
Abstract
Background:
Given that amyloid-β (Aβ) peptide is produced and released at synapses, synaptic Aβ is one of the promising therapeutic targets to prevent synaptic dysfunction in Alzheimer’s disease (AD). Although Aβ production begins with the cleavage of the amyloid-β protein precursor (AβPP) by β-site AβPP cleaving enzyme 1 (BACE1), the mechanism on how BACE1 is involved in AβPP processing at synapses remains unclear.
Objective:
This study aimed to identify novel BACE1 interacting proteins regulating Aβ production at the synapse.
Methods:
BACE1 interacting proteins were pulled down using a mass spectrometry-based proteomics of wild-type (WT) rat brain synaptoneurosome lysates utilizing anti-BACE1 antibody. Then, a novel BACE1 interactor was identified and characterized using experimental systems that utilized transfected cells and knockout (KO) mice.
Results:
Synaptic vesicle protein 2B (SV2B) was identified as a novel presynaptic interaction partner of BACE1. In HEK293 cells, co-overexpression of SV2B with BACE1 significantly reduced the sAβPPβ and Aβ levels released in the media; thus, SV2B overexpression negatively affected the AβPP cleavage by BACE1. Compared with those of WT mice, the hippocampal lysates of SV2B knockout mice had significantly elevated Aβ levels, whereas the β-secretase activity and the AβPP and BACE1 protein levels remained unchanged. Finally, a fractionation assay revealed that BACE1 was mislocalized in SV2B KO mice; hence, SV2B may be involved in BACE1 trafficking downregulating the amyloidogenic pathway of AβPP.
Conclusion:
SV2B has a novel role of negatively regulating the amyloidogenic processing of AβPP at the presynapses.
INTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative disorder characterized by the presence of senile plaques, synaptic loss, neurofibrillary tangles (NFTs), and neuronal cell death [1]. It is the leading cause of dementia. Considering that the main components of senile plaques and NFTs are amyloid-β protein (Aβ) and hyperphosphorylated tau, respectively, they have been long considered as the promising targets for AD treatment. Importantly, synaptic loss most closely correlates with the memory impairment, which is the pathological hallmark of AD [2, 3]. Synapses have been extensively regarded as the primary and upstream targets in AD. In addition, synaptic dysfunction can be induced by soluble Aβ oligomers before the onset of cognitive decline [4]. Further, a recent study on synaptic pathology in the early pathological stages of AD demonstrated that synaptic Aβ oligomers rather than NFTs are closely related to the onset of cognitive decline [5].
Aβ is produced by sequential cleavage of the amyloid-β protein precursor (AβPP) at differential sites by the β-site AβPP cleaving enzyme 1 (BACE1) and the γ-site AβPP cleaving enzyme (γ-secretase). AβPP cleavage by BACE1 is the first step and is responsible for the rate-limiting step to produce Aβ [6]. The activity and expression level of BACE1, which is a major neuronal β-secretase, were significantly increased in mild cognitive impairment brains as well as AD brains [7–9]. Interestingly, an AβPP rare missense variant (A673T) that is reportedly protective against late-onset AD makes AβPP a less-favorable substrate for β-secretase, resulting in the reduction of Aβ levels by 40% compared to wild-type (WT) AβPP in transfected HEK293 cells [10]. Hence, aberrant amyloidogenic processing of AβPP by BACE1 may play an important role at an earlier stage of the disease.
Recently, BACE1 has been found to be preferentially localized at presynaptic terminals including synaptic vesicles (SV) [11–15]. Although BACE1 interacts with a growing number of proteins that could modulate β-secretase activity [16–18], the synaptic binding partners of BACE1 have remained poorly understood. In the current study, we focused on and searched for potential synaptic proteins interacting with BACE1 that can modulate β-secretase activity at synapses.
Methods
Plasmids, cell cultures, and transfection
BACE1-FLAG plasmid was purchased from Sino Biological, Inc. Human plasmids of WT synaptic vesicle protein 2A (SV2A) and WT synaptic vesicle protein 2B (SV2B) were purchased from ORIGENE and R&D, respectively. SV2A and SV2B were cloned into pcDNA 3.1 V5 His Topo vector (Invitrogen, Waltham, MA, USA). The empty pcDNA3.1 V5 His Topo vector was used as a control in experiments overexpressing either SV2A or SV2B plasmid. GFP was cloned into pcDNA3.1/CT-GFP-TOPO (Invitrogen). Human Embryonic Kidney 293 (HEK293) cells were cultured in DMEM (Invitrogen) supplemented with 5% FBS (Life Technologies, Waltham, MA, USA) and 1% penicillin/streptomycin (Invitrogen) in a 37°C and 5% CO2 incubator. HEK293 cells were transiently transfected with the abovementioned expression plasmids by using Transfectin (Thermo Fisher Scientific, Waltham, MA, USA).
Synaptoneurosome (SNS) extraction
SNSs were isolated as described previously [19, 20] with minor modifications. In brief, cortices or hippocampi from an 8-week-old male Wistar rat were mechanically homogenized in 2 ml ice cold buffer A (25 mM HEPES, 120 mM NaCl, 5 mM KCl, and 1 mM MgCl2) supplemented with 2 mM dithiothreitol (DTT) and HALT protease and phosphatase inhibitor cocktail (Thermo Fisher Scientific). To remove cellular debris, we filtered the samples through two layers of 80μm nylon filters (Millipore, Temecula, CA, USA). Next, we filtered the samples through a 5μm Supor membrane filter (Pall Corp., Port Washington, NY, USA) to remove large organelles and nuclei, and then centrifuged the filtrates at 1000×g for 10 min to pellet SNSs. The pellets were lysed in 1% CHAPSO buffer (50 mM Tris, 100 mM NaCl, pH 7.4 and 1% CHAPSO) supplemented with 2 mM DTT and HALT protease and phosphatase inhibitor cocktail (Thermo Fisher Scientific). Meanwhile, the soluble supernatant fraction was used for immunoprecipitation and western blot.
Antibodies
The primary antibodies used were as follows: Anti-BACE1 (D10E5, Cell signaling Technology, Danvers, MA, USA), anti-SV2B rabbit polyclonal antiserum (Synaptic Systems, Goettingen, Germany), anti-SV2B mouse monoclonal antibody (Synaptic Systems), anti-SV2A mouse monoclonal antibody (Synaptic Systems), anti-FLAG M2 monoclonal antibody (Sigma Aldrich, St. Louis, MO, USA), anti-V5 monoclonal antibody (MBL, Woburn, MA, USA), anti-myc (9B11, Cell Signaling Technology), anti-amyloid precursor protein (APP) CT (A8717, Sigma-Aldrich), anti-GFP rabbit antibody (Invitrogen), anti-normal rabbit IgG (MBL), and anti-β-actin mouse monoclonal antibody (Sigma Aldrich). For western blot, we used the horseradish peroxidase (HRP)-conjugated antibodies (Jackson Immuno Research, West Grove, PA, USA).
SV2B knockout (KO) mice
The brain samples of SV2B KO mice were kindly provided by Dr. Kawachi and Dr. Fukusumi of Niigata University [21]. In brief, SV2B-mutant mice (129P2/OlaHsd background) [22] were generated from frozen sperm in Jackson Laboratory (Stock No. 006383). SV2B KO mice were obtained by mating SV2B heterozygous mice. Moreover, the age-matched WT mice were also provided as controls.
Mouse brain lysates
Brain cortices and hippocampi were dissected from WT mice and SV2B KO mice aged 4 and 5 months. They were homogenized in a radio-immunoprecipitation assay (RIPA) buffer (50 mM Tris-HCl, 150 mM NaCl, 1% Triton X-100, 1% NP-40, 0.5% deoxycholate, 0.1% SDS, pH 8.0). Then, we incubated brain lysates for 1 h at 4°C and centrifuged them at 14,000×g to collect the soluble fractions, which were subjected to western blot and enzyme-linked immunosorbent assay (ELISA). All experiments that involved the use of a mouse brain tissue were approved by the Subcommittee for Research Animal Care at Kyoto University.
Western blot
Total lysates were diluted with 4×LDS sample buffer (Invitrogen) containing 2% β-mercaptoethanol and then denatured at 95°C for 3 min. Immunoprecipitated or total proteins were separated by SDS-PAGE on 4%–12% Bis-Tris NuPage polyacrylamide gels (Thermo Fisher Scientific). After electrophoresis, the proteins were transferred to PVDF membranes, which were subsequently probed with respective primary and corresponding HRP-conjugated secondary antibodies (Jackson Immuno Research). The blots were visualized using the Western Lightning chemiluminescence reagent (Nacalai tesque, Kyoto, Japan).
Immunoprecipitation
Aliquots of the supernatant from SNS lysed with 1% CHAPSO buffer or HEK293 cell samples lysed with 1% Triton-X buffer containing equal amounts of total protein were incubated with anti-BACE1 and anti-V5 antibodies overnight at 4°C with end-over-end rotation. We used normal rabbit IgG or mouse anti-Flag antibodies as negative controls. After the overnight incubation, each sample was added with 15μl of Protein G Dynabeads (Invitrogen) and then incubated for 10 min at room temperature. Thereafter, the Dynabeads were collected and washed with the same lysis buffer 5 times. Bounded proteins were eluted by boiling at 95°C for 5 min with 2×LDS sample buffer (Life Technologies) containing 2% β-mercaptoethanol.
ELISA for Aβ and sAβPPβ
To determine the extracellular Aβ levels in HEK293 cell culture experiments, we exchanged the conditioned media to fresh serum-free OPTIMEM (Life Technologies) 24 h after transfection and the cells and the media were collected after another 12 h. The cell pellets were suspended and homogenized by pipetting in 1% Triton X-100 buffer (50 mM HEPES, 100 mM NaCl, pH 7.4, and 1% Triton X-100) supplemented with HALT protease and phosphatase inhibitor cocktail and rotated for 1 h at 4°C. The soluble fraction was obtained after centrifugation at 14000 rpm for 15 min. The collected media were also centrifuged to pellet the debris; meanwhile, the supernatant was subjected to ELISA. According to the manufacturer’s instructions, Aβ40, Aβ42, and sAβPPβ levels were measured using the human/rat Aβ40 ELISA kit (Wako, Tokyo, Japan), the human/rat Aβ42 ELISA kit (Wako), and the human sAβPPβ-w (highly sensitive) assay kit (IBL, Fujioka, Japan), respectively. Each value recorded in the ELISA was normalized to the protein concentration of the corresponding cell extract measured with the BCA protein assay kit (Thermo Fisher Scientific).
In vitro β-secretase activity assay
We measured the β-secretase activity by using Beta-secretase activity assay kit (Abcam, England, UK) according to the manufacturer’s instruction. In brief, we homogenized the hippocampal tissues extracted from either WT or SV2B KO mice in an extraction buffer and centrifuged them at 4°C at top speed. We adjusted each supernatant to 1 mg/ml concentration with the extraction buffer and applied 50μl aliquots to a 96-well white microplate (Greiner, Austria). In each well, we added β-secretases substrate, which was designed to produce fluorescent signals when cleaved with β-secretases. After incubation at 37°C for 1 h, we measured the fluorescent output on a fluorescent microplate reader (TECAN, Männedorf, Switzerland) at Ex/Em = 335/495 nm.
Fractionation of mouse brain lysates by discontinuous OptiPrep density gradient
The subcellular fractionation of mice brain lysates was prepared according to the procedures described previously [30] with minor modifications. In brief, brain cortices dissected from either WT mice or SV2B KO mice aged between 3 and 4 months were homogenized by pushing and twisting the pestle (25 strokes) in a cold buffer (25 mM Tris HCl, 130 mM KCL, 25 mM NaCl, 1 mM EGTA, pH 7.4) supplemented with protease inhibitors (Roche, Indianapolis, IN, USA). Then, we centrifuged the homogenates at 1,000×g for 10 min at 4°C and the collected supernatants at 10,000×g for 15 min at 4°C. Each supernatant was loaded on a step OptiPrep gradient of 2.5%–20% with 2.5% increments and centrifuged in SW 41 Ti rotor (Beckman Coulter Life Sciences, Indianapolis, IN) at 126,000×g at 4°C for 1 h. Then, 13 fractions were collected from the top by an aliquot of 1 ml, and equal amounts of the fractions were subjected to SDS-PAGE and western blot.
Liquid chromatography (LC)-mass spectrometry (MS)/MS analysis
For MS analysis, we used rat SNS lysates prepared in lysis buffer containing 1% Triton X-100 (see above) were used for MS analysis. The soluble supernatant fraction was divided into equal aliquots that were incubated with anti-BACE1 antibody as a bait with or without 2 mM CaCl2 . We used anti-HA antibody as a negative control. The bound proteins were eluted with 2×SDS Nupage sample buffer (Life Technologies) and separated by electrophoresis using 4%–12% 1.5 mm Bis-TrisNupage gels (Life Technologies). The gel was stained using a silver staining kit for protein (GE health care, life sciences) according to the manufacturer’s instruction. We excised bands of approximately 70 kDa that displayed differential protein profile between the Ca2+ (+) and Cs2+ (–) conditions and analyzed by the Division for Mass Spectometry of Kyoto University’s Medical Research Support Center (http://support-center.med.kyoto-u.ac.jp/SupportCenter/en/room01).
Statistical analyses
For quantitative western blot analysis, the intensity of respective protein bands was quantified using the National Institute of Health (NIH) Image software. Statics were calculated with Microsoft Office Excel 2007 or GraphPad Prism 8 software using Student’s t-test and one-way ANOVA with multiple comparison Tukey-Kramer’s post-hoc test. Data were shown as means±SEM, and the value p < 0.05 indicated a significant difference.
Results
SV2B is screened as a candidate for a novel binding partner of synaptic BACE1
Synaptic Aβ production and its subsequent secretion are closely linked with a neuronal activity regulated in a Ca2+-dependent manner [23–25]. Thus, to search for potential interaction partners of BACE1 at the synapse, we performed a MS proteomics screening of SNSs from either WT rat cerebral cortex or hippocampus solubilized with 1% Triton X-100 with or without 2 mM Ca2+, with an anti-BACE1 antibody used as a bite. Indeed, synaptic marker proteins were more concentrated in the SNS fraction than that in the total fraction (Fig. 1A, left panel). The efficiency of immunoprecipitation with 2 mM Ca2+ was similar to that of immunoprecipitation without Ca2+ (Fig. 1A, right panel). Next, each sample immunoprecipitated with the anti-BACE1 antibody was run on SDS-PAGE, and then the gels were subsequently silver-stained. Based on the difference in the band intensities between Ca2+ (+) and Ca2+ (–) conditions, the gel slices with more intense bands of approximately 70 kDa in Ca2+ (+) condition were excised from Ca2+ (+) and Ca2+ (−) conditions, respectively (Fig. 1B, arrowheads). The excised gel samples were sent for MS analysis. Synaptic vesicle protein 2B (SV2B) and vesicle-fusing ATPase (N-ethylmaleimide-sensitive factor; NSF) were screened as candidates for novel BACE1-interacting presynaptic proteins that show calcium-dependent profile of the interaction (Table 1). Interestingly, previous human genetic data demonstrated that SV2B gene expression was significantly downregulated in the hippocampal tissues with AD who harbored APOE4 allele(s), which is the strongest genetic risk factor for sporadic AD, or who exhibited progression of cognitive decline [26, 27]. In the present study, we focused on SV2B as a potential BACE1 interactor for regulating the amyloidogenic processing of AβPP at the presynapse.
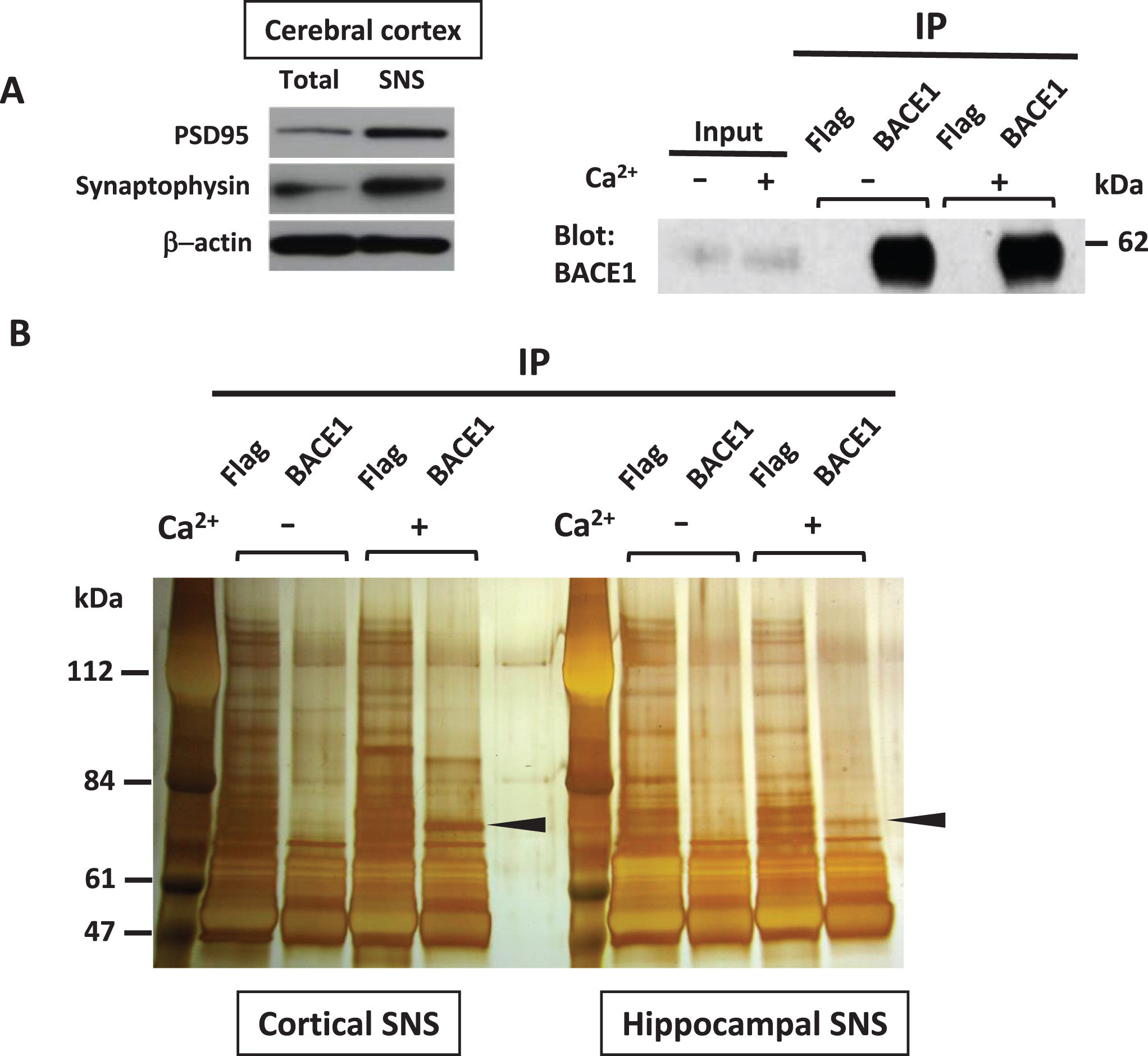
The co-immunoprecipitation assay of synaptoneurosome (SNS) fractions from WT rat cerebral cortices or hippocampi for potential synaptic interactors of BACE1. A) Western blot analysis showed that the synaptic markers, PSD95 and synaptophysin, were concentrated in SNS fractions compared to total homogenates (left). Right: Comparison in the immunoprecipitation efficiency of endogenous BACE1 using anti-BACE1 antibody between Ca2+ (−) and Ca2+ (+) condition. B) Each sample of SNS fractions from WT rat cerebral cortex or hippocampus was immunoprecipitated with anti-BACE1 antibody or anti-Flag antibody in the absence or presence of 2 mM Ca2+. The immunoprecipitates were run on SDS-PAGE and the gels were subsequently silver-stained. For MS analysis, the gel slices with more intense bands in Ca2+ (+) condition were excised from the Ca2+ (+) and Ca2+ (–) conditions, respectively (arrow heads).
Mass spectrometry-based proteomics screening identified SV2B and NSF as candidates of novel BACE1-interacting presynaptic proteins with calcium-dependent profile of the interactions in WT rat synaptoneurosomes
The number of total peptides and the sequence coverage for SV2B and NSF in Ca2+ (–) and Ca2+ (+) are shown.
Endogenous SV2B interacts with BACE1 in intact isolated synaptic terminals
In our MS proteomics screening combined with BACE1 pull-down assay, adding high doses of Ca ions (2 mM CaCl2) in the extracts can induce unexpected protein-protein interaction due to possible improper protein folding, especially in calcium-binding proteins. Therefore, to verify the physiological interaction between BACE1 and SV2B on endogenous levels, we performed co-immunoprecipitation assay (co-IP) combined with western blot using 1% Triton-X 100-solubilized extracts of the SNSs obtained from the hippocampi or cerebral cortices of WT rats. Indeed, in both cortical and hippocampal SNSs, SV2B was co-immunoprecipitated with BACE1 without adding Ca ions, indicating the physiological interaction between both at the synapses (Fig. 2A, B).
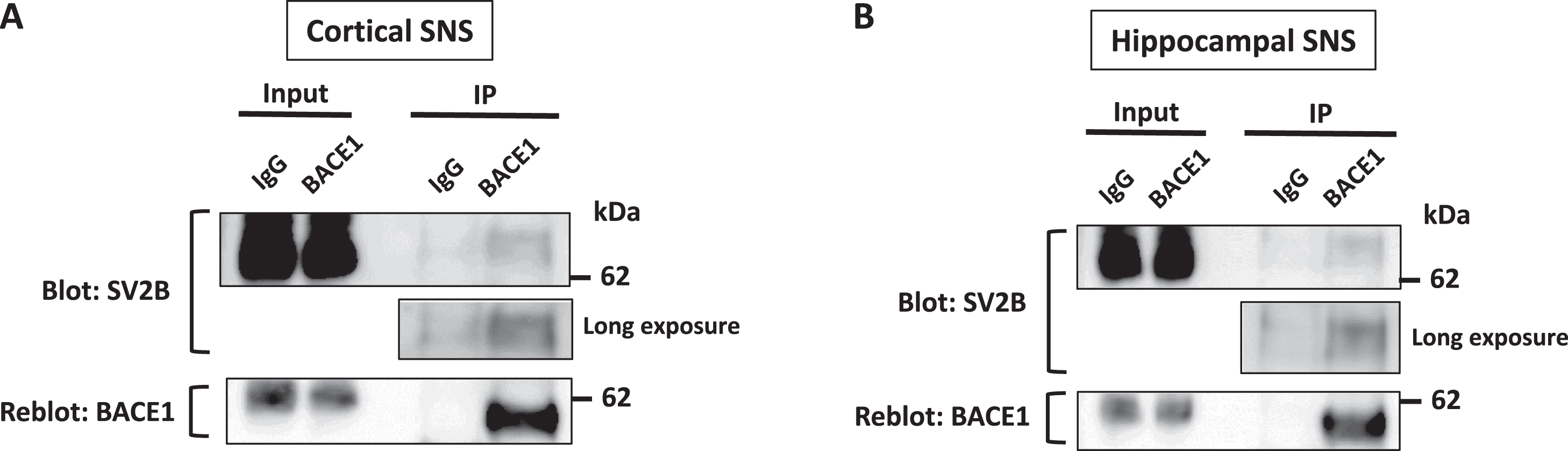
BACE1 interacts with SV2B on endogenous level. SV2B co-immunoprecipitates with BACE1 in SNS fractions isolated from WT rat cerebral cortices (A) or hippocampi (B). SNS lysates were solubilized in 1% CHAPSO buffer. The co-immunoprecipitation assay was conducted using anti-BACE1 antibody for pull-down or normal rabbit IgG as a negative control; the detection antibodies are indicated to the left of each blot, n = 3.
SV2B rather than SV2A preferentially interacts with BACE1 and inhibits the amyloidogenic processing of AβPP in cell culture experiments
To verify the biological significance of SV2B-BACE1 interaction, we sought to clarify the possible effect of SV2B on AβPP processing by BACE1 in vitro. To this end, a plasmid to encode V5-tagged SV2B was transiently cotransfected with FLAG-tagged BACE1 plasmid and GFP plasmid as a marker for transfection efficiency into HEK293 cells by 1 : 1:0.2 ratio. To characterize the potential effect of SV2B as a novel BACE1 interactor, we utilized a plasmid encoding V5-tagged SV2A as an isoform control instead of SV2B-V5 plasmid, in addition to an empty plasmid (pcDNA3.1-V5) as the negative control. The overexpression of either SV2B or SV2A did not significantly affect the expression levels of BACE1 compared with that of the negative control (Fig. 3A, B). Next, we investigated the physical interaction between BACE1 and either SV2B or SV2A in the transfected HEK293 cells through co-IP assay. Equal amounts of cell lysate obtained from each transfected cell were immunoprecipitated using the anti-V5 antibody. The cell lysates (2.5% input), as well as the immunoprecipitates (IP), underwent western blot using the anti-FLAG antibody. As depicted in Fig. 3C, the SV2B-transfected cells (lane 3) had considerably higher BACE1 immunoreactivity containing mature and immature forms than the SV2A-transfected cells (lane 2). Quantitative analysis verified that the SV2B-transfected cells displayed significantly higher BACE1 binding affinity, which is the ratio of BACE1:SV2B or SV2A in the IP, with the 4.2-fold higher BACE1 co-IP with SV2B versus SV2A (Fig. 3D). To investigate the SV2B-BACE1 interaction effect on Aβ production through BACE1-mediated AβPP processing, we measured the levels of sAβPPβ, Aβ40, and Aβ42 that were secreted into conditioned media obtained from each transfected cell by ELISA. The overexpression of SV2B significantly reduced the sAβPPβ, Aβ40, and Aβ42 levels in the conditioned media compared with that of the negative control, whereas those levels in the SV2A-transfected cells were reduced slightly, but not statistically significantly (Fig. 4A). Furthermore, the ratios of Aβ42:Aβ40 were comparable among SV2A and SV2B expressing cells (Fig. 4B); hence, SV2B overexpression did not significantly affect the processing of AβPP C-terminal fragment by γ-secretase to determine Aβ40 or Aβ42 as direct products. Therefore, we conclude that the BACE1-SV2B interaction suppresses the AβPP processing by BACE1, resulting in reduced Aβ production.
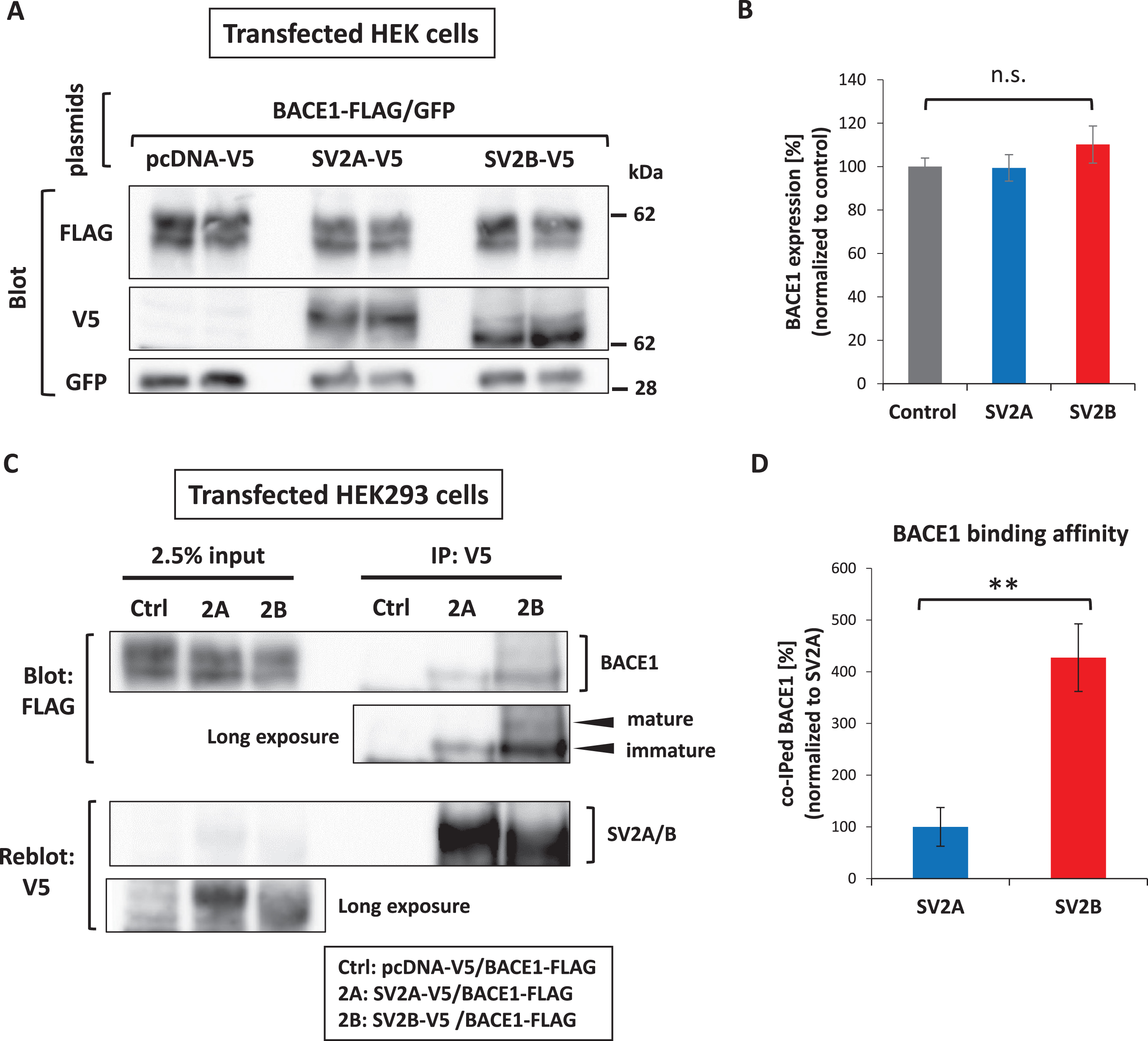
SV2B rather than SV2A preferentially interacts with BACE1 and inhibits the amyloidogenic processing of AβPP. A) A plasmid to encode either SV2B-V5, SV2A-V5, or pcDNA-V5 was transiently cotransfected with BACE1-FLAG plasmid and GFP plasmid as a marker for transfection efficiency into HEK293 cells (each n = 6). Western blot analysis was conducted with the detection antibodies indicated to the left of each blot. B) Quantitative analysis showed no significant differences in the protein expression levels of BACE1 among them. The expression levels of BACE1-FLAG, SV2A-V5, and SV2B-V5 were adjusted by the co-expression level of GFP. C) Western blot analysis of the BACE1-FLAG co-immunoprecipitated with anti-V5 antibody from the cotransfected HEK293 cells lysed in 1% Triton X-100 buffer (n = 6). The immunoprecipitation efficiency was determined by re-probing the membrane with anti-V5 antibody. D) SV2B exhibits a much higher binding affinity to BACE1 than SV2A (n = 6). The quantitative data in all panels are presented as mean±SEM. Statistical significance was determined using the student’s t-test, **p < 0.01, compared with SV2A-expressing cells.
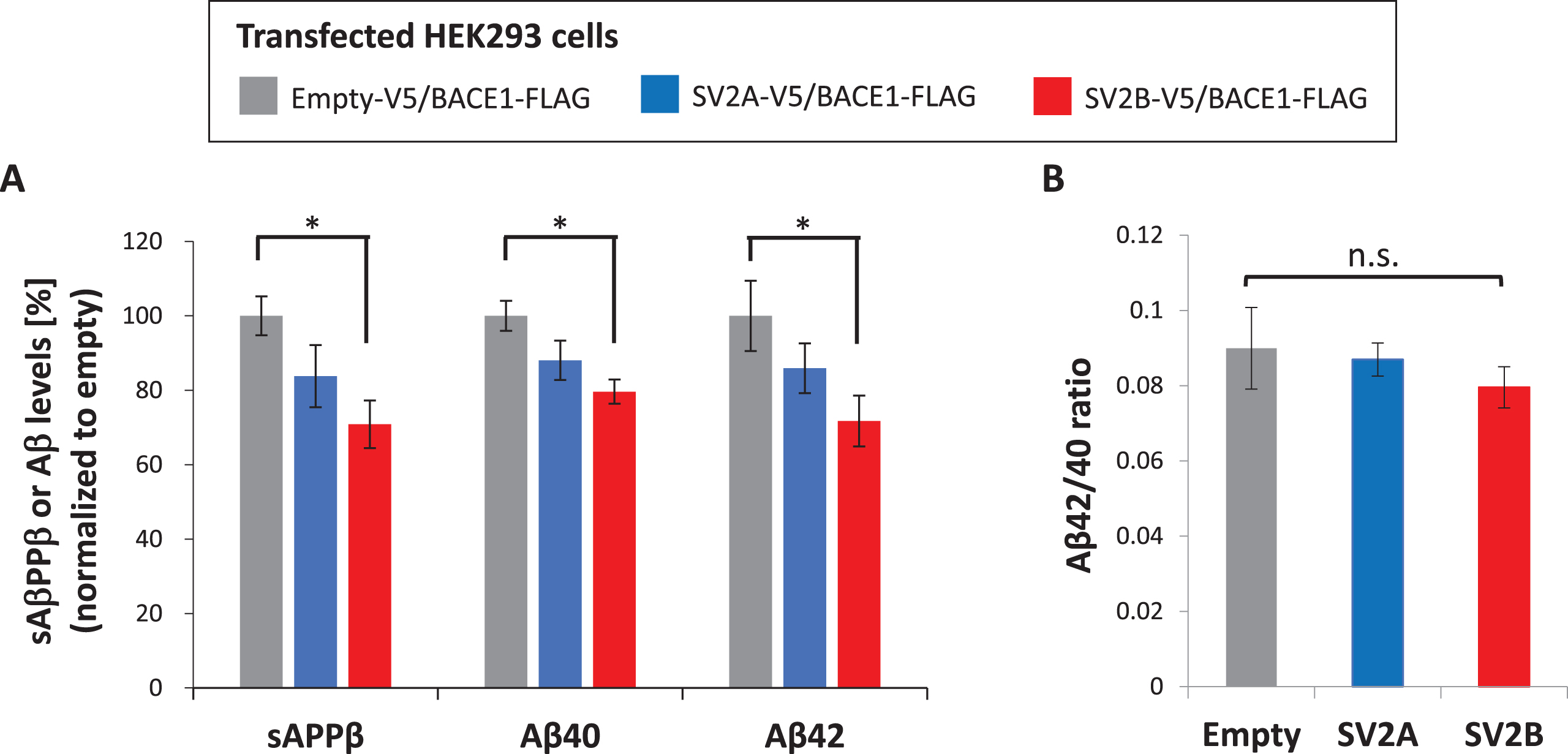
The secreted levels of Aβ and sAβPPβ are inhibited by co-overexpression of SV2B and BACE1 in HEK293 cells. A, B) A plasmid to express either SV2B-V5, SV2A-V5, or Empty-V5 was transiently cotransfected with BACE1-FLAG plasmid into HEK293 cells (each n = 6). The secreted levels of Aβ40, Aβ42, and sAβPPβ measured by the specific ELISA were normalized to the total protein concentration of the corresponding cell extract. The data in all panels of the figure are presented as mean±SEM. Statistical significance was determined using one-way ANOVA with multiple comparison Tukey-Kramer’s post-hoc test, *p < 0.05.
Endogenous mouse Aβ levels are significantly increased in SV2B KO mice in vivo
To further explore the role of SV2B on BACE1-mediated processing of AβPP in vivo, we compared the endogenous protein levels of AβPP and BACE1 and the activity of β-secretase in the hippocampal tissues from WT and SV2B KO mice. Equal amounts of 1% Triton X-100-solubilized lysates underwent western blot analysis to identify AβPP and BACE1 proteins. Quantitative analysis showed no significant differences in AβPP and BACE1 protein levels between WT and SV2B KO mice, indicating that SV2B is unlikely involved in expression of AβPP and BACE1 at the levels of transcription and translation (Fig. 5A, B, left and middle panels). To next examine a possibility that SV2B directly affects the catalytic properties of BACE1 via the interaction, we performed an in vitro β-secretase activity assay using hippocampal preparations from WT and SV2B KO mouse brains. As illustrated in Fig. 5B (right panel), the β-secretase activity assayed in vitro was comparable between WT and SV2B KO mice; hence, the β-secretase proteolytic activity was not directly affected by the absence of SV2B. Meanwhile, the levels of hippocampal Aβ40 and Aβ42 measured by ELISA using equal amounts of 1% Triton X-100-solubilized lysates were modestly but significantly increased in SV2B KO mice compared with those in WT mice, whereas the Aβ42/Aβ40 ratio was comparable between both mouse groups (Fig. 5C). Taken together with the in vitro findings on the novel BACE1-SV2B interaction, these results suggest that SV2B diminishes the amyloidogenic processing of AβPP in vivo, presumably by regulating the accessibility of BACE1 to AβPP rather than affecting the sequential processing by γ-secretase to determine the Aβ42/Aβ40 ratio.
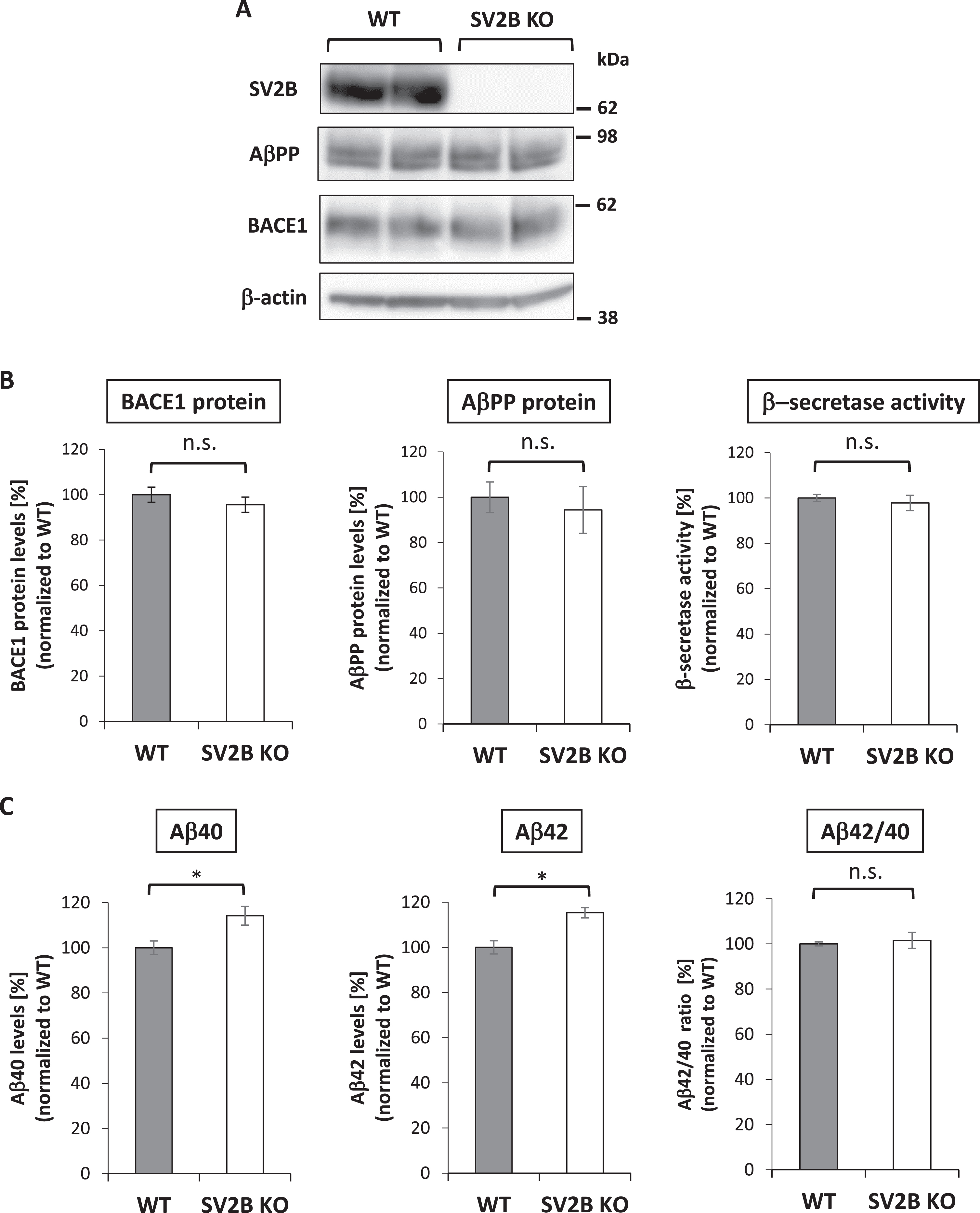
Endogenous mouse Aβ levels in the hippocampi are significantly increased in SV2B KO mice. A) The expression levels of AβPP and BACE1 in the hippocampi from WT or SV2B KO mice were quantified by western blot analysis. Note that no SV2B expression was completely undetectable in SV2B KO mice. B) Quantitative analysis showed no significant differences in AβPP and BACE1 protein levels between both mouse groups (n = 4). The hippocampal β-secretase activity assayed in vitro was comparable between WT and SV2B KO mice (n = 6). C) ELISA measurements of the endogenous Aβ40, Aβ42, and Aβ42/Aβ40 ratio in the hippocampal lysates (n = 6) The quantitative data in all panels are presented as mean±SEM. Statistical significance was determined using the student’s t-test, *p < 0.05.
Loss of SV2B alters BACE1 distribution in the synaptic protein-enriched fractions extracted from the cortical tissues of mouse brains
Considering that SV2B is involved in SV trafficking that leads to neurotransmitter release, we hypothesized that SV2B can negatively regulate the accessibility of BACE1 to AβPP in a trafficking-dependent manner as a key first step in the pathway leading to Aβ production. To determine whether SV2B can influence BACE1 distribution, we compared BACE1 distribution in the homogenates of cerebral cortices from WT and SV2B KO mice through fractionation assay. To exclude the possibility that the lack of SV2B may influence the distribution of synaptic vesicle proteins, we ensured equal amounts of fraction lysate for western blot to detect the major marker for the glutamatergic synaptic vesicle proteins such as synaptotagmin 1 (Syt1) and vesicular glutamate transporter 1 (vGlut1), in addition to SV2B. As expected, SV2B KO mice had no immunoblotting bands specific for SV2B; however, the distribution of SV2B in WT mice was highly concentrated in fraction numbers 3 and 4 (Fig. 6B left, upper panels). Syt1 and vGlut1 in SV2B KO mice were similarly distributed in fraction numbers 3–5 as in WT mice, indicating that the lack of SV2B did not cause altered location of synaptic vesicle proteins in general (Fig. 6B left, middle and lower panels). Interestingly, we found that the distribution of BACE1 in SV2B KO mice showed a statistically significant shift from fraction number 4 to number 5 compared with that in WT mice; meanwhile, AβPP in SV2B KO mice was similarly distributed in fraction numbers 3–5 as in WT mice (Fig. 6B left, C). Taken together, BACE1 can undergo aberrant translocation toward enhanced accessibility to AβPP even without SV2B.
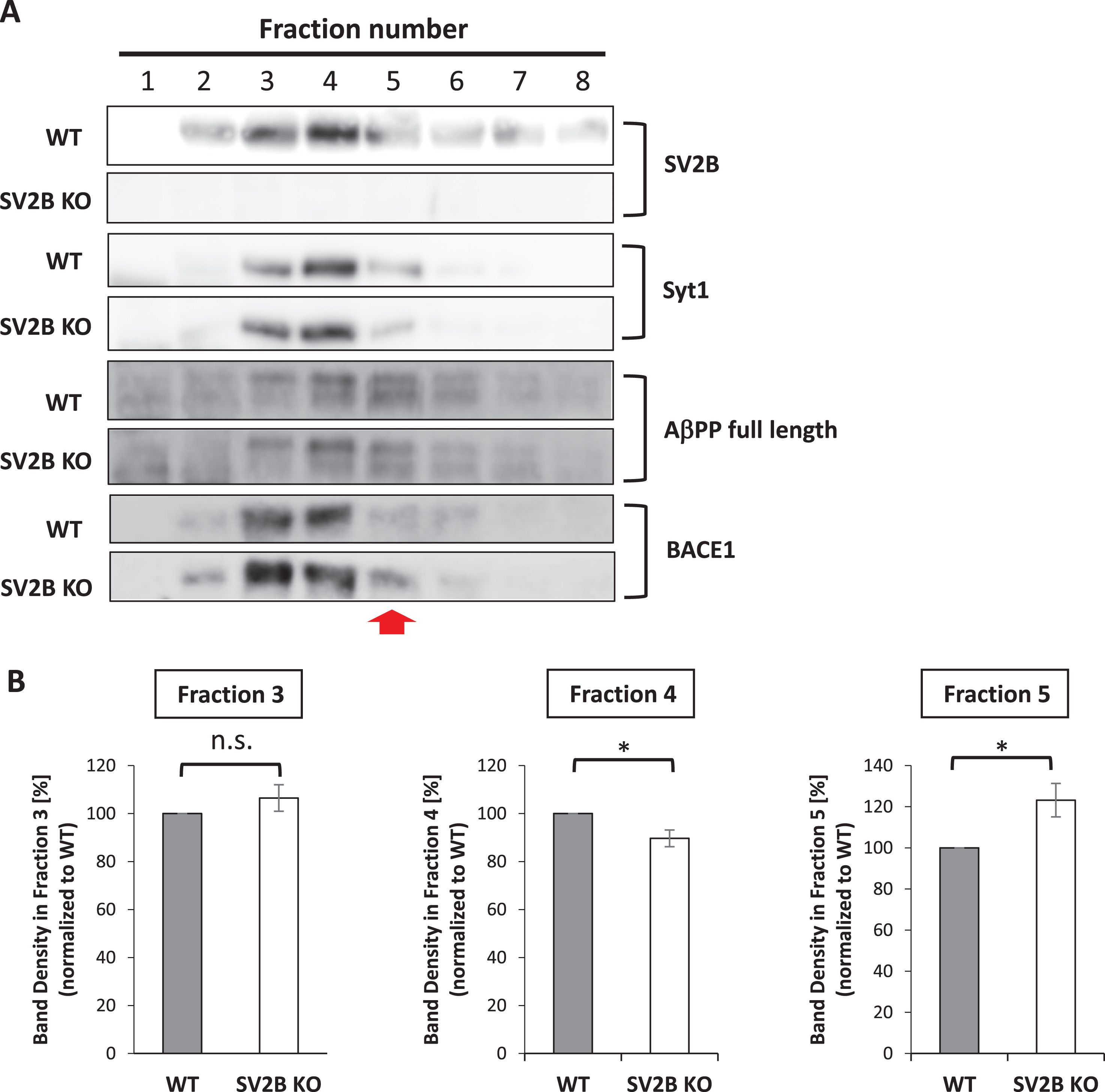
Loss of SV2B alters BACE1 distribution in the synaptic protein-enriched fractions. A) Western blots show distribution of SV2B, Syt1, full length of AβPP, and BACE1 in 8 subcellular fractions of cortical tissues from WT and SV2B KO mice. The red arrow indicates a shift in the subcellular distribution of BACE1 in SV2B KO brains. Note that the subcellular distribution of synaptotagmin1 (Syt1) and full length of AβPP are almost comparable between WT and SV2B KO mouse brains. B) Graphs represent the ratio of the band density of an indicated fraction to the total band density from Fraction 1 to Fraction 8 (n = 5). Data are presented as mean±SEM. Statistical significance was determined using the student’s t-test, *p < 0.05.
DISCUSSION
As generally perceived, sporadic AD is mainly caused by the reduced clearance of Aβ from the brain, whereas familial AD is largely caused by the increased production of total Aβ peptides or the Aβ42 peptide. However, the activity and expression level of BACE1, which is responsible for a major neuronal β-secretase to yield a direct substrate of γ-secretase for Aβ production, were significantly increased in mild cognitive impairment brains as well as AD brains [7–9]. Jonsson et al. [10] reported that the AβPP rare missense variant, A673T, is protective against late-onset AD and age-associated cognitive decline in elderly Icelanders. Importantly, the A673T mutation adjacent to the BACE1 cleavage site makes AβPP a less favorable substrate for β-secretase, resulting in the reduction of Aβ levels by 40% in in vitro study. Thus, Aβ overproduction led by aberrant activation of BACE1 may play an important role at an earlier stage of AD. Meanwhile, Aβ can be produced in a neuronal activity-dependent manner preferably at synapses [23–25]. As previously reported, BACE1 is enriched in presynaptic compartments [11–13], and active BACE1 and PS/γ-secretase complex, as well as AβPP, were found in SV fractions from rat brains by biochemical methods [12, 14]. These physiological findings are particularly important to elucidate the underlying molecular mechanisms in AD pathogenesis, considering that synaptic degeneration is an early pathological hallmark of AD. Intriguingly, synaptotagmin 1 (Syt1), which is the major Ca2+ sensor, interacts with PS1 [20] or AβPP [28], regulating the production and secretion of Aβ (1)]. Further, a very recent report by Maesako et al. [30] demonstrated that synapsin1, which is a key SV protein for neurotransmitter release as well as Syt1, modulates synaptic Aβ production through affecting the AβPP-BACE1 interaction, supporting the link of the amyloidogenic AβPP processing with the neurotransmission machinery. However, the mechanistic research on the molecular mechanisms underlying the BACE1-mediated AβPP processing at the presynapse remains poorly understood. In the current study, SV2B was found to be a novel binding partner of BACE1. Further, SV2B negatively regulates BACE1-mediated Aβ production through the direct interaction of SV2B and BACE1. To our knowledge, this study is the first to show that a particular SV protein involved in neurotransmitter release regulates the amyloidogenic processing of AβPP as a BACE1 interactor.
SV2 is an integral membrane protein with 12 transmembrane domains which is specifically identified in the synaptic vesicles of neurons and endocrine granules [22, 32]. Three SV2 isoforms of SV2A, SV2B, and SV2 C are characterized by different expression levels during brain development and adulthood [33, 34]. Of the SV2 isoforms, SV2B is expressed primarily in glutamatergic synapses in the cortical and hippocampal networks [33], playing a key role in cognitive function, whereas SV2A is ubiquitously expressed in both glutamatergic and GABAergic networks in all brain areas [33]. SV2A binds to levetiracetam, which is an anti-epileptic agent that is widely used to treat seizures in patients with epilepsy. Interestingly, previous studies have demonstrated that levetiracetam could be beneficial for improving cognitive function through suppressing the hippocampal hyperactivation in AD model mice and patients with mild cognitive impairment due to AD [35–37]. Meanwhile, SV2B, not SV2A, is reportedly affected in AD brains. Indeed, SV2B is downregulated in the hippocampi of AD cases with APOE4/4 alleles, which is the strongest genetic risk factor for sporadic AD, in comparison with that of AD cases with APOE3/3. Further, SV2B gene expression is downregulated in the hippocampi and neocortices of patients with AD compared with that in the control [27, 38]. As illustrated in Fig. 7, a potential link between SV2B downregulation and AD may be partially explained by the reduction of inhibitory effect on Aβ production through the interaction of BACE1 with SV2B rather than SV2A found in the current study.
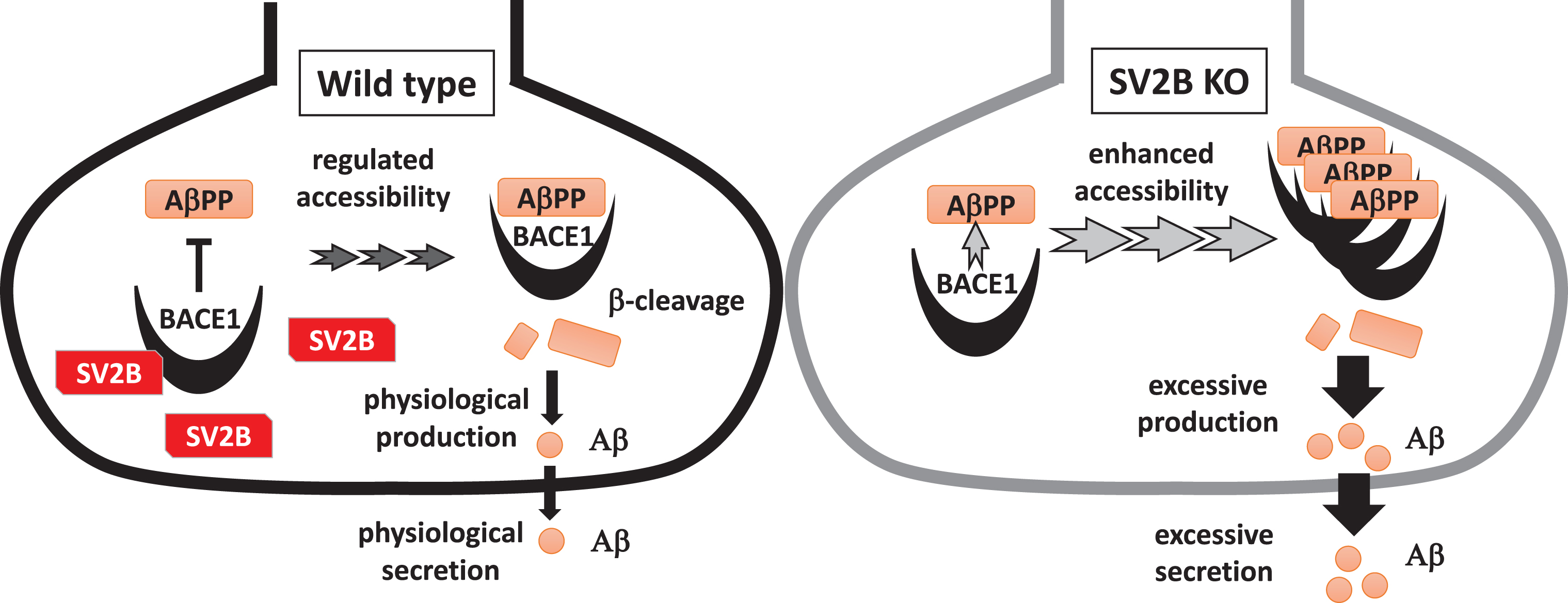
Schematic diagram showing the potential inhibitory effect of the BACE1-SV2B interaction on the amyloidogenic processing of AβPP. In WT presynapses, accessibility of BACE1 to AβPP is tightly regulated under the presence of SV2B. Meanwhile, BACE1 is aberrantly accessible to AβPP in SV2B-null presynapses, resulting in excessive Aβ production.
Overall, SV2B negatively regulates the amyloidogenic processing of AβPP as a novel BACE1 interactor. To elucidate the impact of BACE1-SV2B interaction in the upstream pathogenesis of AD, we plan in our future studies to identify the expression levels of SV2B and the association between BACE1 and SV2B in the brain tissues of patients with AD at an earlier stage.
In conclusion, SV2B is a novel presynaptic interactor of BACE1 regulating the amyloidogenic processing of AβPP to limit Aβ production. However, a deeper understanding of the mechanisms by which the interaction between BACE1 and SV2B negatively regulate the amyloidogenic processing of AβPP is necessary to provide potential new therapeutic strategies for early AD targeting synaptic Aβ.