
Abstract
Background:
Perioperative, modifiable factors contributing to perioperative neurocognitive disorders (PND) have not been clearly defined.
Objective:
To determine the contribution of anesthesia lengths and the degrees of surgical trauma to PND and neuroinflammation, a critical process for PND.
Methods:
Three-month-old C57BL/6J mice were subjected to 2 h or 6 h isoflurane anesthesia plus a 5 min or 15 min left common carotid artery exposure (surgery) in a factorial design (two factors: anesthesia with two levels and surgery with three levels). Their learning and memory were tested by Barnes maze and novel object recognition paradigms. Blood, spleen, and hippocampus were harvested for measuring interleukin (IL)-6 and IL-1β. Eighteen-month-old C57BL/6J mice (old mice) were subjected to 6 h isoflurane anesthesia or 2 h isoflurane anesthesia plus 15 min surgery and then had learning and memory tested.
Results:
Three-month-old mice with 15 min surgery (long surgery) under 2 h or 6 h anesthesia performed poorly in the learning and memory tests compared with controls. Anesthesia alone or anesthesia plus 5 min surgery did not affect mouse performance in these tests. Similarly, only mice with long surgery but not mice with other experimental conditions had increased IL-6 and IL-1β in the blood, spleen, and hippocampus and decreased spleen weights. Splenocytes were found in the hippocampus after surgery. Similarly, old mice with long surgery but not the mice with isoflurane anesthesia alone had poor performance in the Barnes maze and novel object recognition tests.
Conclusion:
Surgical trauma, but not anesthesia, contributes to the development of PND and neuroinflammation. Splenocytes may modulate these processes.
INTRODUCTION
More than 50 million patients each year have sur-gery in the United States [1]. Perioperative neurocognitive disorders (PND) are a significant complication after surgery [2–4] and are associated with increased length of hospital stay and high one-year mortality [5–7]. Various unmodifiable risk factors, such as age and preexisting cognitive disorders, have been identified for PND [2–4]. Understanding perioperative, modifiable factors contributing to its development and the mechanisms for the contribution are important to design approaches to reduce PND.
Patients with surgery are exposed to surgical tra-uma and anesthetics. The possible role of anesthe-tics in PND has been implied by early animal studies showing learning and memory impairment after a pr-olonged anesthetic exposure [8, 9]. However, no lear-ning and memory impairment after general anesthesia has also been reported [10]. Interestingly, general anesthetics can provide neuroprotection against various injurious stimuli [11, 12]. The contribution of general anesthetics to PND in the presence of injurious surgical trauma is not clearly defined as stated by a group of investigators in a special article published in 2020 [4].
Neuroinflammation is considered a key pathological process for PND [2, 13]. Surgery may be the major cause to induce neuroinflammation [14]. Different surgeries may have different incidences of PND in humans [6, 7]. One study pooled data of three projects and showed that patients with total hip joint replacement had a lower incidence of PND than patients with coronary artery bypass graft surgery 7 days after surgery but this difference disappeared 3 months after surgery [15]. One animal study showed that rats after cardiac surgery had impairment in more domains of learning and memory than those after abdominal surgery [16]. Of note, animals in that study had 45 min cardiac or intestine ischemia [16], which does not occur clinically. These clinical and laboratory studies may suggest that surgery types are a contributing factor for PND. However, it is not known whether different levels of surgery trauma of the same surgery will lead to different degrees of learning and memory impairment.
The contribution of peripheral macrophages to the development of neuroinflammation after surgery has been reported [17]. The sources of inflammatory cells infiltrated into the brain are not known. Since the spleen stores many immune cells, the spleen may be a source of these cells.
Thus, we hypothesize that the general anesthesia length and surgical trauma severity, two potenti-ally modifiable factors, contribute to PND and that splenocytes are a source of the cells infiltrated into the brain. These hypotheses were tested by subjecting mice to 2- or 6 h isoflurane anesthesia with or without a short (5 min) or long (15 min) surgery. Their learning and memory were evaluated and blood, spleen, and brain samples were harvested to measure proinflammatory cytokines. The migration of splenocytes into the brain was investigated by immunofluorescence staining, labeling splenocytes and examining their appearance in the brain.
METHODS
The experimental protocols were approved by the Institutional Animal Care and Use Committee of the University of Virginia (Charlottesville, VA, USA; protocol number 3114).
Animal grouping
Three-month-old male C57BL/6J mice (young adult) weighing 24–29 g were randomly assigned in a factorial design to the following groups (diagram 1): 1) control group (no anesthesia or surgery); 2) 2 h anesthesia group (isoflurane anesthesia for 2 h); 3) 6 h anesthesia group (isoflurane anesthesia for 6 h); 4) 2 h anesthesia plus short surgery group (5 min surgery to expose left carotid artery plus 2 h isoflurane anesthesia); 5) 6 h anesthesia plus short surgery group (5 min surgery plus 6 h isoflurane anesthesia); 6) 2 h anesthesia plus long surgery group (15 min surgery plus 2 h isoflurane anesthesia); and 7) 6 h anesthesia plus long surgery group (15 min surgery plus 6 h isoflurane anesthesia). Since the long anesthesia duration was three times that of the short anesthesia, the same rule was applied to determine the duration of long surgery. The animals were used for learning and memory tests starting 4 days after the surgery or their tissues were harvested for assays at 6 h, 24 h, 48 h, and 72 h after the surgery.
Schematic presentation of experimental groups.
In another experiment, 3-month-old male C57BL/6J mice were randomly assigned to: 1) control group, and 2) 2 h anesthesia plus long surgery group (diagram 1). These mice were used to harvest hippocampus for immunofluorescent staining.
In the third experiment, 18-month-old male C57BL/6J mice (old mice) weighing 30–36 g were randomly assigned to: 1) control group, 2) 6 h anesthesia group, and 3) 2 h anesthesia plus long surgery group (15 min surgery plus 2 h isoflurane anesthesia). They were subjected to learning and memory tests starting 4 days after the surgery
Anesthesia and surgery
The surgery was left carotid artery exposure [18, 19]. Briefly, mice were anesthetized by 2% isoflurane and kept on spontaneous respiration with a facemask supplied with 100% oxygen. Rectal temperature was maintained at 37°C with a heating blanket (TCAT-2LV, Physitemp Instruments Inc., Clifton, NJ). For short surgery, a 1 cm midline neck incision was made at least 20 min after the onset of isoflurane anesthesia. Soft tissues over the trachea were separated gently. Half-centimeter long left common carotid artery was carefully dissected out from adjacent tissues without damage to the vagus nerve. The wound was then closed by using 4-0 surgical sutures. The surgery was performed under sterile conditions and lasted for 5 min. The total duration of general anesthesia was 2 h or 6 h. For the long surgery, a 2 cm midline neck incision and soft tissue dissection with 1 cm long common artery exposure were performed. The procedure lasted for 15 min. After the surgery, all animals received a subcutaneous injection of 3 mg/kg bupivacaine.
Behavioral testing
Learning and memory were evaluated by novel object recognition and Barnes maze tests. All behavioral tests were conducted at 10:00 am –5:00 pm in a sound-isolated room.
Novel object recognition test
As described previously [20], mice were put in an open-field chamber for 5 min for habituation 4 days after surgery. Two of the same objects were placed at adjacent angles of the chamber on the learning day (post-surgery day 5). Mice were put into the chamber with their backs turned towards the objects and allowed to explore the chamber freely for 5 min. An animal was eliminated if the total exploration time on two objects was less than 5 s. One object was replaced by a novel object 30 s or 24 h later. The mouse was put into the chamber and allowed to explore for 5 min. Animal behavior was recorded by ANY-maze behavioral tracking software (Stoelting Co., IL). Exploratory time of new (T2) and old (T1) objects within 5 min was recorded and the memorization ability of the mouse was quantified by discrimination index (DI): DI = T2/(T1 + T2). The DIs at 30 s and 24 h after the training reflected the instant and long-term memory, respectively [21].
Barnes maze
Seven days after surgery, animals were subjected to the Barnes maze test as previously described [22, 23]. Barnes maze is a circular platform with 20 equally spaced holes (SD Instruments, San Diego, CA). One hole was connected to a dark chamber that was called the target box. The test started by placing animals in the middle of the Barnes maze. Aversive noise (85 dB) and bright light (200 W) shed on the platform were used to encourage mice to find this box. Mice were subjected to four 3 min training sessions with a 15 min inter-session interval on each training day. After training for 4 days, their reference memory was tested on day 5 and day 12. No test was performed during the period from day 5 to day 12. The latency to enter the target box during each trial was recorded by an ANY-Maze video tracking system (SD Instruments).
Spleen weight
The spleen was harvested from each mouse and weighed at 6 h, 24 h, 48 h, and 72 h after surgery or anesthesia. The spleen weight was normalized to the body weight of the mouse.
Splenocyte isolation and Cy5-labeled splenocyte injection
Mice were euthanized by deep isoflurane anesthe-sia. Mouse spleen was removed under sterile condi-tions and sliced into small pieces in cell culture medium. The fragments of the spleen were placed onto a 40μm cell strainer attached to a 50 ml conical tube and then pressed through the strainer with excess cell culture medium using the plunger end of a syringe. The cell suspension was centrifuged at 1,000 rpm for 5 min at room temperature. The cell pellet was re-suspended in 2.0 ml ammonium-chlo-ride-potassium lysis buffer and incubated for 15 min to remove red blood cells. After centrifugation at 1,000 rpm for 5 min at room temperature, the cells were re-suspended in cell culture medium. They were incubated with cyanine 5 (Cy5, catalog number: 5436, R&D Systems) for 30 min and washed 3 times by phosphate buffered saline (PBS). About 1.0 x 105 of Cy5-labeled splenocytes were injected into the tail vein of each animal at the end of anesthesia. These mice were euthanized at 6 h, 24 h, and 48 h after the surgery. Their hippocampus was harvested for detecting Cy5 signal.
Serum, brain, and spleen harvesting
Mouse blood was collected using cardiac puncture under deep isoflurane anesthesia at 6 h, 24 h, 48 h, or 72 h after anesthesia, kept at room temperature for 4 h, and centrifuged at 1300 rpm for 20 min at 4°C. Serum was collected and kept at –80°C. The mouse was perfused transcardially with ice cold saline after blood was collected. Brain and spleen were harvested. These samples were used for ELISA analysis. Whole brain at Bregma –3 to –6 mm was used for immunofluorescent staining.
Immunofluorescent staining
The immunofluorescent labeling and quantification of the staining were performed as we have described before [16, 17]. Briefly, coronal 20μm (for CD68 and cy5 images) or 60μm [for quantification of ionized calcium binding adaptor molecule 1 (Iba-1)] thick sections were cut from Bregma –3 to –6 mm. After being washed in Tris-buffered saline (TBS), sections were blocked in 10% donkey serum plus 1% bovine serum albumin in TBS containing 0.3% triton-x 100 for 2 h at room temperature and then incubated at 4°C overnight with the following primary antibodies: mouse monoclonal anti-CD68 antibody (1:100 dilution, catalog number: ab955, Ab-cam) and rabbit monoclonal anti-Iba-1 antibody (1:200 dilution; catalog number: 019–19741, Wako Chemical Co., Richmond, VA). The donkey anti-mouse IgG antibody conjugated with Alexa Fluor 488 (1:200 dilution, catalog number: A21202; Invitrogen) or donkey anti-rabbit IgG antibody conjugated with Alexa Fluor 594 (1:200 dilution, catalog number: A21207; Invitrogen) were incubated with the sections for 1 h at room temperature in the dark. The sections were incubated with Hoechst 33342 (Thermo Scientific) and mounted with Vectashield mounting medium. Staining images were acquired by z-stack with an LSM710 microscopy system. To quantify the staining, the whole dentate gyrus region that was covered by 2-3 non-overlapping fields from each of six sequential sections of one mouse was imaged. The number of pixels per image with intensity above a predetermined threshold level was considered as a positively stained area for a protein and quantified using the Image-pro plus 6.0 (Media Cybernetics, Inc., Rockville, MD) and presented as integrated optical density and percentage of positive area in the total area. The 6 measurements per mouse were averaged to reflect the level of positive staining. All quantitative analyses were performed in a blinded manner.
ELISA analysis
Interleukin (IL)-1β and IL-6 levels in the spleen, serum and hippocampus of mice were determined by using ELISA kits (catalog number: SMLB00C for IL-1β catalog number: SM6000B for IL-6; R&D Systems) according to the manufacturer’s instructions and as we described previously [16, 17]. The detection threshold of the kits for IL-6 and IL-1β was 1.8 pg/ml and 4.8 pg/ml, respectively. The amount of IL-1β and IL-6 in each sample was normalized by its protein content.
Statistical analysis
Parametric results in normal distribution are presented as means±S.E.M. Non-normally distributed data are in a box plot. Data from Barnes maze training sessions were analyzed by two-way repeated measures analysis of variance (ANOVA) followed by Tukey’s test. For young adult mice, two-way ANOVA was used to determine whether anesthesia and surgery were significant factors in determining the performance of mice in the memory phase of the Barnes maze test and novel object recognition test and their spleen weights and the concentrations of IL-1β and IL-6 in the serum, spleen, and hippocampus. Two factors (anesthesia and surgery), two anesthesia lengths (2 h and 6 h), and three surgery levels (no surgery, short surgery, and long surgery) were considered in these analyses. Additionally, these data and the data of old mice were analyzed by one-way ANOVA followed by Tukey’s test for normally distributed data or by one-way analysis of variance on rank followed by the Tukey test for non-normally distributed data. A significant difference was defined as p < 0.05. Statistical analyses were performed with SigmaStat (Systat Software, Inc., Point Richmond, CA).
RESULTS
Long surgery induced learning and memory dysfunction in young adult mice
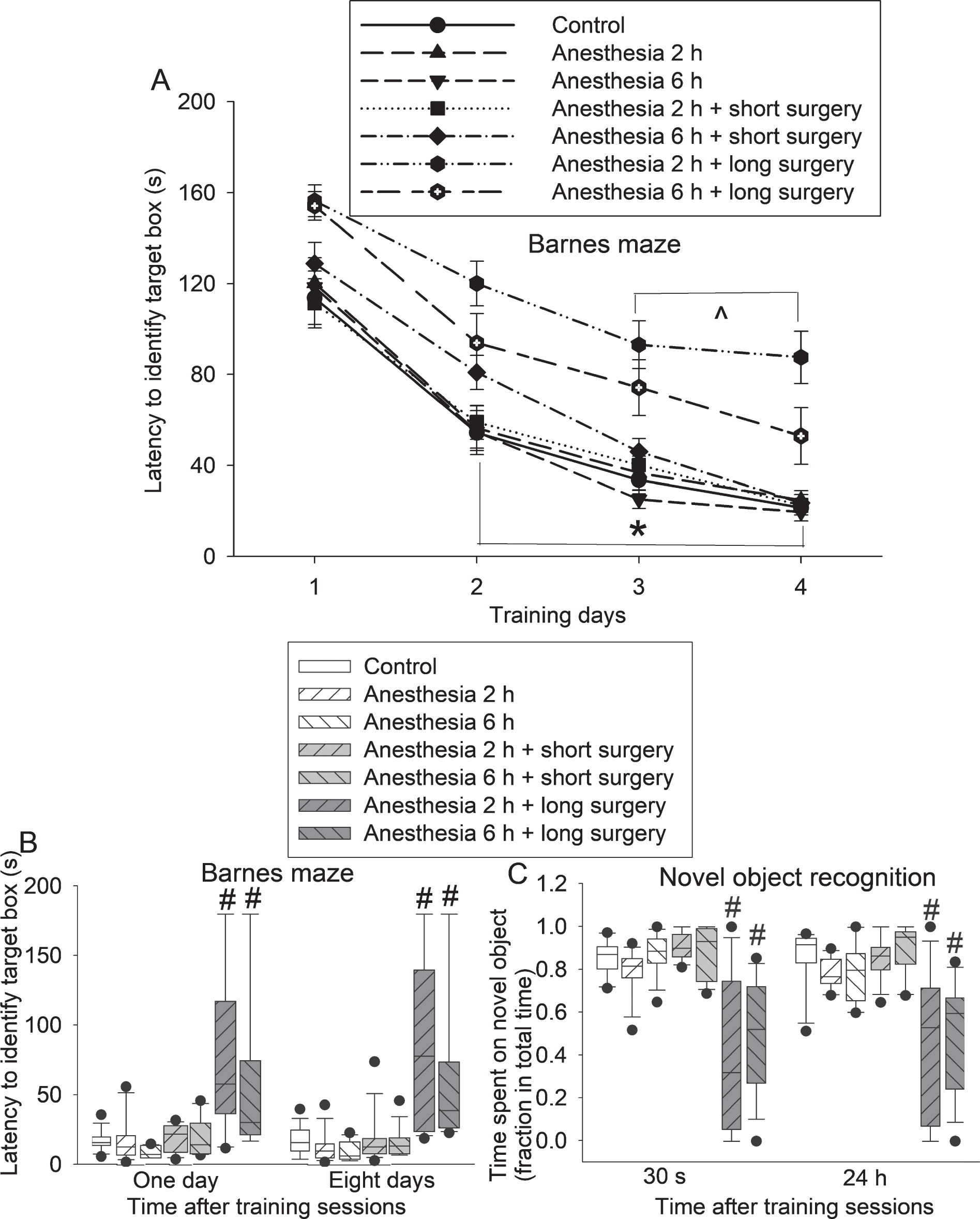
The time for young adult mice in all 7 groups to identify the target box in the Barnes maze test was decreased with increased training sessions (Fig. 1A). Anesthesia (both 2 h and 6 h) was not a factor to affect the time needed to identify the target box in the training sessions of Barnes maze test [F(2,98) = 2.380, p = 0.098]. Surgery was a significant factor to affect the time needed to identify the target box [F(2,98) = 31.691, p < 0.001]. Additional two-way repeated measures ANOVA analyses showed that anesthesia alone or anesthesia plus short surgery was not a factor to affect the time needed to identify the target box in the training sessions of Barnes maze test (all ps>0.05 for the 4 comparisons: control versus 2 h anesthesia, control versus 6 h anesthesia, control versus 2 h anesthesia plus short surgery, control versus 6 h anesthesia plus short surgery). However, long surgery was a significant factor to affect this time [F(1,27) = 35.309, p < 0.001 for comparison between control and 2 h anesthesia plus long surgery; F(1,27) = 11.012, p = 0.003 for comparison between control and 6 h anesthesia plus long surgery]. The times needed to identify the target box for those two groups of mice with long surgery were also longer than those of control mice 1 and 8 days after the training sessions of Barnes maze test (Fig. 1B). Again, surgery but not anesthesia was a significant factor to affect this time [for example, one day after the training sessions: F(2,82) = 22.004, p < 0.001 for surgery; F(1,82) = 1.459, p = 0.231 for anesthesia]. There was no interaction between anesthesia and surgery [for example, one day after the training sessions: F(2,80) = 0.629, p = 0.535]. The longer time needed for surgical mice to identify the target box may not be due to the impairment of motor function because mice with this surgery do not appear to have motor function impairment [24]. Thus, our results suggest that mice with long surgery have spatial learning and memory dysfunction.
Learning and memory dysfunction after a long surgery in young adult mice. Mice were subjected to various experimental conditions and tested in the novel object recognition paradigm 4 days after surgery and the Barnes maze 7 days after surgery. A) Performance in the training sessions of Barnes maze test. B) Performance in the memory phase of Barnes maze test. C) Performance in novel object recognition paradigm. Results are mean±S.E.M. (n = 14–15 for panels A and B, = 11–13 for panel C). *p < 0.05 for the comparisons of values of day 1 with those of day 2, values of day 1 with those of day 3, and values of day 1 with those of day 4 for the control group, 2 h anesthesia group, 6 h anesthesia group, 2 h anesthesia plus short surgery group, and 6 h anesthesia plus short surgery group. ∧p < 0.05 for the comparisons of values of day 1 with those of day 3 and values of day 1 with those of day 4 for the 2 h anesthesia plus long surgery group and 6 h anesthesia plus long surgery group. #p < 0.05 compared with control group.
Mice with long surgery spent less time with novel object than control mice no matter whether this novel object recognition test was performed 30 s or 24 h after the training session (Fig. 1C). Surgery but not anesthesia was a significant factor to affect this time [for example, 30 s after the training sessions: F(2,71) = 38.205, p < 0.001 for surgery; F(1,71) = 1.089, p = 0.300 for anesthesia]. There was no interaction between anesthesia and surgery [for example, 30 s after the training sessions: F(2,69) = 0.746, p = 0.478]. These results suggest that mice with long surgery have dysfunction of non-spatial learning and memory.
Surgery induced systemic and neural inflammation in young adult mice
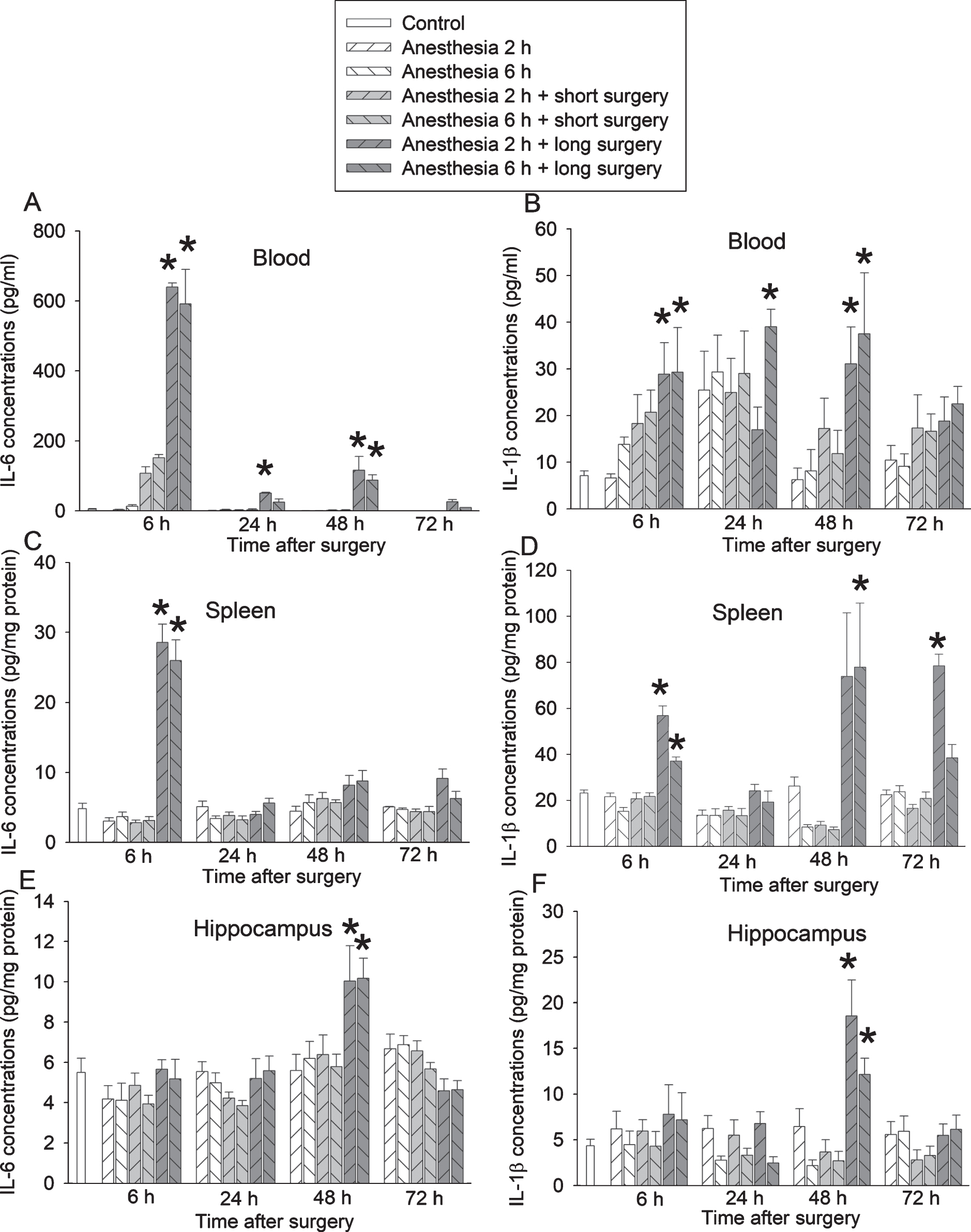
Mice with long surgery had increased IL-6 and IL-1β in the blood, spleen, and hippocampus. The increase of IL-6 and IL-1β in the blood and spleen started 6 h after surgery (Figs. 2A-D). This increase in the hippocampus was 24 h after surgery (Figs. 2E, F). Surgery but not anesthesia was a significant factor to increase the proinflammatory cytokines [for example, IL-6 increase in the blood 6 h after surgery: F(2,44) = 140.447, p < 0.001 for surgery; F(1,44) = 0.161, p = 0.690 for anesthesia]. There was no interaction between anesthesia and surgery [for example, IL-6 increase in the blood 6 h after surgery: F(2,42) = 0.565, p = 0.573]. These results suggest that long surgery increases proinflammatory cytokines in the blood, spleen, and brain. As a control procedure, we determined the internal measurement variations of the samples that were measured in duplicate by ELISA kits. The Pearson correlation coefficient between these duplicates was 0.999 (n = 28) and 0.987 (n = 28), respectively, for IL-6 and IL-1β measurements. The difference between the duplicates was 0.29±0.26% and 4.82±0.68%. These results suggest that the internal variations of measurements of samples are very small.
Proinflammatory cytokine increase after a long surgery in young adult mice. Mice were subjected to various experimental conditions. Their blood, spleen and hippocampus were harvested at various times after surgery. A) IL-6 in blood. B) IL-1β in blood. C) IL-6 in spleen. D) IL-1β in spleen. E) IL-6 in hippocampus. F) IL-1β in hippocampus. Results are mean±S.E.M. (n = 8). *p < 0.05 compared with control group.
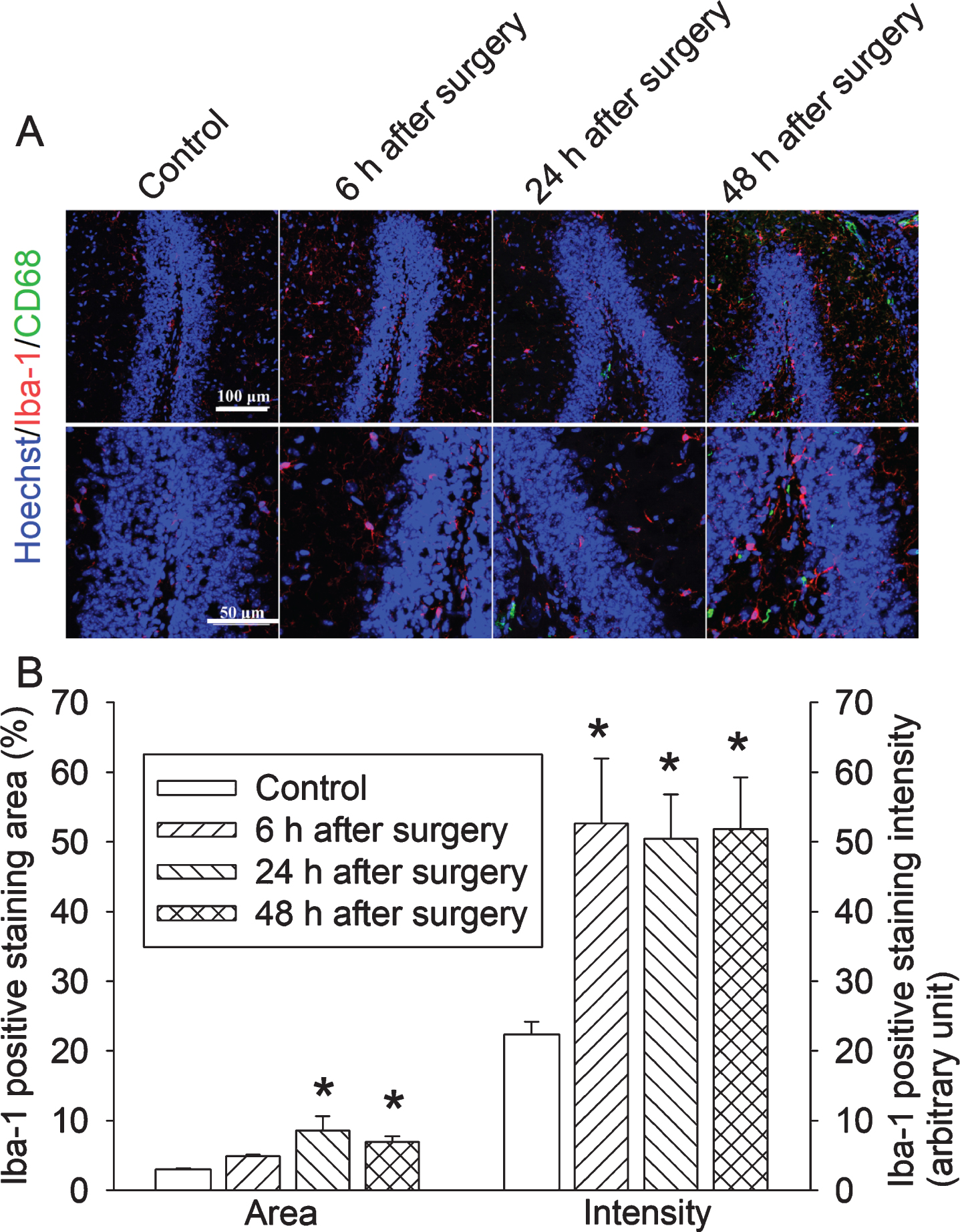
To determine whether surgery affects the expression of Iba-1, a marker for microglia [18, 19], we studied mice with 2 h anesthesia plus long surgery. Iba-1 expression in the hippocampus started to be increased at 6 h after the surgery (Fig. 3). This re-sult and the increased proinflammatory cytokines in the hippocampus suggest that surgery induces neuroinflammation. Interestingly, some cells in the hippocampus were positively stained for CD68 (Fig. 3), a marker for peripheral monocytes [17], mostly at 24 h after surgery. This result suggests the infiltration of peripheral monocytes into the brain.
Increased Iba-1 expression after a long surgery in young adult mice. Mice were subjected to 2 h anesthesia plus long surgery. There hippocampus was harvested for immunofluorescent staining. A) Representative images of immunofluorescent staining. Scale bar = 100μm in the up panel, = 50μm in the lower panel. B) Quantification of immunofluorescent staining. Results are mean±S.E.M. (n = 6). *p < 0.05 compared with control group.
Surgery reduced spleen weights and induced splenocyte migration into the brain in young adult mice
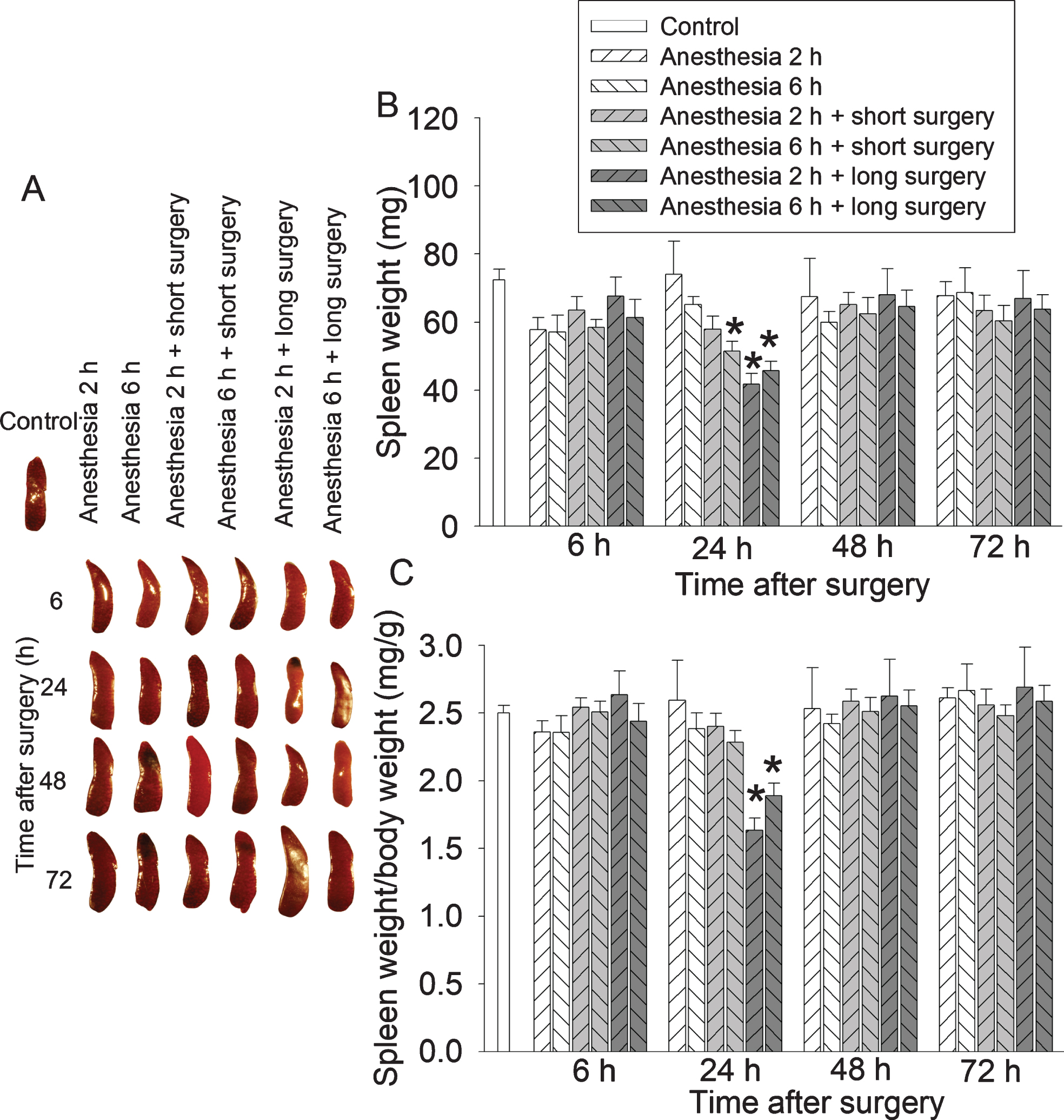
Long surgery decreased the spleen weights 24 h after surgery (Fig. 4). Surgery but not anesthesia was a significant factor to reduce spleen weights [for example, the ratio of spleen weights/body weights 24 h after surgery: F(2,44) = 13.213, p < 0.001 for surgery; F(1,44) = 0.040, p = 0.841 for anesthesia]. There was no interaction between anesthesia and surgery [for example, the ratio of spleen weights/body weights 24 h after surgery: F(2,42) = 1.361, p = 0.267].
Spleen weight decrease after a long surgery in young adult mice. Mice were subjected to various experimental conditions. Their spleen was harvested and weighed at various times after surgery. A) Representative images of spleens. B) Spleen weights. C) Ratio of spleen weights/body weights. Results are mean±S.E.M. (n = 8). *p < 0.05 compared with control group.
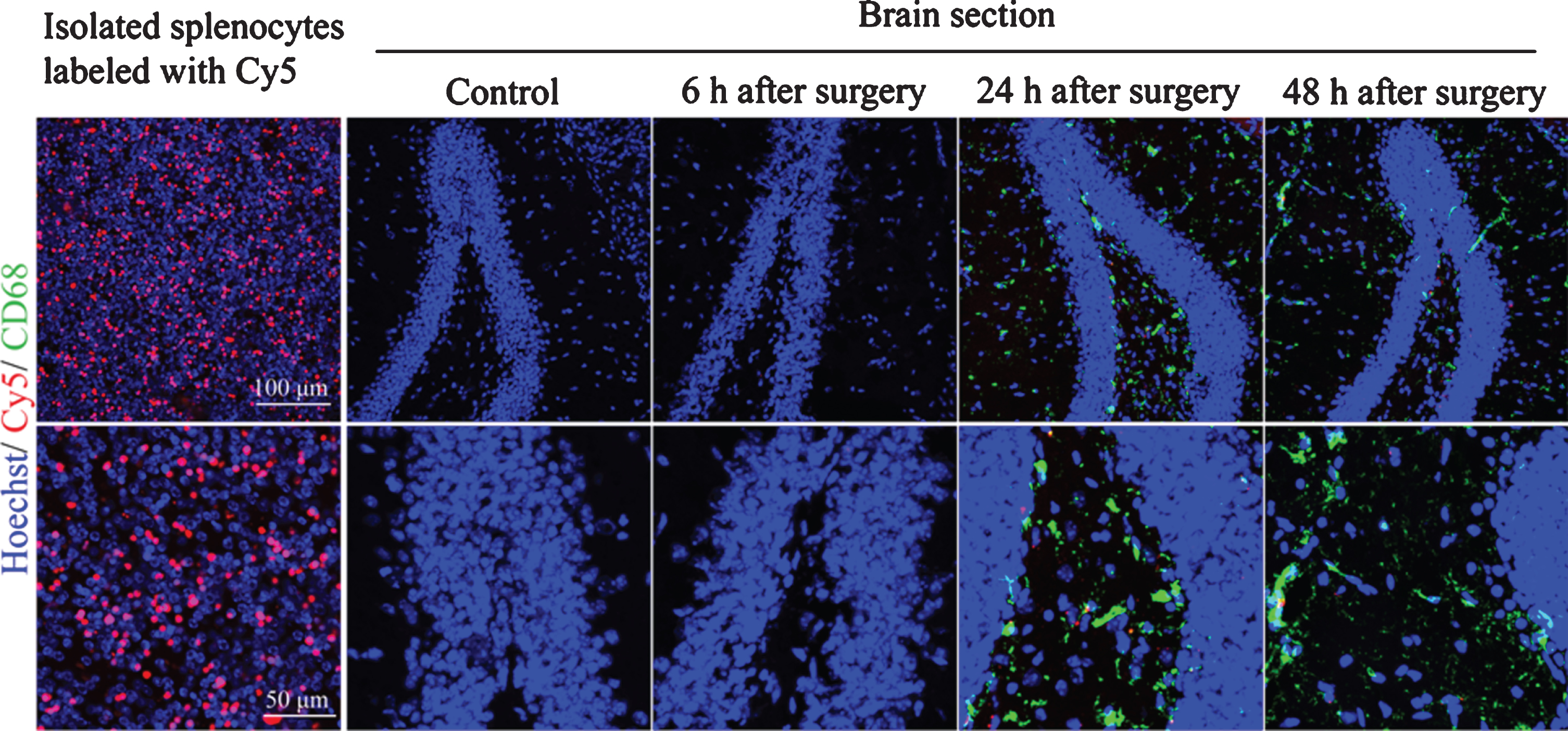
To determine whether splenocytes could infiltrate into the brain, cy5 labeled splenocytes were injected intravenously into mice with 2 h anesthesia plus long surgery at the end of anesthesia. There were cy5 positive cells in the hippocampus of these mice 24 h after the surgery. Some cy5 positive labeling was co-localized with CD68 labeling (Fig. 5), suggesting that splenocytes infiltrate into brain after surgery.

Infiltration of splenocytes into hippocampus in young adult mice. Mice were subjected to 2 h anesthesia plus long surgery. Their hippocampus was harvested for immunofluorescent staining. Splenocytes were isolated from the control mice (without being exposed to anesthesia and surgery before spleen harvesting) and labeled in vitro with cy5. These labeled cells were injected into control mice (no anesthesia and surgery) or mice with surgery at the end of surgery and anesthesia.
Surgery induced learning and memory impairment in old mice
The time for old mice to identify the target box was decreased with increased training in the Barnes maze test (Fig. 6A). Surgery but not anesthesia was a main factor to affect the time for mice to iden-tify the target box during the training sessions [F(1,16) = 8.517, p = 0.010 for comparison between control and 2 h anesthesia plus long surgery; F(1,16) = 0.361, p = 0.556 for comparison between control and 6 h anesthesia]. Mice in the surgery group but not in the anesthesia alone group took longer than control mice to identify the target box 1 day or 8 days after the training sessions in the Barnes maze test (Fig. 6B). Similarly, mice with surgery but not those with anesthesia alone spent less time with the novel object than control mice at 30 s or 24 h after the training sessions in the novel object recognition tests (Fig. 6C).

Learning and memory dysfunction after a long surgery in old mice. Old mice were subjected to various experimental conditions and tested in the novel object recognition paradigm 4 days after surgery and the Barnes maze 7 days after surgery. A) Performance in the training sessions of Barnes maze test. B) Performance in the memory phase of Barnes maze test. C) Performance in novel object recognition paradigm. Results are mean±S.E.M. (n = 9). *p < 0.05 for the comparisons of values of day 1 with those of day 2, values of day 1 with those of day 3 and values of day 1 with those of day 4 for all groups. #p < 0.05 compared with control group.
To evaluate if aging was a factor for the worsened learning and memory after anesthesia and surgery, we analyzed the data of the control group and 2 h anesthesia plus long surgery group of young adult and old mice by two-way repeated measures ANOVA (for Barnes maze training data) or two-way ANOVA (age and surgery as the two factors). Age was not a significant factor for determining the time to identify the target box in the training and memory phases of Barnes maze tests. Age was not a factor to determine the performance of mice in the novel object recognition test but the p values were small [F(1,37) = 2.461, p = 0.125 and F(1,38) = 2.697, p = 0.109 for the data at 30 s and 24 h, respectively, after the training sessions].
DISCUSSION
Our results from young adult mice suggest no effects of anesthesia on PND development no matter whether anesthesia alone, anesthesia plus short surgery, or anesthesia plus long surgery condition was considered. However, the long surgery is a significant factor to affect learning and memory. Similarly, old mice with long surgery but not with long anesthesia alone had impaired learning and memory. Since age has been consistently identified as a risk factor for PND in humans [2–4], studying old mice is an approach to indicate potential usefulness of the findings in understanding the pathophysiological changes of PND. Our results of young adult and old mice indicate that surgical trauma but not anesthesia is a critical factor for the development of PND. Of note, a possible difference in PND incidences after various surgeries has been reported [6, 25]. However, direct comparison of different degrees of surgical trauma of the same procedure has not been reported. We used two different degrees of surgical trauma. Mice with long surgery had a longer incision (2 cm versus 1 cm skin incision) and more tissue dissection (dissecting out 1 cm versus 0.5 cm long carotid artery) and tissue manipulation (5 min versus 15 min) than mice with short surgery. Also, mice with long surgery had higher proinflammatory cytokine levels than mice with short surgery, an indirect sign of more severe surgical trauma in mice with long surgery. Our results provide evidence to suggest a novel idea that surgical trauma severity of the same surgery may be a significant contributing factor for the development of PND.
PND is more common in elderly patients than younger adults [2–4]. Consistent with this clinical phenomenon, our previous study has shown that more mice in the old mouse group have learning and memory decline than those in the young mouse group after anesthesia and surgery when individual mouse-based assessment instead of the commonly used group-based assessment is used to identify the individuals with learning and memory decline [24]. A previous study has shown that partial hepatectomy induces learning and memory dysfunction in 18-month-old but not in 2-month-old female rats (group-based assessment) [26]. Our current study does not provide evidence to support the idea that old mice had worsened learning and memory impairment than young adult mice. However, one will need to be cautious about this conclusion. First, we did not perform the assessment of learning and memory of old and young adult mice in the same set of experiment. They were in different sets of experiments because comparing the learning and memory decline between old and young mice was not the major goal of this study. Thus, it may not be appropriate to perform this comparison. Second, we used group-based assessment and did not measure the learning and memory of each mouse at baseline and, therefore, it is not possible to measure the decline of learning and memory for each mouse. Third, the p values for age as a factor to influence the performance in novel object recognition test were small. Increasing the sample size may decrease them to less than 0.05, which will suggest a different conclusion.
Neuroinflammation may be a critical neuropathological process for PND [14, 19]. Consistent with this theory, neuroinflammation occurred in mice after a long surgery but not in mice exposed to anesthesia alone or anesthesia plus short surgery. IL-6 has been suggested to be necessary for PND in an experiment using young adult mice [27]. Our study showed that long surgery but not short surgery or anesthesia alone increased IL-6 in the blood, spleen, and brain. Neuroinflammation can impair neurogenesis and synaptic function and induce cell injury or death [22, 29]. For example, IL-6 blocks the trafficking of glutamate receptors to cell plasma membrane, a biochemical process for learning and memory [30]. These neuroinflammatory effects ultimately lead to cognitive dysfunction after surgery.
We and others have suggested that neuroinflammation may be transmitted from systemic inflammation after a surgery via disrupted blood-brain barrier [14, 29]. The finding that the increase of proinflammatory cytokines in the blood and spleen was earlier than that in the hippocampus supports this suggestion. The transmission of systemic inflammation to the brain may be helped by permeation of proinflammatory factors or cells into the brain [15, 17]. Our results provide evidence to suggest that spleen may be an important storage organ for those factors or cells because the spleen weights were decreased in the mice with long surgery and splenocytes were found in the hippocampus. The spleen is known to store a large amount of proinflammatory cells [31]. Although a spleen weight decrease can be caused by many etiologies, this decrease in mice with a surgery that has very limited blood loss may suggest that splenocytes move away from the spleen. This result and the finding of splenocytes in the brain provide initial evidence that splenocytes may migrate into the brain after surgery. It is not known how important these migrated splenocytes are in PND development. Nevertheless, the role of invasion of peripheral cells into the brain in PND has been suggested [17]. Thus, it is possible that splenocytes may play a role in PND. Interestingly, splenectomy has been an animal model for PND study [32]. In addition to the spleen, many other organs and tissues, such as blood and liver, have immune cells. Thus, animals with splenectomy can develop inflammatory responses and PND [32]. Nevertheless, based on the fact that the spleen stores many immune cells [31], the knowledge that inflammation is an important pathological process for PND and our finding that splenocytes are a source of cells migrating into the brain after surgery, splenectomy may not be a good surgical model for studying PND.
Our study clearly showed that anesthesia does not contribute to PND. Many studies have shown that anesthetic induces learning and memory impairment in animals [4, 33]. The reasons for the different findings between our study and some previous studies are not clear but may be related to different strains of animals and methods used to assess learning and memory. We also carefully maintained the temperature of animals under anesthesia in this study. Also, many previous studies examine learning and memory of animals within 1 to 3 days after the exposure [4, 17]. Our study tests the learning and memory of animals much later. Consistent with our study, some previous studies have shown no learning and memory dysfunction after anesthetic exposure [10, 34]. Similar to the animal studies, patients having general anesthesia had a worse cognition function than patients having regional anesthesia at 1 week after surgery in one study [35], but this difference was not revealed in another study [36]. Also, there was no difference in cognitive functions assessed at 3 months or 6 months after surgery between patients receiving general anesthesia or regional anesthesia with sedation [35–37]. Of note, we used a very long anesthesia duration (6 h). A previous study indicates that isoflurane anesthesia (2%) may not induce significant acidosis and CO2 accumulation [38]. The lack of learning and memory impairment in those mice anesthetized by isoflurane for 6 h indicates minimal influence of potential physiological disturbance by isoflurane anesthesia in these brain functions.
Our study has limitations. We showed that splenocytes migrated into the brain and contributed to the production of proinflammatory cytokine. However, we have not determined the role of these splenocytes in neuroinflammation and PND. To determine this role, we will need to specifically prevent the migration of splenocytes into the brain. We do not know any specific methods that can do that. Clodrolip can almost deplete bone marrow-derived macrophages in the spleen [39]. A previous study has indicated a role of these macrophages in the memory deficit after surgery in mice [39]. However, the spleen also contains other cells, such as lymphocytes [31]. Since almost all splenocytes are immune cells, it can be assumed that these splenocytes that migrate into brain will have modulating effects on the neuroinflammation induced by surgery. Another limitation of our study is that we only tested isoflurane. Extrapolation of this finding to other anesthetics needs to be cautious. However, isoflurane is used clinically and currently there is no evidence to suggest that other anesthetics will be worse than isoflurane [19]. Finally, since old mice is a rare resource, we did not perform the full set of experiments in old mice as we did in 3-month-old mice. However, old mice did not have learning and memory impairment after 6 h anesthesia and had learning and memory impairment after 2 h anesthesia plus 15 min surgery, suggesting a major role of surgery/surgical trauma in learning and memory impairment. These results capture the main findings of studies of 3-month-old mice.
CONCLUSIONS
Our results suggest that the degree of surgical trauma is a major factor to affect the development of PND and neuroinflammation. Splenocytes may contribute to this process. Anesthetics/general anesthesia may not contribute significantly to postoperative neuroinflammation and cognitive dysfunction.
Footnotes
ACKNOWLEDGMENTS
This study was supported by grants (R01 HD089999, RF1 AG061047, and R01 NS099118 to Z. Zuo) from the National Institutes of Health, Bethesda, MD, the Robert M. Epstein Professorship endowment (to Z. Zuo), University of Virginia, Charlottesville, VA, grants from the National Natural Science Foundation of China, Beijing (Grant number: 81641038 and 81870226 to Z Lai) and a grant from the Joint Funds for the innovation of science and Technology of Fujian Province, Fuzhou (Grant number: 2018Y9042 to Z Lai).