM are at risk of developing a similar spectrum of microvascular complications such as retinopathy, nephropathy, and neuropathy. Older adults with diabetes suffer excess morbidity and mortality compared with older individuals without diabetes [196]. In addition, they are at high risk for polypharmacy, functional disabilities, and common geriatric syndromes that include cognitive impairment, depression, urinary incontinence, falls, and persistent pain [197].
Oh and the research team developed a new mouse model called JYD, which develops T2DM when they grow old [198]. JYD mice show characteristics of IR, hyperglycemia, and hyperinsulinemia in old age without obesity. They hypothesized that the mechanism of age-dependent T2DM in this model relates to caveolin-1 status in skeletal muscle, which appears to regulate insulin sensitivity in the mice. Upregulation of caveolin-1 in aged diabetic JYD mice significantly improved insulin sensitivity with a concomitant increase of glucose uptake in the skeletal muscle [198]. A research study hypothesized that an age-associated decline in mitochondrial function contributes to IR in the older population [199].
The Honolulu-Asia aging study evaluated the association of diabetes alone or combined with the apolipoprotein E (APOE) gene with incident dementia and neuropathological outcomes in a population-based cohort of 2,574 Japanese-American men, including 216 subjects who underwent an autopsy [200]. They have reported that T2DM was a risk factor for VaD. The association between diabetes and dementia was particularly strong among carriers of the APOE ɛ4 allele. The neuropathological data were consistent with the clinical results [200].
The Massachusetts Male Aging Study examined the association between low testosterone and sex hormone-binding globulin (SHBG) levels and the subsequent development of T2DM in men [201]. Analyses were conducted on the cohort of the Massachusetts Male Aging Study, a population-based random sample of men aged 40-70. The study suggested that low levels of testosterone and SHBG play some role in the development of IR and subsequent T2DM [201].
Since older adults with diabetes have a two- to threefold increased risk of physical disability, another population-based study attempted to study muscle strength and quality in older adults with T2DM [202]. They have concluded that diabetes is associated with lower skeletal muscle strength and quality and contribute to the development of physical disability in older adults with diabetes [202].
In 2006 the Honolulu-Asia aging study investigated the association between T2DM and markers of brain aging on magnetic resonance images, including infarcts, lacunes, and white matter hyperintensities as markers of vascular damage and general and hippocampal atrophy as markers of neurodegeneration in Japanese-American men [203]. They found that older individuals with T2DM have an elevated risk for vascular brain damage and neurodegenerative changes. These pathological changes may be the anatomical basis for an increased risk of cognitive impairment or dementia in T2DM [203].
The aging process causes structural and functional impairments to the neurovascular unit (NVU). The age-related vulnerability of the NVU increases the risk and exacerbates the severity of IS [204]. Aging alters the immunological response to IS. The peripheral immune system plays a critical role in aging and in response to brain injury. Ritzel and the research team demonstrated the differences in the composition of circulating and infiltrating leukocytes recruited to the ischemic brain of old male mice after stroke compared to young male mice [205]. The results suggested that blood neutrophilia and neutrophil invasion into the brain were increased in aged animals. Relative to infiltrating monocyte populations, brain-invading neutrophils had reduced phagocytic potential and produced higher levels of reactive oxygen species, and extracellular matrix-degrading enzymes were further exacerbated with age [205].
A population-based study attempted to compare the prevalence of classic vascular risk factors and polymorphisms of the G20210A coagulation factor II (prothrombin), Arg506Glu coagulation factor V Leiden, C677T methylenetetrahydrofolate reductase (MTHFR), and 4G/5G plasminogen activator inhibitor-1 (PAI-1) and the impact of these gene mutations and classic vascular risk factors on the overall stroke risk in individuals aged 55 years or younger [206]. They found that gene mutations of the procoagulable and proatherosclerotic factors investigated exerted a synergistic action in the development of overall risk of IS in young and middle-aged individuals [206].
Soares et al. (2010) evaluated the outcome of patients aged 70 years or older with a first-ever acute IS and to identify the factors which determine the poor outcome. The study included that data from 115 patients, non-disabled prior to the stroke, consecutively admitted to a medical department of a teaching hospital over a 30-month period, were prospectively collected at stroke onset and 6-month follow-up. They have also recorded the clinical and brain imaging findings and functional status. The outcome of the study was near 50% of these old patients were dependent or dead 6 months after stroke onset, and the main predictor of poor outcome was the neurological severity of stroke [207]. Data from the first dementia study that was representative of the United States population suggest that stroke, the APOE ɛ4 allele, and their interaction were strongly associated with dementia [208].
MacKnight et al. (2002) conducted a longitudinal study to investigate the relationship between diabetes and incident dementia. The study included 5,574 subjects without cognitive impairment. The study concluded that T2DM at baseline was associated with incident vascular cognitive impairment and its subtypes, VaD [209]. Another meta-analysis data investigated the association between diabetes and the risk of all types of dementia (ATD), AD, and VaD. The results showed a 73% increased risk of ATD, a 56% increase of AD, and a 127% increase of VaD in diabetes patients [210].
Human aging is characterized by 1) an exponential increase in the death rate due to a larger number of health issues and diseases, 2) decrease in efficiency of homeostatic mechanisms, and 3) decrease in chemical and physical changes in cell and molecular levels. Overall, the above-mentioned studies clearly stated that how aging accelerates T2DM, IS, and VaD disease processes.
NON-CODING RNAs AS POSSIBLE THERAPEUTIC TARGETS AND GENETIC THERAPIES FOR T2DM, IS, AND VaD
An increasing number of studies have shown that miRNAs and their clusters may have clinical significance as biomarkers and therapeutic targets for the number of diseases. Non-coding RNAs can be used to indicate a disease stage and pathology, genetic linkages, and disease progressions [211]. In the case of miRNA, one or a combination of miRNAs are needed to assess for an accurate diagnosis. Currently, these investigations have been carried out in different preclinical animal models, as well as in human studies [212]. The possible outcome from these preclinical trials is that a combination of miRNA biomarkers can be used as potential therapeutic drugs. To date, there are numerous clinical trials have been completed [212] and important message from these trials is that miRNAs can have several advantages, including miRNAs ubiquitously expressed throughout the body, they can be easily measured from peripheral fluids and tissues such as saliva, urine, cerebrospinal fluid, and other biological samples [213, 214]. Further, since these miRNAs can cross the BBB, they can be quantitated in the brain and peripheral fluids for various neurodegenerative diseases.
Biomarker development is still in the early stages for several neurological diseases. In the medical field, non-coding RNAs seem to provide various potential applications in diseases such as diabetes and its associated complications. However, there are several drawbacks observed from the currently available biomarker studies. These are small sample sizes (cohorts) and there is inconsistency of miRNA assessment in human samples. Therefore, it is important to standardize the miRNA detection methods and increase the sample sizes (a large number of patients and age/sex-matched control subjects) to assess miRNA levels. With these measures, miRNA data can be incorporated into medical practice, and miRNAs appear to be bona fide biomarkers for diabetes in the future [215].
As we stated earlier, non-coding RNAs contributed to the pathophysiological process of many diseases, and IS is not a single disease but a syndrome. More importantly, the changes of non-coding RNAs happened early or immediately after IS, and these changes would affect the local neuronal apoptosis, inflammation, and brain edema via changing the expressions of target genes [216]. In some cases, non-coding RNAs were upregulated, whereas in others downregulated. Many known biomarkers related to IS are not disease specific and have also been associated with other types of brain injuries. Therefore, if restoring the expressions of non-coding RNAs, would help to treat IS and advance the prognosis of stroke patients [217].
A considerable amount of research effort needs to be invested to determine the non-coding RNA(s) profiles in routine medical practice, and their potential benefits in early diagnosis of T2DM, IS, and VaD. Further, more research on genetic therapies and non-coding RNA is urgently needed for T2DM, IS, and VaD.
CONCLUSIONS AND FUTURE DIRECTIONS
The main purpose of this article is to summarize the current state of knowledge regarding miRNAs, lncRNAs, and their roles in the T2DM, IS, and VaD conditions. The miRNAs and lncRNAs have established in the past decades and are emerging as important biomarkers in the regulation of biological processes. Further, recent studies also revealed that differentially expressed miRNAs and lncRNAs are potential therapeutic targets and indicators of biological pathways among T2DM, IS, and VaD conditions. Recent studies from our laboratory and others have demonstrated that non-coding RNAs and their gene transcripts can be considered as therapeutic targets for IS. The impact of cellular and molecular mechanisms between T2DM and IS, T2DM, and VaD remains unknown. There are many questions raised in research linking T2DM, IS, and VaD, which are largely unanswered. The identification of the lncRNA-miRNA-mRNA target is a critical stage to the understanding of non-coding RNAs-mediated gene expression in T2DM, IS, and VaD. Future research is still needed to understand the molecular basis and therapeutic potential of miRNAs and lncRNAs for T2DM, IS, and VaD. It is important to assess non-coding RNAs using a larger number of clinical samples (postmortem brains, fibroblasts, B-Lymphoblasts, primary neurons, and neurons/cells transfected with miRs and lnc-miRs) in disease states such as T2DM, IS, and VaD patients.