
Abstract
Background:
Evidence from clinical studies and basic research has shown a strong correlation between Alzheimer’s disease (AD) and type 2 diabetes. Tau, a neuronal microtubule-associated protein, is hyperphosphorylated and aggregated into neurofibrillary tangles in the AD brain. However, the expression of tau in pancreas is under debate.
Objective:
We determined the expression of tau in mouse pancreas.
Methods:
We used western blots, immunoprecipitation, and immunohistochemical staining to analyze pancreatic expression of tau in mice.
Results:
We found that neither total tau nor phosphorylated tau was detectable in the mouse pancreas by western blots. Immunostaining with pan tau antibodies R134d and Tau-5 revealed bright and dense varicosities in the pancreatic islets and the exocrine pancreas. These varicosities were immunoreactive to synapsin 1, a presynaptic marker which can outline autonomic nerve profiles in pancreas, exhibiting complete colocalization with tau. Importantly, endocrine cells in islets did not exhibit specific immunoreactivity to any of pan tau antibodies tested, nor did the exocrine cells.
Conclusion:
In the mouse pancreas, we found that tau is exclusively expressed in autonomic nerve fibers, but there is no detectable expression in endocrine cells in the islet.
INTRODUCTION
Alzheimer’s disease (AD) is a chronic and progressive neurodegenerative disorder characterized by extracellular deposition of amyloid-β peptide and intraneuronal aggregation of hyperphosphorylated tau [1]. Epidemiological studies have suggested that type 2 diabetes mellitus (T2DM) is a risk factor for AD. Individuals with probable T2DM have nearly a two-fold higher risk of AD than individuals without diabetes [2]. AD mouse models are susceptible to insulin resistance, obesity, and glucose intolerance [3, 4]. TauKO mice show pancreatic β cell dysfunction and glucose intolerance [5]. Amyloid-β and hyperphosphorylated tau promote the diabetic phenotype [6].
Tau is a major neuronal microtubule-associated protein. The main function of tau is to stimulate tubulin assembly and to stabilize microtubule structure. Tau protein plays an important role in axonal transport [7], neurite outgrowth, and microtubule dynamics [8]. Rodent tau shows approximately 90% homology with human tau. Tau is a phospho-protein. Phosphorylation regulates tau’s function and subcellular localization. In AD brain, tau is hyperphosphorylated and aggregated into neurofibrillary tangles [9, 10], a histopathological hallmark of AD.
Tau is mainly expressed in the central nervous system. It was reported more than 20 years ago that tau may also be expressed in rat skeletal muscle, heart, lung, testis, and kidney [11]. Tau may be expressed in pancreatic islets, and tau may play a significant role in pancreatic β-cells. Recent studies also suggested expression of various isoforms of tau in human and mouse pancreatic islets [5, 6] and in RIN-5F pancreatic β-cell line [12]. Increased levels of total tau and phosphorylated tau have been observed in pancreas of individuals with T2D [13] and of a transgenic mouse model of AD and T2D [14]. Exposing RIN-5F cells to sera from T2D patients increases tau expression [12]. Overexpression of tau in RIN-5F cells inhibits insulin secretion [15]. Although the endocrine systems have many functional similarities to the nervous system [16], pancreatic tau expression is not very clear. In the present study, we investigated tau expression in mouse pancreases by western blots and immunohistochemical staining. We found that tau is restricted to autonomic nerve fibers in both islets and exocrine pancreas, but is not detectable in endocrinal cells in the islet.
MATERIALS AND METHODS
Animals
Adult C57BL/6, TauKO [17], and 3xTg-AD [18] mice were purchased from the Jackson Laboratory, bred in our animal colony and housed under a 12 h light-dark cycle, with ad libitum access to food and water. All animal procedures and use were as per the protocol approved by our Institutional Animal Care and Use Committee in accordance with the PHS Policy on Humane Care and Use of Laboratory Animals.
Western blot analysis
Mice were sacrificed by cervical dislocation. The brain and pancreas were quickly dissected out, cooled down in a large volume of ice-cold phosphate-buffered saline (PBS) and then stored at –80°C until used.
The brain or pancreas were homogenized to 10% (wt/vol) final concentration in buffer containing 50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 2 mM EDTA, 2 mM Na3VO4, 50 mM NaF, 1 mM 4-(2-aminoethyl) benzenesulfonyl fluoride hydrochloride (AEBSF), and 10μg/ml each of aprotinin, leupeptin, and pepstatin. The homogenate was boiled in Lemaeli buffer for 5 min, and protein concentrations were measured by using A660 kit (Pierce, Rockford, IL) according to the manufacturer’s instructions. The samples were subjected to 10% SDS-PAGE and electrically transferred to a polyvinylidine difluoride membrane (Milipore, Bedford, MA). The membrane was blocked with 5% fat-free milk in Tris-buffered saline (TBS), and then incubated with primary antibodies listed in Table 1 in 5% fat-free milk and 0.5% NaN3 in TBS overnight at room temperature. After washing three times with 0.5% Tween-20 in TBS (TBST), the membrane was developed with species-matched peroxidase-conjugated secondary antibodies (Jackson ImmunoResearch, West Grove, PA) for 2 h. Following three washes with TBST, the membrane was incubated with ECL kit (Pierce) for 1 min, and then exposed to X-ray film (Kodak, Rochester, NY, USA).
Antibodies used in this study
Mono-, monoclonal; p-, phosphorylated; up-, unphosphorylated; Poly-, polyclonal; M, Mouse; R, Rabbit.
Immunoprecipitation
Brain or pancreas homogenate was adjusted to 50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 0.2% sodium deoxycholate, 0.1% NP-40, 0.1% Triton X-100, 1 mM Na3VO4, 50 mM NaF, 2 mM EDTA, 1 mM AEBSF, and 10μg/ml each of aprotinin, leupeptin, and pepstatin and incubated on ice for 20 min. The homogenate was then centrifuged at 15,000×g, 4°C for 5 min. The supernatant was incubated with monoclonal tau antibody Tau-1 pre-coupled onto protein G beads overnight at 4°C. The beads were washed with lysis buffer twice and with TBS twice. Then the bound proteins were eluted from beads by boiling them in 2×Laemmli sample buffer and subjected to western blots developed with R134d and Tau-1.
Immunofluorescent staining
Mice were transcardially perfused with saline followed by phosphate-buffered 4% paraformaldehyde, and the pancreas was excised and post-fixed in the same fixative overnight at 4°C. The pancreas was dehydrated and embedded in paraffin using the standard histological procedure. Paraffin sections, 6μm in thickness, were prepared, and then deparaffinized, rehydrated and subjected to immunofluorescent staining. After heat antigen retrieval in pH 6.0 citrate buffer, sections were blocked in normal goat serum, and incubated overnight at 4°C with primary antibodies R134d, Tau-1, Tau-5, anti-synapsin 1, or a combination of Tau-1 or Tau-5 and anti-synapsin 1 for double labeling. After washed in PBS, the sections were incubated with species-matched Alexa Fluor 488 or 555-conjugated secondary antibodies (1 : 1,000 each, Life Technologies, Rockford, IL) and for nuclear staining with TO-PRO 3 iodide (1 : 1,000, Thermo Fisher Scientific) at room temperature for 2 h. Sections were then washed in PBS, covered with anti-fade mounting medium and visualized using a Nikon EZ-C1 laser scanning confocal microscope.
RESULTS
No detectable tau in mouse pancreas by western blots
As a major neuronal microtubule associated protein, tau is predominantly expressed in the central nervous system. To investigate whether tau is expressed in the pancreas, homogenates of pancreas from C57BL/6 wild type (WT) and tauKO mice were subjected to western blots. The brain homogenates of WT and TauKO mice were used as positive and negative controls, respectively. The western blots developed with polyclonal pan-tau antibodies, R134d and 92e [19], revealed 50–65 kDa immunoreactive tau bands in WT but not in TauKO mouse brains (Fig. 1A), confirming the specificity of R134d and 92e toward tau. However, tau was not detected in WT and TauKO mouse pancreases, even when 60-folds more pancreatic proteins than the brain proteins were loaded for the western blots (Fig. 1A). Interestingly, the blot developed with 77G7, a monoclonal pan-tau antibody against the microtubule binding repeats [20], showed ∼50-kDa immunosignals in both brains and pancreases of WT and TauKO mice (Fig. 1B), suggesting that this protein is a non-tau protein that cross reacts to 77G7 or the secondary antibody used. In addition, 77G7 blot showed weak but corresponding immunosignals at 25 kDa in mouse pancreas (Fig. 1B).
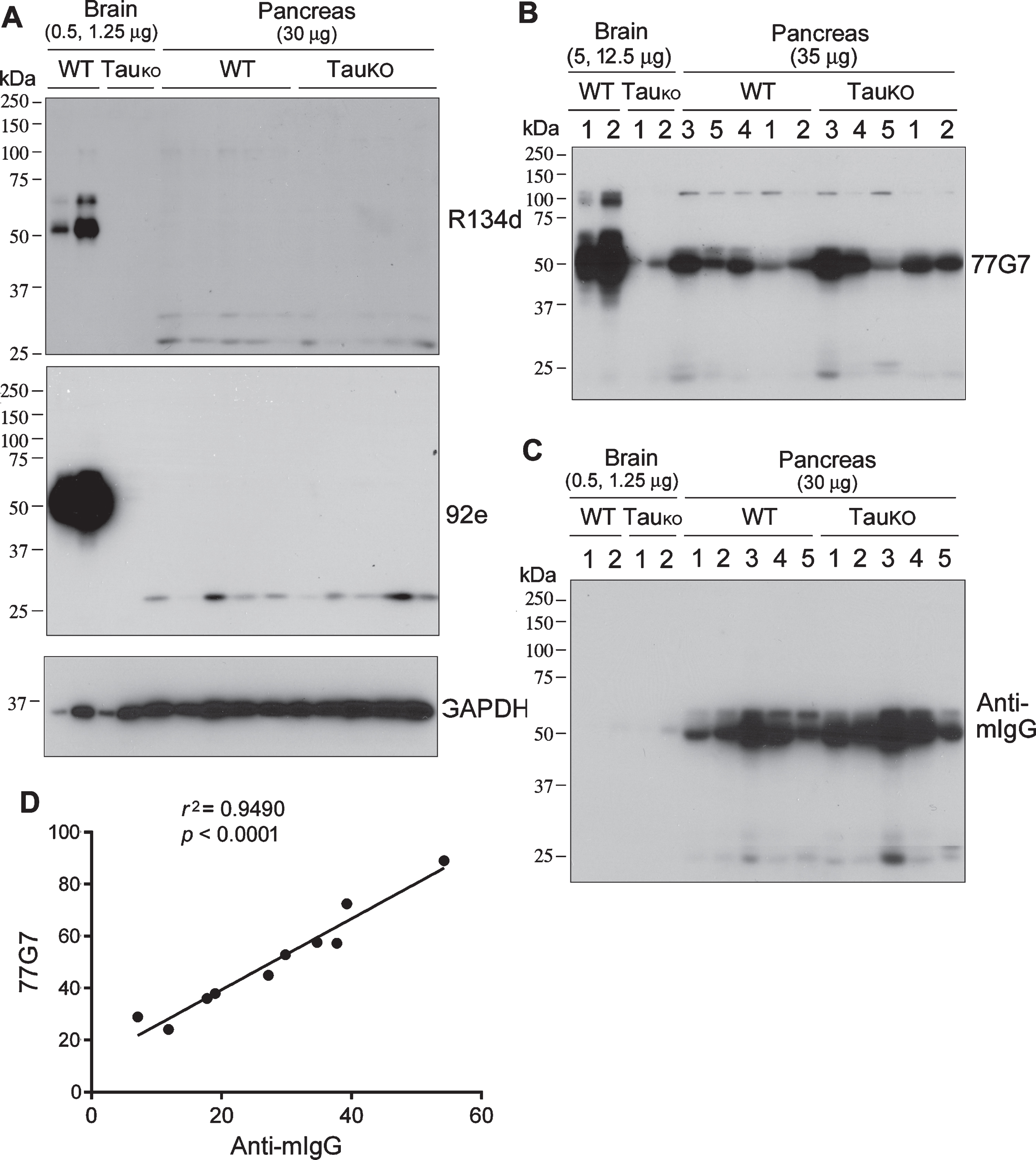
Tau is not detectable in the mouse pancreas by western blots developed with tau antibodies produced in rabbits. Various amounts of brain or pancreas homogenates from C57BL6 wild type (WT) or tau knockout (TauKO) mice were analyzed by western blots developed with rabbit polyclonal tau antibodies R134d or 92e (A) or mouse monoclonal tau antibody 77G7 (B). C) Western blot as in panel A was developed with anti-mouse IgG without primary antibody. D) 77G7 immunosignals of pancreas in panel B was plotted against the anti-mIgG immunosignals in panel C. Pearson correlation was employed.
The pancreatic islets account for 1–2% of the pancreas volume and receive 10–15% of its blood flow. The 50 kDa and 25 kDa immunosignals only recognized by mouse antibody led us to speculate that they could be the heavy and light chains of IgG. To verify this 50-kDa immunosignal is the mouse IgG heavy chain, the blot was developed with HRP-anti-mouse IgG without the primary antibodies. We also found strong and similar immunosignal at ∼50 kDa in the pancreas (Fig. 1C), as seen in the blots developed with 77G7 (Fig. 1B). Furthermore, the anti-mIgG immunosignal in the pancreas was positively and strongly correlated with 77G7 immunosignal (Fig. 1D). Thus, these data strongly suggest that the mouse pancreas does not express tau and that 50-kDa immunosignal detected by mouse 77G7 most likely is mouse IgG heavy chain.
No detectable phosphorylated tau in mouse pancreas
Tau is a phospho-protein, and phosphorylated tau was previously reported to be present in the pancreas [5]. Thus, we analyzed mouse pancreas homogenates by western blots developed with several site-specific and phosphorylation dependent tau antibodies (Table 1). Similar to blots in Figure 1B, there were ∼50-kDa immunosignals in the pancreases of WT and TauKO mice, but no or very weak signals in the brain, in blots developed with monoclonal mouse antibodies, AT8 (pSer202/Thr205), PHF-1 (pS396/404), and Tau-1 (non-phosphorylated tau at Ser195, Ser198, Ser199, and Ser202) (Fig. 2A). No immunosignals were observed in the pancreas of either WT or TauKO mice in the blots developed with rabbit-generated antibodies, anti-pSer199-tau and anti-Ser396-tau (Fig. 2B). Phosphorylated tau was found in WT, but not TauKO mouse brains, confirming the specificity of these antibodies.
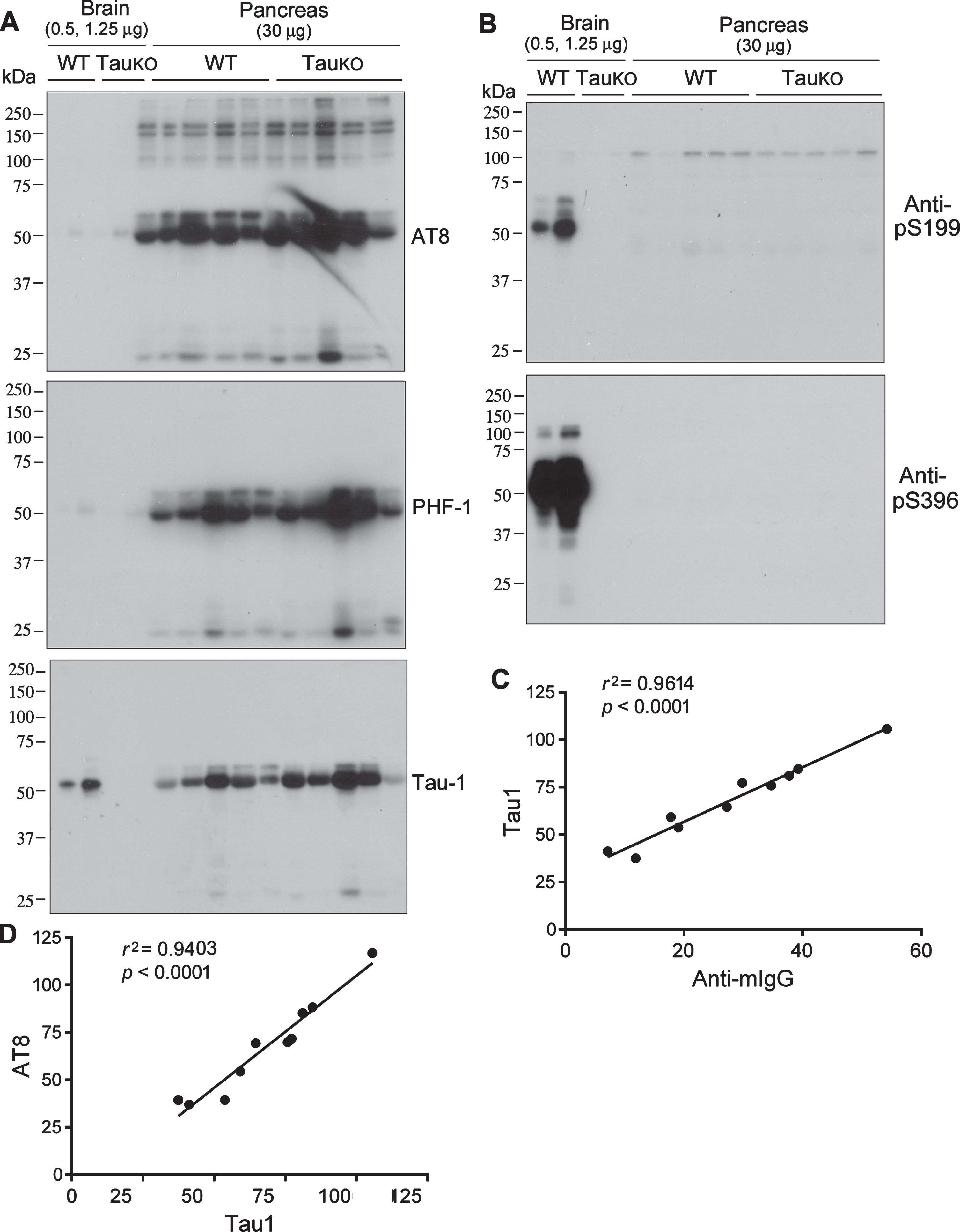
Phosphorylated tau is not detectable in mouse pancreas. Various amount of brain or pancreas homogenates from WT and TauKO mice were analyzed by western blots developed with site specific and phosphorylation dependent tau antibodies produced in mouse (A) or in rabbit (B). Tau-1 immunosignal of pancreas homogenates was plotted against anti-mIgG immunosignal (C) or AT8 immunosignals (D). Pearson correlation was used for their correlation.
We analyzed the relationship between the immunosignals of Tau-1 and anti-mIgG (without primary antibody) in mouse pancreas by Pearson correlation analysis. We found a great positive correlation between Tau-1 and anti-mIgG immunosignals (Fig. 2 C). Tau-1 antibody recognizes tau that is not phosphorylated atS195/198/199/202. AT8 recognizes tau phosphorylated at Ser202 and Thr205. Opposite immunoreactions are usually seen by these two antibodies. In contrast, we found that Tau-1 immunosignal was strongly and positively correlated with AT8 immunosignal (Fig. 2D). Taken together, these results suggest that the ∼50-kDa immunosignals in the mouse pancreas detected by mouse antibodies are mouse IgG heavy chain and not phospho-tau.
Tau was reported to be expressed in human and mouse islets [5 , 15]. Because islets account for only 2% of the whole pancreas, our failure to detect tau by western blots could be due to tissue dilution. We thus incubated 140μg or 300μg proteins of pancreas crude extracts from WT and TauKO mice with Tau-1 to enrich tau and analyzed the immunoprecipitates with western blots. We found that tau was immunoprecipitated by Tau-1 from brain extract of WT mice, but not TauKO mice and almost no tau was found in unbound brain fraction by Tau-1 (Fig. 3A). Thus, Tau-1 immunoprecipitated tau from mouse brain effectively and specifically (Fig. 3A,B). However, no detectable tau was immunoprecipitated from up to 300μg protein of pancreas extract in blots developed with tau rabbit polyclonal antibody R134d. Blots developed with mouse monoclonal antibody Tau-1 showed ∼50 kDa-IgG heavy chain in the immunocomplexes from brain or pancreas of WT and TauKO mice (Fig. 3B).
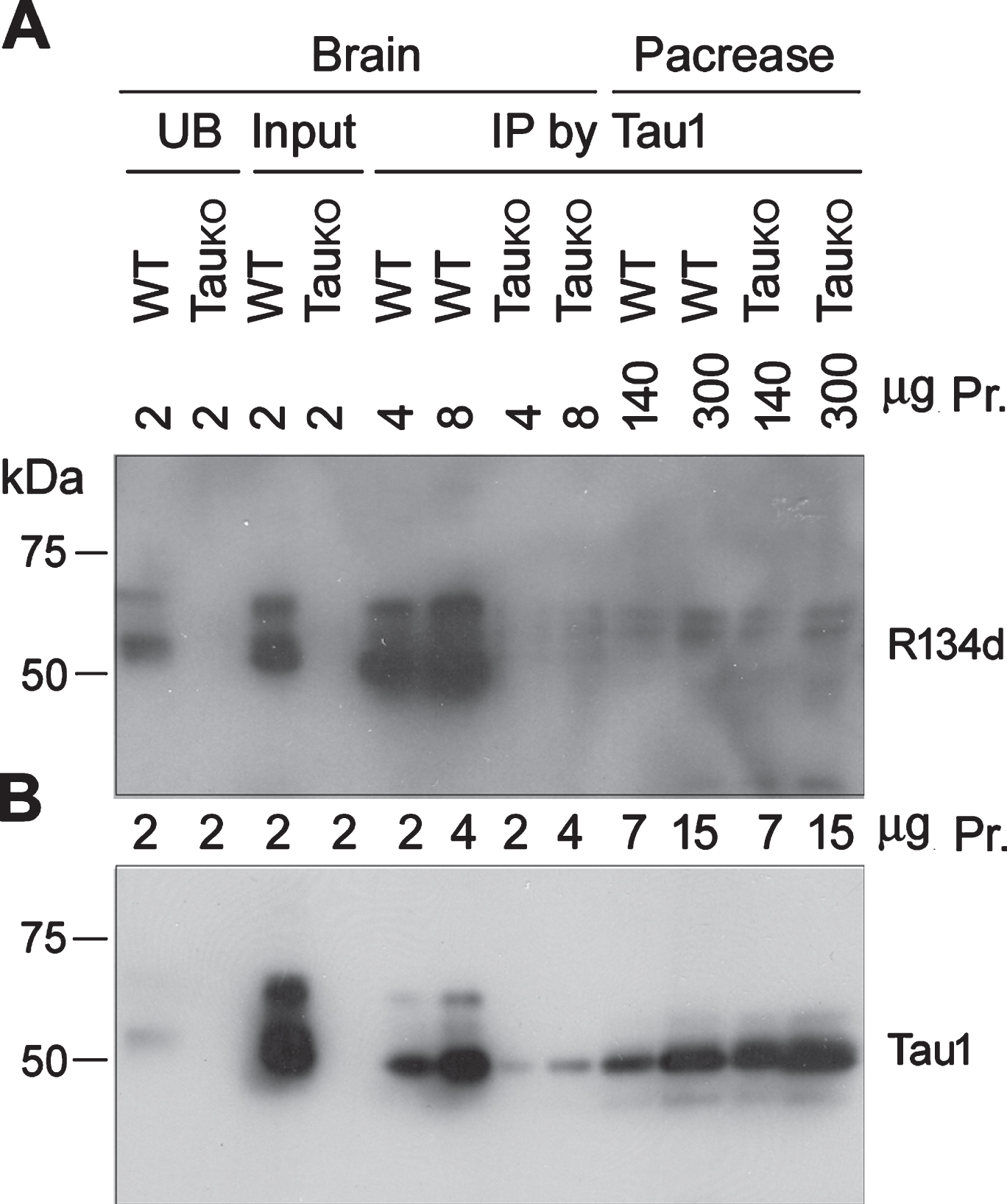
No detectable tau is immunoprecipitated from mouse pancreas. Crude extracts of mouse brain or pancreas were incubated with Tau-1 to immunoprecipitate tau. Immunoprecipitated tau from various amounts of corresponding crude extracts was analyzed by western blot developed with R134d (A) or Tau-1 (B).
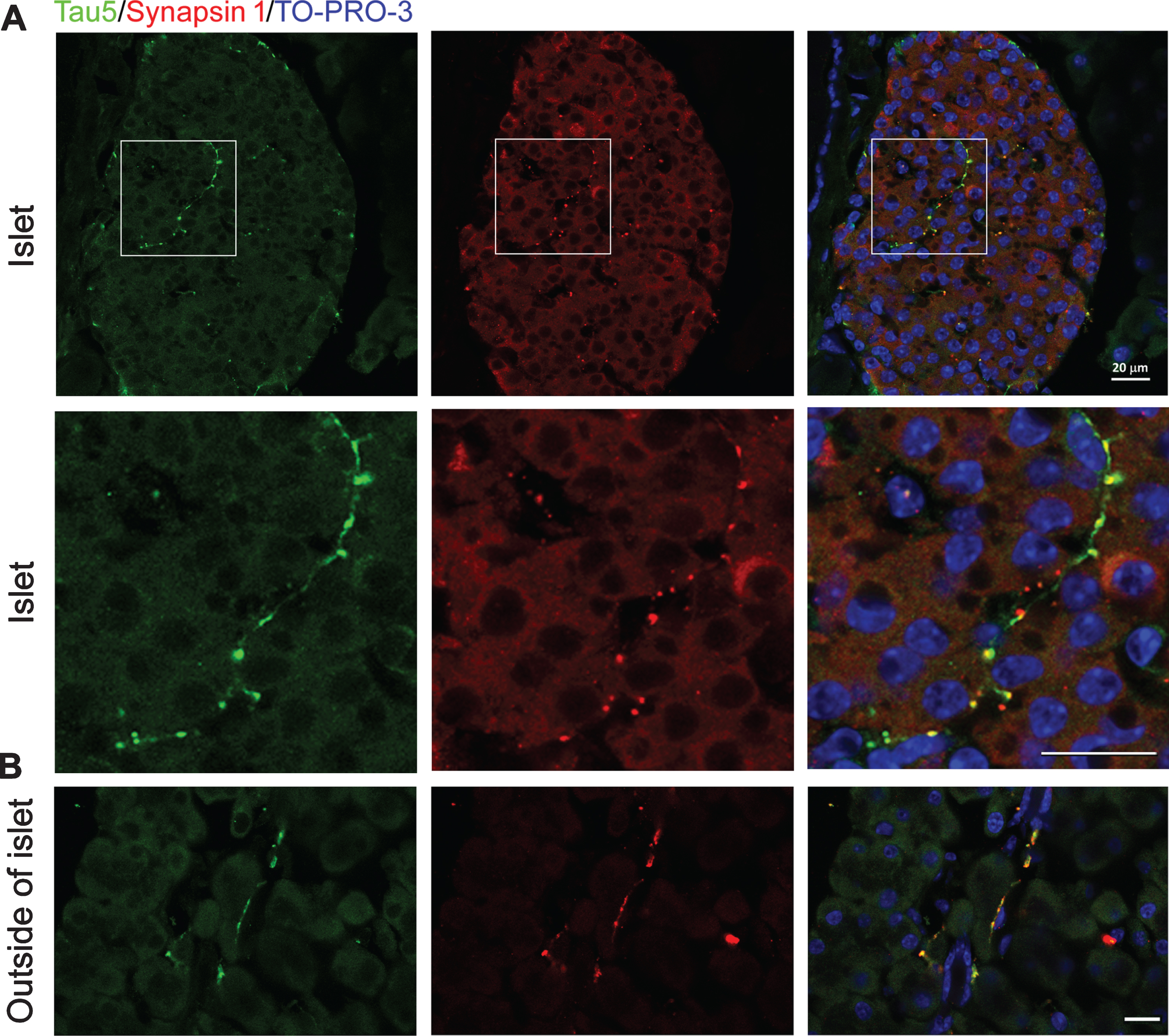
Tau displays beaded strings morphology in both islets and exocrine pancreas
The above western blot data indicate that the expression level of tau in the pancreas, if any, is undetectable. To visualize the tau protein in situ in the pancreas, especially in the islets, we immunostained pancreatic sections with R134d. Intriguingly, immunostaining with R134d revealed bright beaded strings in the pancreatic islets in wild-type mice; these beaded strings both encapsulated the islet and penetrated and densely populated inside the parenchyma of the islet (Fig. 4). Importantly, tau immunoreactivity did not show clear cellular localization within endocrine cells. In addition, tau-positive beaded strings were also present in the exocrine pancreas, despite a much sparser population (Fig. 4). These immunostained beaded strings looked like autonomic nerve fibers, which is consistent to neuronal expression of tau. By contrast, the pancreas of tau knockout mice did not show any specific tau staining either in the islets or in the exocrine regions (Fig. 4).
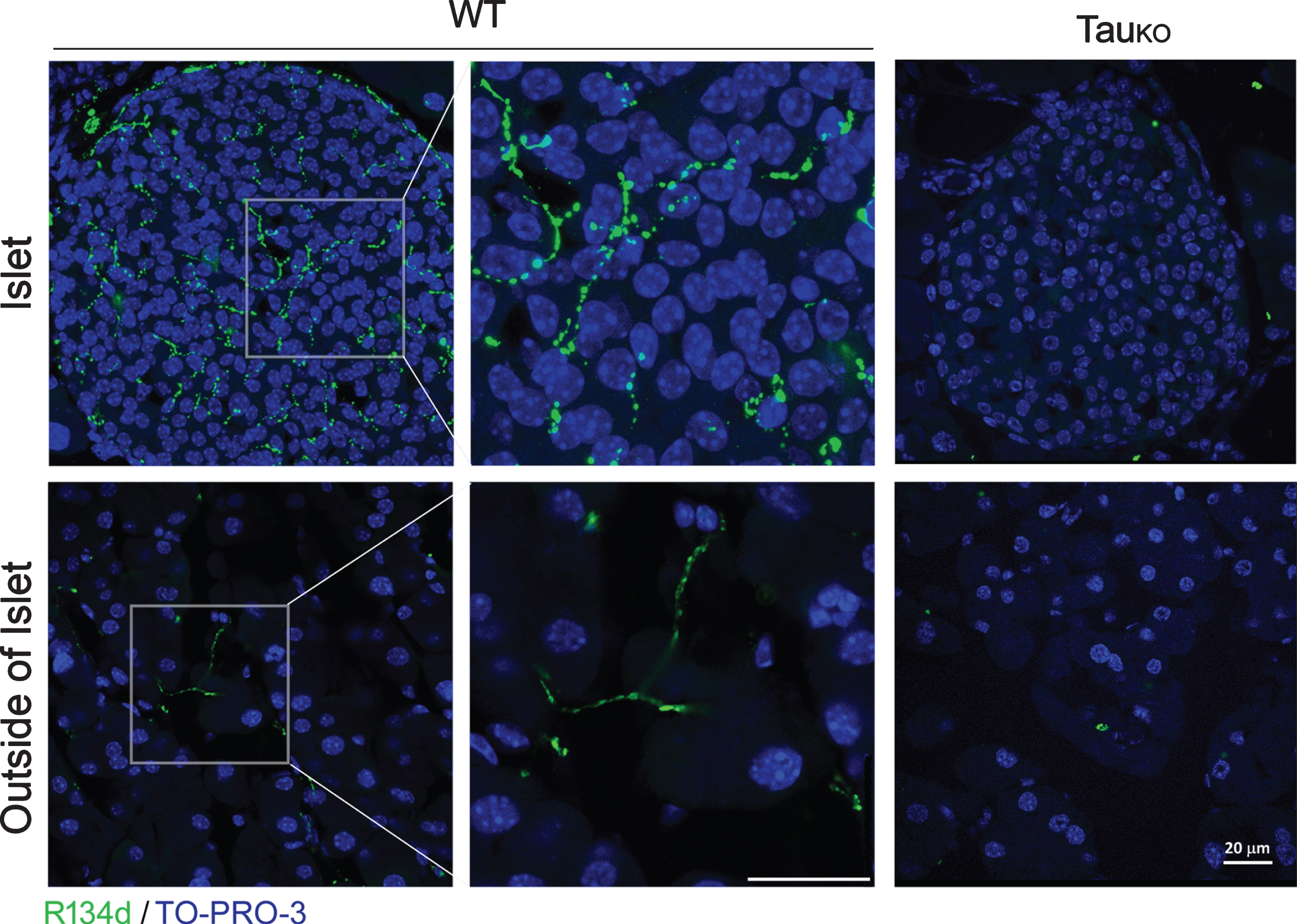
Beaded-strings distribution of tau crosses endocrine and exocrine cells of mouse pancreas. Paraffin sections of pancreases of WT and TauKO mice were immunostained with R134d. TO-PRO-3 was used to stain nuclei. Scale bar: 20μm.
Tau is co-localized with synapsin-1 in autonomic nerve fibers in the pancreas
To reveal the identity of tau-positive beaded-strings, we employed another pan-tau antibody Tau-5 and double-stained mouse pancreatic sections with rabbit polyclonal anti-synapsin 1 antibody; synapsin 1 is a commonly used presynaptic marker. We found the same immunostaining pattern of the pancreas with Tau-5 as with R134d. In addition, Tau-5 staining showed complete colocalization with synapsin-1 in both the islets (Fig. 5A) and the exocrine pancreas (Fig. 5B); whereas the pancreas of tau knockout mice only showed synapsin 1 but no Tau-5 (Fig. 5A,B) immunoreactivity. The morphology of synapsin-1 immunoreactive varicosities is consistent with a recent study showing the autonomic innervation of mouse pancreas [21].
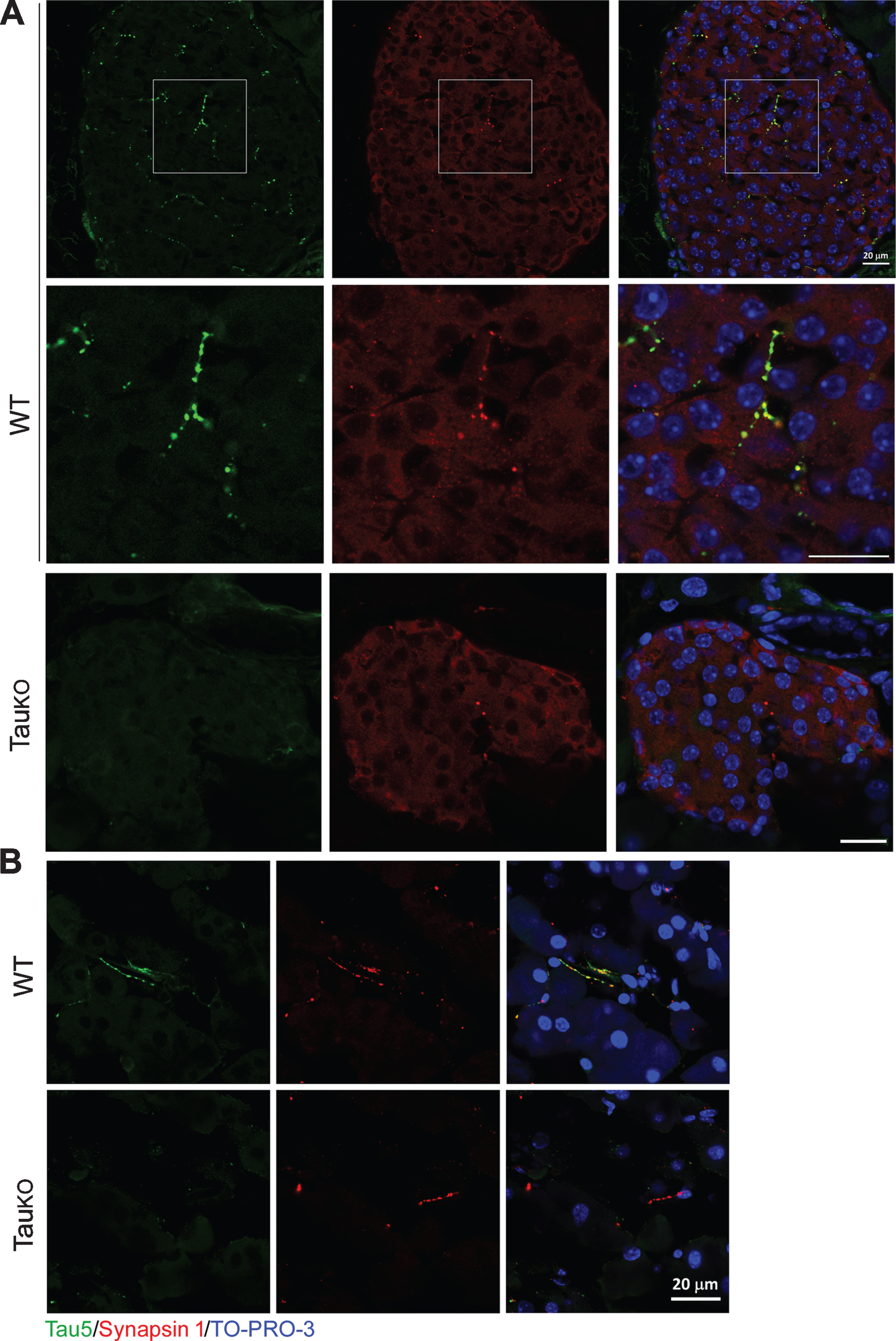
Pancreatic tau is localized in axons of both endocrine and exocrine glands. Mouse pancreas sections were dual-immunostained with Tau-5 (green) and Synapsin I (red). In both Islets (A) and exocrine gland (B) of wild-type mice, Tau-5 and Synapsin I were completely colocalized in the beaded strings; whereas in TauKO mice no Tau-5 immunoreactivity was observed in Synapsin I positive beaded strings. Scale bar: 20μm.
We then double-immunostained the pancreas with Tau-1 and anti-synapsin 1. Consistently, we found that similar beaded-string immunosignal crossed the cells, but cells of the islets (Supplementary Figure 1A) and outside of the islets (Supplementary Figure 1B) were not immunostained by Tau-1, which was co-localized with the synapsin 1 (Supplementary Figure 1) in both inside and outside of the islets.
Taken together, our immunohistochemical data indicate that tau is restricted in autonomic nerve fibers in both islets and exocrine glands, but not in endocrine cells in the islets or exocrine cells outside of the islets.
Tau is expressed in autonomic nerve fibers in pancreas of 3xTg-AD mice
The 3xTg-AD is a widely used mouse model of AD, in which tauP301 L is overexpressed under Thy 1 promoter [18]. In this AD mouse model, AD-like neuropathology was reported to extend to the pancreas, leading to insulin resistance [22]. To determine the distribution of tau in the pancreas of 3xTg-AD mouse, we performed double immunostaining with Tau-5 and synapsin 1. We found beaded-strings Tau-5 immunoactivity crossed the cells in the islets (Fig. 6A) and outside of the islets (Fig. 6B), which were similar to those in C57BL/6 wild-type mouse pancreas. Dual immunostaining of pancreas sections of 3xTg-AD mice with Tau-1 and anti-synapsin 1 also showed complete colocalization of the two markers in beaded strings in both inside the islets (Supplementary Figure 2A) and the exocrine regions (Supplementary Figure 2B). These results indicate that tau is also expressed in the autonomic nerve fibers in 3xTg-AD mouse pancreas and confirm the expression of tau in autonomic nerve fibers, but not in endocrine and exocrine cells of the pancreas.
Tau is expressed in axons within the pancreas in 3xTg-AD mice. Sections were immunostained with Tau-5 and anti-synapsin 1. Beaded-string co-localization of Tau-5 and anti-synapsin 1 was found in islets (A) and exocrine glands (B). Scale bar: 20μm.
DISCUSSION
Tau is a major neuronal microtubule-associated protein, but its peripheral expression including pancreas islets was reported. In the present study, we investigated tau in the mouse pancreas biochemically and immunohistochemically by using several monoclonal and polyclonal tau antibodies. We found no detectable level of tau expression by using western blots as well as after immuno-enrichment by tau antibody. However, by immunohistochemistry we observed bright and beaded tau immunosignal in both the endocrine and exocrine glands, which were completely colocalized with synapsin I. These positive tau stainings were absent in TauKO mice. Thus, tau is expressed in axons which innervate both islets and the exocrine glands in the pancreas, but not expressed in the endocrine or exocrine cells.
As a neuronal protein, tau was also found in several other peripheral tissues [11]. In the present study, we used several mouse monoclonal and rabbit polyclonal tau antibodies to analyze tau in pancreas. We found a 50 kDa band in pancreas homogenates from both WT and TauKO mice in the blots developed with the mouse antibodies, but not in the blots developed with rabbit antibodies, regardless of phospho-dependent or independent tau antibodies. Furthermore, the 50 kDa band was developed with goat anti-mouse IgG, but not with goat-anti-rabbit IgG. Thus, the 50 kDa band is mouse IgG heavy chain, but not tau. In addition, several tau antibodies showed non-specific immunoactivity [23, 24]. In the present study, we used TauKO mice to validate the specific immunoactivity of tau biochemically and immunohistochemically.
Previous studies showed expression of tau in the islets, which occupy 2% of pancreas in mass. Therefore, we immuno-enriched tau by Tau-1 antibody from 140 and 300μg proteins of pancreas crude extract, but we still failed to detect tau. Tau-1 immunoprecipitated tau from mouse brain crude extract efficiently, which serves as a control for the assay. Thus, tau expression in the pancreas, if any, is not detectable biochemically.
A recent study showed that autonomic innervation of mouse pancreas immunostained by anti-synapsin 1 appeared as varicosities [21]. By using tau antibodies, Tau-5, Tau-1 and R134d, we observed similar beaded-immunosignaling crossing the sections of islets and exocrine glands, suggesting tau may be localized in nerve fibers. Double immunostaining showed beaded synapsin 1 immunosignal, which was completely colocalized with Tau-1 in both endocrinal and exocrinal glands, confirming expression of tau in autonomic nerve fibers.
In summary, we found tau in the mouse pancreas can be visualized immunohistochemically as beaded strings winding along islets and exocrine glands, which are colocalized with synapsin 1, but it is not detectable biochemically in wild-type C57BL/6 and 3xTg-AD mice. These data indicate that expression of tau in mouse pancreatic islets is restricted to autonomic nerve fibers.
Footnotes
ACKNOWLEDGMENTS
This work was supported in part by Nanjing Medical University, New York State Office for People with Developmental Disabilities, and Nantong University and by grants from the U.S. Alzheimer’s Association (Grant DSAD-15-363172) and the Neural Regeneration Co-innovation Center of Jiangsu Province.