
Abstract
Frontotemporal lobar degeneration (FTLD) is a progressive neurodegenerative syndrome. Defects of copper (Cu) and iron (Fe) homeostasis are involved in the development of several neurodegenerative diseases and their homeostasis is interconnected by the Cu-protein ceruloplasmin (Cp), responsible for Fe oxidative state. In this study we assessed Fe, transferrin (Trf), ferritin, Cp specific activity (eCp/iCp), Cp/Trf ratio, and Trf saturation in 60 FTLD patients and 43 healthy controls, and discussed the results in relation to Cu homeostasis. The significant decrease of the eCp/iCp in the FTLD patients supports the involvement of Fe imbalance in the onset and progression of FTLD.
INTRODUCTION
Frontotemporal lobar degeneration (FTLD) is a progressive neurodegenerative syndrome, highly heterogeneous, both in its clinical phenotypes and pathology. It is characterized by focal atrophy of the frontal and/or temporal lobes of the brain with consequent deterioration of behavior, personality, executive functions, and language. On the basis of its clinical presentation, it has been classified as behavioral variant of frontotemporal dementia (bvFTD) [1, 2], non-fluent variant of primary progressive aphasia and semantic variant of primary progressive aphasia (nvPPA and svPPA, respectively) [1, 3]. Furthermore, FTLD could overlap with extrapyramidal syndromes, like progressive supranuclear palsy (PSP) [4] or corticobasal syndrome (CBS), and with motor neuron disease (FTD-MND) with both upper and lower motor neuron signs [5–7]. In these presentations, posterior frontal lobe atrophy (including premotor cortices) is associated with FTD-MND; midbrain atrophy with PSP and atrophy in frontal/parietal regions and in the basal ganglia for CBS.
FTLD account for 5–15% of all dementia with an onset between the ages of 40 and 65 and it is among the commonest cause of presenile dementia [8].
The neuropathological signature of FTLD reflects its clinical heterogeneity, and the most common forms are represented by FTLD-tau [9] or FTLD-transactive response DNA-binding Protein (FTLD-TDP43) [10]. Furthermore, FTLD-FET shows the aggregation of all members of the FET protein family, comprising fused in sarcoma protein, Erwing’s sarcoma protein, and TATA-binding protein associate factor 2N [11].
In the light of this complex scenario describing an extremely heterogeneous disease, in both clinical manifestation and underlying molecular mechanisms, further efforts are needed to find new molecular biomarkers for its diagnosis, classification and treatment.
To date, countless reports support a strong correlation between copper (Cu) and iron (Fe) dyshomeostasis and neurodegeneration. These transition metals are essential for all living organisms, acting as cofactor of metabolic enzymes. Altered homeostasis of these metals, interconnected by ceruloplasmin (Cp), the cross-talking protein between Cu and Fe metabolism, and responsible for Fe distribution, results in neurodegenerative diseases, such as Menkes’ and Wilson’s disease and aceruloplasminemia [12]. Furthermore, defects in their handling and homeostasis are tightly implicated in Alzheimer’s disease (AD), where it has been found an accumulation of Fe in the brain and an overall copper misbalance [13–16]. Indeed, in AD it has been demonstrated that Cu and Fe prompt and/or stabilize the formation of plaques of amyloid-β, the hallmark of AD [17–19]. However, a possible involvement of derangement of these two essential metals in other neurodegenerative diseases is not well defined yet.
In a previous work from our group, we performed the analysis of an extensive panel of Cu-linked biomarkers (e.g., Cu levels, Cp, Cu:Cp ratio) in the serum of patients affected by bvFTD and PPA. The results obtained discourage a Cu involvement in FTLD, even though they need to be confirmed in other and bigger patient populations [20].
On the other hand, a magnetic resonance imaging (MRI) study, performed on postmortem brains of patients affected by different neurodegenerative diseases (i.e., FTLD, AD, amyotrophic lateral sclerosis, Lewy body disease), demonstrated that only FTLD patients were characterized by a significant increase in Fe accumulation in the deep gray nuclei, compared with the other diseases analyzed [21]. This evidence was further confirmed by Gazzina et al., which demonstrated that FTLD neurodegeneration could also be related to genetic impairment in Fe uptake, which fosters Fe deposition in the basal ganglia [22].
Thus, in the present study we analyzed the serum profile of 60 FTLD patients, affected by either bvFTD or PPA. We assessed the level of a wide panel of Fe homeostasis markers (e.g., Fe, transferrin, ferritin), discussing their relationship with serum Cu levels, in order to find a possible correlation between the Fe/Cu serum homeostasis and FTLD.
METHODS
Subjects
For this study, 103 subjects were included, namely 60 FTLD (bvFTD, n = 34 and PPA, n = 26) and 43 healthy controls. Demographic and clinical characteristics are reported in Table 1. Healthy controls were mainly spouses, selected among subjects without any sign of neurological pathology and with normal cognitive function. Patients were recruited at the Memory Clinic of the IRCCS Fatebenefratelli in Brescia (Italy) and underwent clinical and neurological examinations. The inclusion criteria for patients were the diagnosis of FTLD according to current clinical criteria [1–3].
Demographic and clinical characteristics of the FTLD and healthy control individuals participant to the study
*Student t -test for continuous Gaussian distributed variables; ∧Mann-Whitney U test for non-Gaussian distributed variables; §chi-squared statistic test for categorical variables. CTRL, healthy controls; FTLD, frontotemporal lobar degeneration patients; MMSE, Mini-Mental State Examination; APOE, Apolipoprotein E; IADL, instrumental activity of daily living; BADL, basic activity of daily living; CDR, clinical dementia rating.
Serum samples were isolated according to standard procedures and stored at Fatebenefratelli Biobank (Brescia, Italy) and patients signed an informed consent (approval number 2/1992; 26/2014). The current study was approved by the Local Ethics Committee (number 53/2016).
Biochemical investigations
Fasting blood samples were collected in the morning and sera samples were separated by centrifugation (3000 rpm, 10 min, 4°C). They were then divided into 0.5 mL aliquots and rapidly stored at –80°C. The samples were shipped to Fatebenefratelli “San Giovanni Calibita” Hospital, Rome, Italy for blinded biochemical analyses. The aliquots were thawed just before the assay. All the analyses on serum were performed in duplicate on the multiple biochemical analyzer Cobas Mira Plus (ABX Diagnostic, Montpellier, France). Fe was measured by photometric test using Ferene. Briefly, Fe bound to transferrin (Trf) is released in an acidic medium as ferric iron and is then reduced to ferrous iron in the presence of ascorbic acid; ferrous iron forms a blue complex with Ferene [23]. Trf [24, 25] and ceruloplasmin (iCp, immnunological Cp) [26] levels were measured by immunoturbidimetric assays (Horiba ABX, Montpellier, France) by using specific antibodies as previously reported [27]. The enzymatic activity of CP (eCp) was measured following an automation of the manual Schosinsky o-dianisidine eCp assay [28, 29], adapted from our laboratory for multiple biochemical analyzers and previously reported [30]. For each sample, we also computed the ratio eCp/iCp, to investigate the Cp specific activity (enzymatic activity per mg of Cp concentration in IU/mg*10–1). Furthermore, we also computed and log transformed for Gaussianity the ratio between Cp and Trf serum concentrations (Cp/ Trf). Trf saturation (% Trf-sat) was calculated as follows: serum iron was divided for the total iron-binding capacity (TIBC = Trf in mg/dL *1.25) and multiplied by 100.
Statistical analysis
Statistical analyses were performed using SPSS (version 21.0, IBM SPSS Statistics for Windows). Demographic and clinical characteristics were analyzed through means and standard deviations (SD) for continuous variables and frequencies for discrete variables. The Kolmogorov-Smirnov test was performed in all continuous variables to define the presence of normality. To compare demographic and clinical characteristics student t-test or Mann-Whitney U test was performed (for continuous Gaussian or non-Gaussian distributed variables) and chi-squared statistic test was performed for categorical variables. In order to take into account correlation among biological variables all measured for whole study sample, multivariate ANOVA model (MANOVA) with post-hoc test with Bonferroni correction was used to compare biological variables between the two study groups. Statistical significance was set at p < 0.05.
RESULTS
Table 1 reports the demographic and the comparison of the clinical characteristics of patients (n = 60) and healthy controls (n = 43). The groups are age and sex-matched and the statistical analysis of semantic and phonemic fluency tests, the Token test, the Rey-Osterrieth complex figure copy and recall, clearly show significant differences in cognitive performance (p < 0.001) between the two groups (Table 1)
The analyses of the Fe serum markers performed in the two groups (adjusted for sex) are reported in Table 2. The MANOVA model did not show an overall significance (p = 0.165); indeed, the comparisons between the healthy controls and the FTLD patients did not show any statistically significant differences in Fe, Tfr, iCp, ferritin, eCp, the ratio Cp/Tfr, Tfr-saturation, and TIBC. A significant difference was found only between groups in the ratio eCp/iCp (p = 0.003), with patients with FTLD showing a decrease ratio eCp/iCp (Fig. 1A).
Serum iron profile of the sample: multivariate ANOVA (MANOVA)
iCp, immunological ceruloplasmin; eCp, enzymatic activity of ceruloplasmin; eCp/iCp, ceruloplasmin specific activity; Cp/Trf, ceruloplasmin/Transferrin, index of oxidative stress related to iron metabolism; Tfr-sat, transferrin saturation; TIBC, total iron binding capacity.
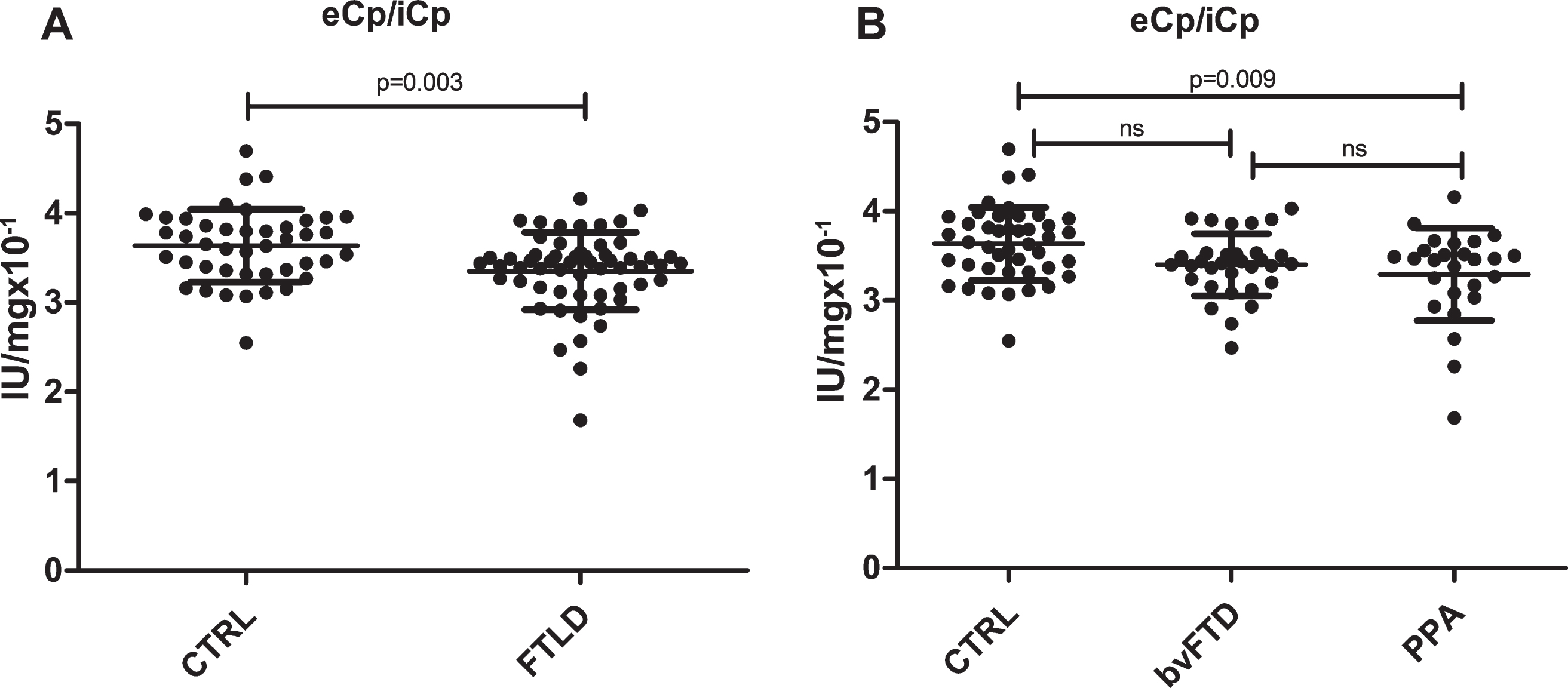
FTLD patients show a decrease eCp/iCp ratio. Graphic representation of the MANOVA analysis performed between A) healthy controls (n = 43) and FTLD patients (n = 60) and B) among PPA (n = 26), bvFTD (n = 34) patients, and healthy controls (n = 43).
We further stratified patients on the basis of clinical phenotypes, i.e., PPA and bvFTD. In Table 3 are reported the results of MANOVA in the comparison of the analyzed variables for the three groups (overall significance was p = 0.320). Once more, no significant differences were found in most of the variables investigated except for the ratio eCp/iCp, being healthy controls statistically different from PPA but not from bvFTD (Fig. 1B).
Serum iron profile of the patients stratified on the basis of the FTLD subtypes
∧CTRL versus PPA p = 0.009; CTRL versus bvFTD p = 0.123; bvFTD versus PPA p = 1.000.
DISCUSSION
The involvement of metal dyshomeostasis in the onset and progression of neurodegenerative diseases, such as AD and Parkinson’s disease (PD) has been extensively described [31, 32]. Furthermore, in the latest years, ferroptosis, a Fe-regulated cell death, has been strictly related to neurodegeneration, strengthening the key role played by Fe in neurological diseases [33, 34].
On this basis, we focused our attention on FTLD in order to delineate the involvement of Cu and Fe balance in the development of the pathology. Our previous data demonstrated that Cu homeostasis was not altered in FTLD [20].
In this paper, on the basis of literature reporting of Fe accumulation in the brain of FTLD patients, in particular in the basal ganglia [21, 22], we investigated the serum Fe profile of FTLD patients and focused our analysis on two different FTLD phenotypes, PPA and bvFTD. We considered in our analysis a number of biomarkers of Fe homeostasis, in order to obtain a comprehensive picture of the metal serum profile in this pathology. We found that none of the parameters analyzed (Fe, ferritin, Trf, Trf-sat, TIBC, iCp and eCp and their ratios) significantly differs between the groups analyzed with the exception of the Cp specific activity (eCp/iCp ratio), being significantly decreased in the FTLD patients. A decreased functionality of Cp suggests potential abnormalities in Fe handling, since Cp controls Fe oxidative state and Fe mobility [35]. Ceruloplasmin catalyzes the oxidation of Fe2 + to Fe3 + permitting Fe to be bussed to various tissues and organs by Trf, which accepts only Fe3 +. Aceruloplasminemia is the paradigmatic disease of Cp defect and it is typified by Fe accumulation in the brain. Even though the difference between FTLD and healthy controls in terms of eCp/iCp values appears minimal, they are still effective to identify a defect of Fe handling in PPA and bvFTD. This decrease of Cp specific activity is in accordance with the observation of Fe deposits in FTLD affected postmortem brains [21], further supporting the hypothesis about the involvement of Fe imbalance in the onset and progression of FTLD. An analogous trend for serum Cp specific activity was reported by Siotto et al. (2016) [36], Brewer et al. (2010) [37], Torsdottir et al. (2011) [38], and Kristinsson et al. (2012) [39] in AD and PD patients.
The discovery of ferroptosis further links Fe metabolism to neurodegenerative disorders [40]. Ferroptosis is due to a Fe-dependent accumulation of lipid reactive oxygen species. Of note, both iron accumulation in specific regions of the central nervous system and lipid peroxidation are related to neurodegeneration. Indeed, the involvement of ferroptosis has been related to the development of several neurodegenerative disorders such as Alzheimer’s, Huntington’s and Parkinson’s diseases, amyotrophic lateral sclerosis, and Friedreich’s ataxia [41–46]. Thus, we cannot exclude that also in FTLD Fe accumulation/imbalance may lead to a neuron cell death via ferroptosis, even though at the stage of our research this remains speculative. Ceruloplasmin specific activity has been also challenged in discriminating patients from different liver pathologies e.g. Wilson’s disease, hepatic encephalopathy, and chronic liver disease [30]. It better discriminated individuals suffering of hepatic encephalopathy syndrome, presenting cirrhosis, other liver dysfunction and neuropsychiatric abnormalities, from the other liver diseases [30]. Current findings might pave the way for clinical investigations on liver failure or hypofunction in FTLD.
In light of these new results, our previous findings [20] should be taken with caution. The current data show, in fact, how it is partial and not exhaustive to study only one metal, Cu or Fe, at a time (although several biomarkers of their profile are analyzed). This is because the metabolism of Cu and Fe are strictly interconnected by the cross-talking protein Cp.
Current investigation did not detect changes in other major Fe biomarkers generally associated severe morbid conditions. Serum ferritin is a Fe storage protein and has large variations from individual to individual. Only major deficiencies have effects on physiology. States of Fe deficiency are also characterized by low Fe levels, high Trf/TBC and low Trf-sat. Ferritin is high in people with hemochromatosis and other excess iron storage disorders and in those who have had multiple blood transfusions. Fe levels are generally high in the same conditions. Ferritin is an acute phase reactant and increases in inflammation states. During malignancy and infection, ferritin increases, sequesters Fe, and reduces free Fe available to tumor cells or pathogens. High levels can be also found in liver disease, chronic infection, autoimmune disorders, sideroblastic anemia, and some types of cancer, generally associated with increased levels of Fe and Trf. Transferrin is a negative acute phase protein that decreases during inflammation. Macrophages and red blood cell internalize Trf sequestering iron, thus inhibiting microbial Fe scavenging [47]. In addition, recently, it has been shown that ferritin, either in plasma or in CSF, could have a potential role in discriminating AD patients in the preclinical phase, predicting mild cognitive impairment conversion to AD [48–50]. Authors reported that increased ferritin levels in AD suggests increase Fe level in CSF and brain that might be associated with AD through ferroptosis (reviewed in [51]).
To date, studies analyzing the modulation of serum metals homeostasis in the different forms of FTLD are lacking. Apart from the role of Cu and Fe here investigated, it has been shown the involvement of calcium dysregulation in the degeneration of FTLD-Tau neuron derived from induced pluripotent stem cells [52] and an increase of zinc, Cu, and manganese in the spinal cord of a TDP-43A315T transgenic muse model of FTLD-TDP43 [53]. However, our study has a number of limitations, consisting primarily in the diagnosis of FTLD patients based on clinical criteria that hampered to exclude a bias related to possible misdiagnosis in the early stage of dementia. Additional limitations concern a possible preclinical change in controls, and the FTLD clinical phenotypes investigated, limited to the bvFTD and PPA.
To better elucidate the role played by Cu and Fe in these complex pathologies, extensive studies in bigger cohorts that also take into account patients with other forms of FTLD such as PSP, CBD, or FTD-MND are warranted. Future in-depth analyses could lead to the identification of a new target/biomarker in the early-stage of FTLD involved in the modulation of Fe homeostasis to be used in the treatment of this complex neurodegenerative disease.