
Abstract
Background:
Alzheimer’s disease (AD) is the most general, chronic, and progressive neurodegenerative senile disorder characterized clinically by progressive cognitive deterioration and memory impairment. Neoline is effective against neuropathic pain models, but the effects of neoline against AD-like phenotypes have not been investigated.
Objective:
We offer the investigation of the effects of neoline in AD.
Methods:
In this study, a Tg-APPswe/PS1dE9 AD mouse model was treated orally with neoline at a concentration of 0.5 mg/kg or 0.1 mg/kg starting at 7.5 months and administered for three months, and its anti-AD effects were evaluated.
Results:
Neoline improved memory and cognition impairments and reduced the number of amyloid-beta plaque and the amount of amyloid-β in the brain of AD mice. Furthermore, neoline reduced the anxiety behavior in the AD mouse model. The chronic administration of neoline also induced AMPK phosphorylation and decreased tau, amyloid-β, and BACE1 expression in the hippocampus. These findings indicate that chronic administration of neoline has therapeutic effects via AMPK activation, and BACE1 downregulation resulted in a decrease in the amyloid-β levels in the brain of Tg-APPswe/PS1dE9 AD mice.
Conclusion:
Our results suggest that neoline is a therapeutic agent for the cure of neurodegenerative diseases like AD.
INTRODUCTION
Alzheimer’s disease (AD) is the most general, chronic, and progressive neurodegenerative senile disorder, affecting approximately 7%of individuals aged 65 and above and clinically characterized by progressive cognitive deterioration and memory impairment. The neuropathological hallmarks of AD are the deposition of extracellular amyloid-β (Aβ) proteins and intracellular neurofibrillary tangles composed of hyperphosphorylated tau proteins in the hippocampus and cerebral cortex regions of the brain, resulting in neuronal loss [1, 2]. Aβ peptides are generated primarily from the cleavage of the transmembrane glycoprotein amyloid-β protein precursor (AβPP) by the enzymes, β-secretase (BACE-1) and γ-secretase [3]. High BACE1 enzymatic activity has been found in human AD brain extracts, which is consistent with the finding that neurons produce the highest levels of Aβ [4, 5]. Moreover, in the postnatal brain of AD mice, the highest BACE1 protein level in neuritic dystrophies in the vicinity of Aβ plaques was reported [6–8], and BACE1 has been demonstrated to be involved directly in Aβ production based on the data from several knockout (KO) mouse models [7–9]. Therefore, BACE1 is believed to be essential for Aβ production and has been pharmacologically targeted to reduce Aβ production.
Dysregulated inflammation of the central nervous system (CNS) may induce neurodegeneration. Ade-nosine monophosphate-activated protein kinase (AMPK), a highly conserved serine/threonine-pro-tein kinase, serves as a sensor of the cellular energy status and is a regulator of the energy balance and controls many cellular processes, such as autophagy, cell death, and inflammation, to maintain cellular homeostasis [10]. AMPK dysregulation was associated with chronic neurodegenerative diseases, like multiple sclerosis, AD, Parkinson’s disease, Huntington’s disease, amyotrophic lateral sclerosis, and neuropathies [10]. AMPK also regulates autophagy for the clearance of Aβ in AD. Therefore, Aβ clearance promotion is also regarded as another valuable strategy in anti-AD research because a failure of Aβ clearance is strongly associated with AD pathogenesis [11, 12].
Despite the serious health problems caused by AD, only five drugs, including two classes of drugs, have been approved for AD. These treatments serve to control symptoms rather than alter the course of the disease [13]. Additionally, relatively few clinical trials have been undertaken in AD over the last decade, and these have had a 99.6%failure rate [14]. As a result, innovation in synthetic new drugs or the discovery of natural drugs to treat AD is a challenge for basic science and clinical medicine.
The progressed lateral root of Aconitum carmich-aelii Debeaux (Ranunculaceae) is an extensively used traditional Chinese medicine that is known as Fuzi in China. The root was used to cure acute my-ocardial infarction, low blood pressure, coronary heart disease, chronic heart failure, and pain associ-ated with coldness. Fuzi, with its active compounds, possesses wide-reaching biological activities, including effects on the cardiovascular system, anti-infl-ammation and analgesic action, anti-tumor activity, development of immune system, hypoglycemic and hypolipidemic effects, anti-aging effects, effects of protecting kidney, and effects on energy metabolism. Among them, neoline is effective against neuropathic pain models induced by paclitaxel [15, 16] and partial ligation of the sciatic nerve [17]. On the other hand, the effects of neoline against AD-like phenotypes have not been investigated. APP/PS1 is a dual transgenic mouse expressing chimeric mouse/human amyloid precursor protein (Mo/HuAPP695swe) and mutant human presenilin 1 (PS1-dE9) were directed to CNS neurons. Both mutations were associated with early-onset AD. Therefore, AD symptoms, such as amyloid plaque formation and memory loss, can appear as early as 6 months compared to other AD animal models. So, this study examined the anti-AD effects of neoline in the Tg-APPswe/PS1dE9 transgenic mouse model.
METHODS
AD mouse model
This study was approved by the Animal Care and Ethical Committee of Dongguk University (Cli-nical trial approval number IACUC-2019-017-2). All animal handling procedures followed the instructions of the Institute for Laboratory Animal Research of the Dongguk University. Tg-APPswe/PS1dE9 transgenic mice indicating human mutated APP and PS1 (APPswe/PS1dE9) were originally purchased from the Jackson Laboratory Order number: (JAX# 000664) (Bar Harbor, ME, USA) and trained to maintain a C57BL6×C3H F1 hybrid, as described previously [18]. The animals were accommodated under laboratory settings at a restrained temperature (20±2°C) and sustained under 12 h light-dark cycles (light from AM 7 to PM 19 h) with food and water available ad libitum. Tg-APPswe/PS1dE9 males mice at 7.5 months of age were randomized into the control (Normal, n = 7), neoline (Chengdu Biopurify Phytochemicals Ltd, China. purity ≥98%) (Neoline_L, 0.1 mg/kg/day, n = 7; Neoline_H, 0.5 mg/kg/day, n = 7), and donepezil hydrochloride (Eisai Korea, Inc.) (Donepezil, 1 mg/kg/day, n = 7) groups. Lab chow containing neoline or donepezil was prepared and fed for three months beginning at 7.5 months of age. Previously, we have measured the average amount of feed consumed by male mouse per day. Male mouse consumed an average of 4 g per day. Therefore, neoline was prepared and administered to feed at a concentration of 0.1 mg/kg or 0.5 mg/kg and 1 mg/kg of donepezil. After the experiment began, weight and feed intake amount were measured once a week (Supplementary Figure 1).
Step-through passive-avoidance test
The equipment (AP model; O’Hara Co., Tokyo, Japan) for the step-through passive avoidance test contained two divisions, illuminated division [10 cm×12 cm×10 cm; light at the top of the divi-sion (27 W, 3000 lx)] and dark compartment (10 cm×17 cm×10 cm). A guillotine door separated the compartments. During the training stage, a mouse was placed in the illuminated intact chamber. As the chamber was lit, the mouse was go downward to the opened guillotine door into the dark compartment. The time lost in the illuminated chamber was characterized as the latency time. Once the mouse reached the dark chamber after three seconds, a foot agitate (0.3 mA, 50 V, 50 Hz ac, for 3 s) was supported to the floor grids in the dark chamber. The mouse could only escape from the agitate by moving backward to the protected, illuminated chamber. During the training stage, such procuration trials were transferred out at 7.5 months of age. This was evaluated as training avoidance from foot-impact, if the mouse was continued in the illuminated compartment for 300 s after being fixed there. The retention trials were moved out at 24 h after the training trial to calculate the detainment of avoidance memory. The latency time was reached for up to 300 s without conveying a foot shock. The mouse retained the avoidance memory when it remained in the illuminated safe compartment for 300 s.
Morris water maze test
This behavioral test, which involved a maze test, was performed at 7.5 months of age, and the data were analyzed. The water was formed milky by white ink, and the temperature of the water was maintained at 22∼25 C. The diameter and height of the water were 100 cm and 35 cm, respectively. A transparent acrylic plastic platform was immersed 1 cm under the water. During training, the test subjects were placed in the water and allowed 120 s to search for the platform. The mice were allowed 10 s to remain on the platform and then returned to the home cage. A video camera was seated on the ceiling above the pool and connected to a video recorder and tracking device (Ethovision XT, Noldus, VA, USA), allocating on and offline motorized recording of the path appropriated by the mouse.
Open field test
For the open field test, individual mice were placed in 42 cm×42 cm×25 cm acrylic animal cages for 5 min, during which their horizontal and vertical movements were monitored. The software was then used to calculate, in both the horizontal plane, the total number of movements, the distance traveled, and the time spent moving for each mouse.
Y maze spontaneous alternation test
The Y-maze contains three identical arms seated in the shape of a “Y.” Each arm was 15 inches long and 3.5 inches wide with 3 inch high walls formed from black painted wood. Each mouse was placed at the midpoint of the apparatus and permitted to carry openly over the maze for 7 min. The series of arm entries was recorded visually. Interchange was characterized as consecutive entry into the three arms on the overlapping triplet sets. The interchange (%) was calculated as the ratio of actual variation to possible variations (determined as the amount of mobilized entries minus two) multiplied by 100.
Aβ42 quantification by enzyme-linked immunosorbent assay (ELISA)
The cortex was removed immediately and frozen. The tissues were stored at –80°C prior to use. Particular brain was pasteurized in 8 vol (wt./vol) of cold 5 M guanidine, Tris–HCl HCl/50 mM, and blended at room temp for 4 h. The passages were diluted in Dulbecco’s phosphate-buffered saline having 5%bovine serum albumin (BSA) and 0.03%Tween 20 (pH 7.4) augmented with 1×protease halted cocktail (Roche, Germany), displaced by centrifugation at 16,000 g for 20 min at 4°C. The aliquots were diluted with a sample buffer provided by the manufacturer and used to measure the Aβ42 levels by ELISA (Invitrogen, USA).
Immunohistological and microscopic works
For immunohistochemistry, the perfect division was postfixed overnight among 4%paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) at 4°C and cut coronally within 40μm-thick sectors with a vibratome (Leica VT 1000S; Leica Instruments, Nussloch, Germany). Free-floating sections were blocked by 5%regular goat serum, 2%BSA along with 2%fetal bovine serum (FBS). A biotinylated horseradish peroxidase (HRP) system was applied for color development. Anti-Aβ antibody Bam-10 (A5213) (Sigma, St. Louis, MO, USA). Optical microscopy studies were moved out applying an Olympus BX 51 microscope assembled with a DP71 camera along with DP-B software (Olympus Co., Tokyo, Japan). The plaque levels were quantified from microscopic figures of anti-Aβ antibody (Bam-10) dyed superior prefrontal cortex. The amounts of patches in the field were applying TOMORO ScopeEye 3.6 program (Techsan Community, Seoul, Korea).
Western blot analysis
The hippocampal tissues were removed and hom-ogenized in a lysis buffer accommodating 150 mM NaCl, 50 mM Tris-base (pH 7.5), 2 mM EDTA, 10 mM NaF, 10 mM Na-pyrophosphate, 1%glycerol, 1%NP-40, as well as protease inhibitors (5μg/ml aprotinin, 0.1 mM phenylmethylsulfonylfluoride, and 5μg/ml leupeptin). A 30μg sample of tissue lysate was electrophoresed using sodium dodecyl sulfate (SDS)-polyacrylamide gels also conveyed through nitrocellulose membranes. The membranes were then incubated with anti-Aβ42, anti-β-actin, anti-p-Tau, anti-Tau, anti-BACE1, anti-p-AMPKα, and AMPKα (Cell Signaling Technology, Beverly, MA, USA), for 15 h at 4°C. After cleaning with Tris-buffered saline with Tween 20 (TBS-T) (0.05%), the spots were incubated for horseradish peroxidase-conjugated anti-rabbit or anti-mouse IgG, also the bands were featured by applying the ECL system (Thermo Fisher Scientific, USA). Band images were obtained applying a Molecular Imager ChemiDoc XRS+(Bio-Rad, Hercules, CA, USA), and the band severity was evaluated by applying Image Lab™ software version 2.0.1 (Bio-Rad).
Data analysis
All statistical investigations were analyzed with SPSS (ver. 19, Somers, NY, USA). The values are expressed as the means±SEM. All data were considered applying a two-way ANOVA pursued over the Tukey-Kramer multiple connections test. Statistical significance was acknowledged at a p-value lower than 0.05.
RESULTS
Neoline improves learning and memory in the AD mouse model
The effects of neoline on learning and memory in the AD mouse models were assessed by administering neoline in two dosages (low, 0.1 mg/kg/day, Neoline_L or high, 0.5 mg/kg/day, Neoline_H) and donepezil as a positive control for three months beginning at six months of age. The learning and memory improvements were evaluated by performing a passive avoidance test (PAT) initially on day 0, then once a month for three months. At day zero, there was no different latency time between the animal groups (Fig. 1A). On the other hand, at two and three months, neoline and donepezil improved the latency time significantly compared to the normal group but there was no difference between neoline and donepezil group (Fig. 1C, D).
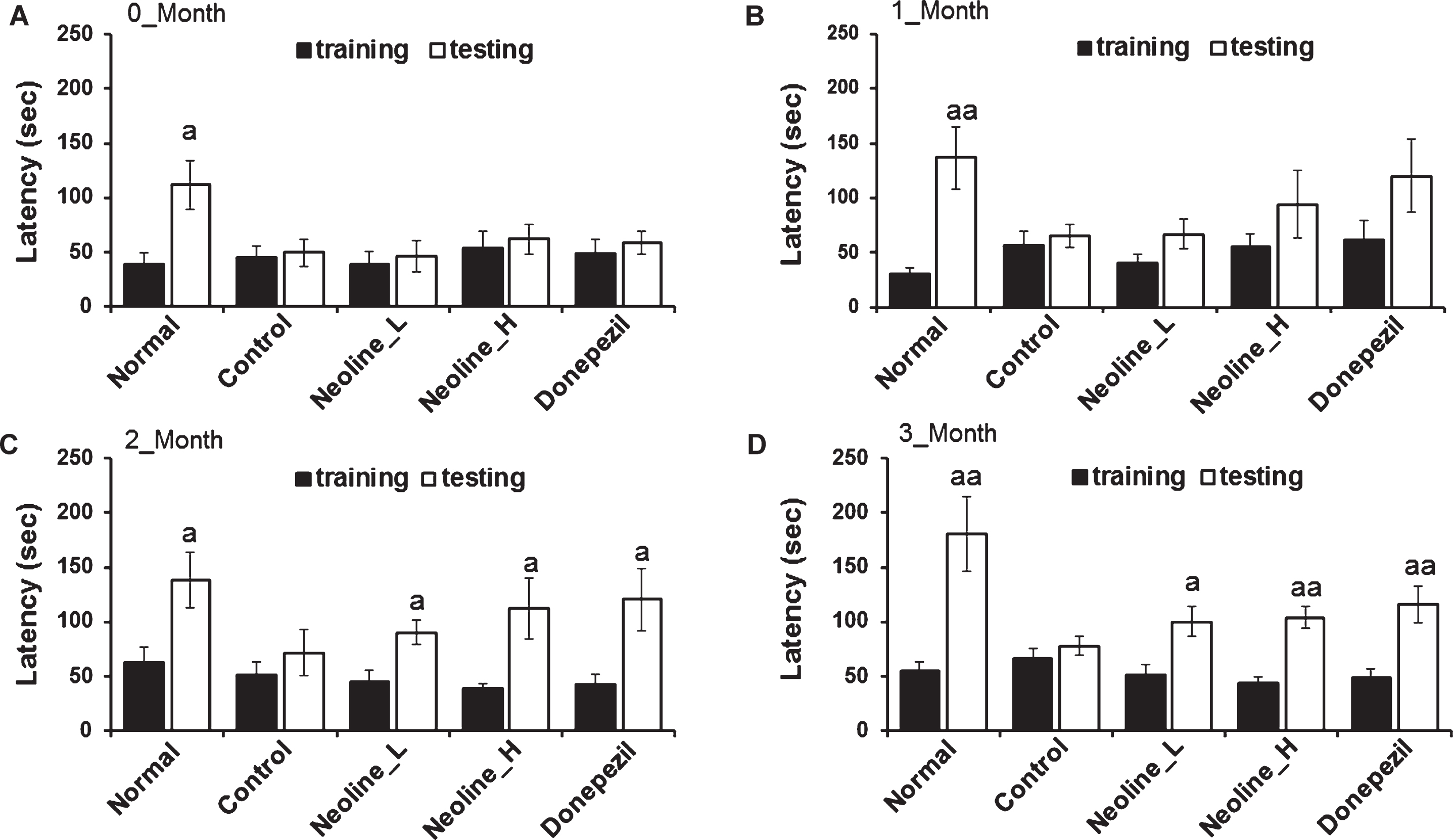
Administration of neoline enhanced the memory deterioration of Tg-APPswe/PS1dE9 mice. In the passive avoidance enforcement test, mice were trained one time, and the mice were then returned to their home cage. One day after training, the passive avoidance achievement was approved. The latency term was specified. Experimental results at the start and after a 0∼3 month treatment (A) start, (B) first month, (C) second month, (D) third month. The data were suggested as the mean±SEM (n = 7, ap < 0.05, aap < 0.01 versus training). Normal: C57BL/6J mice, Control: APP/PS1 mice, Neoline_L: Neoline 0.05 mg/kg/day treated in APP/PS1 mice, Neoline_H: Neoline 0.1 mg/kg/day treated in APP/PS1 mice, Donepezil: donepezil 1 mg/kg/day treated in APP/PS1 mice.
Second, a Y maze test was performed to assess the short-term spatial recognition memory. The spo-ntaneous alteration behavior %, which showed short-term spatial recognition memory, was reduced in the APP/PS AD mice compared to normal mice (Fig. 2A; p < 0.05). The treatment of neoline or donepezil increased spontaneous alteration behavior %significantly in APP/PS AD mice compared to the control APP/PS AD mice (Fig. 2A; p < 0.05).
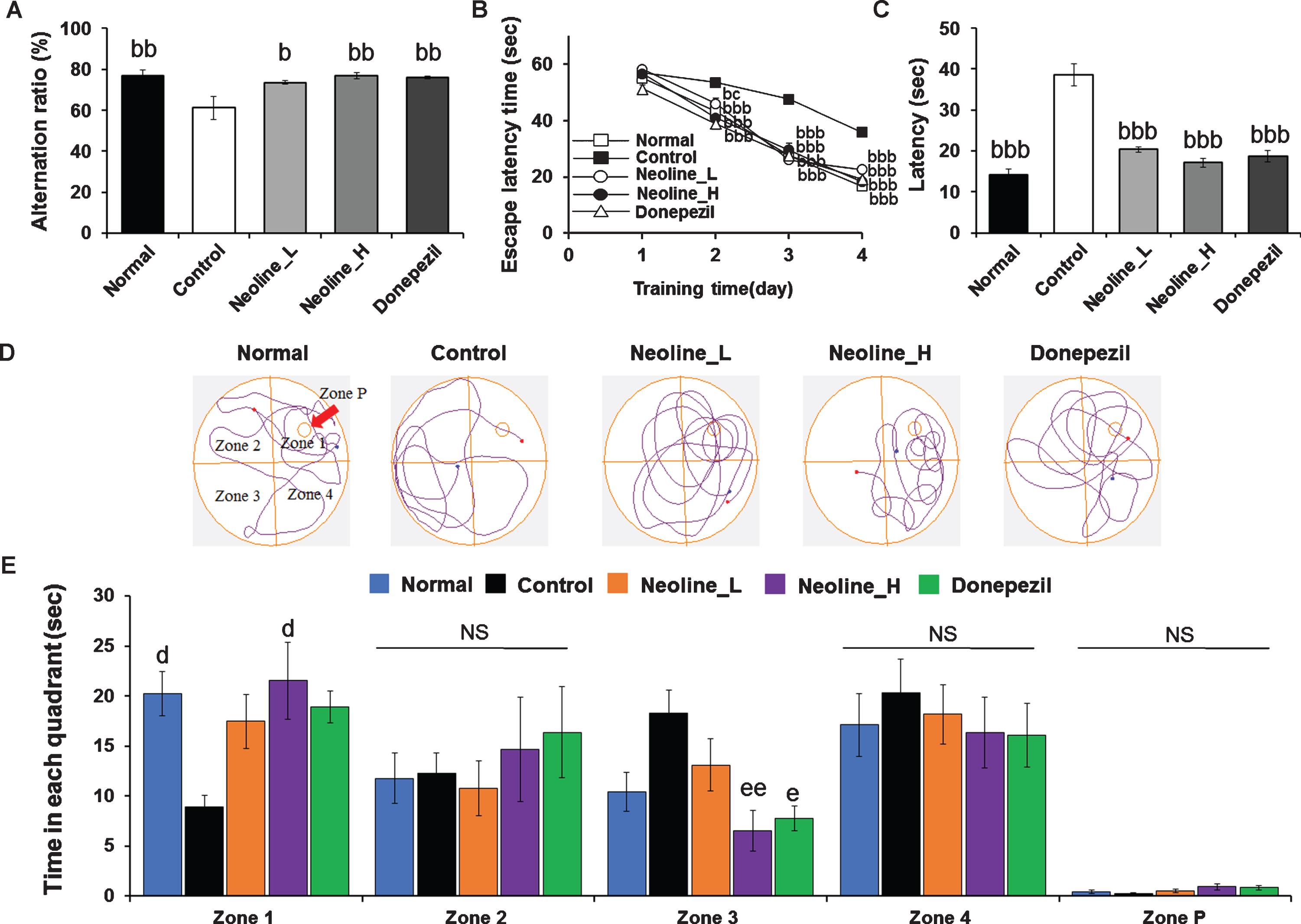
Administration of neoline improved cognitive impairment of Tg-APPswe/PS1dE9 mice. A) Y-maze test and B-E) water maze test. In the water maze test, the animals were qualified for one day, and the memory function was then determined by the escape latencies (s) for four days as described. B) After four days of training. C) Results from the 5th day of the experiment. D) Search paths from representative mice in different groups. E) The time spent in each zone for one minute after platform removal. The results are conveyed as the mean±SEM (n = 7, bp < 0.05, bbp < 0.01, bbbp < 0.001 versus Control; cp < 0.05 versus Donepezil; dp < 0.05 versus Zone 1 Control; ep < 0.05, eep < 0.01 versus Zone 3 Control). Normal: C57BL/6J mice, Control: APP/PS1 mice, Neoline_L: Neoline 0.05 mg/kg/day treated in APP/PS1 mice, Neoline_H: Neoline 0.1 mg/kg/day treated in APP/PS1 mice, Donepezil: donepezil 1 mg/kg/day treated in APP/PS1 mice.
Finally, the anti-AD effect of neoline on the cognitive dysfunction was confirmed using the Morris water maze test. In the training phase, the mice in all groups showed a similar escape latency time (Fig. 2B). On day 4, however, the escape latency time of the neoline or donepezil-treated mice was reduced significantly compared with that of the APP/PS1 mice (Fig. 2B; p < 0.001). In particular, in the post-training probe phase, the neoline mice displayed a comparable escape latency to the normal mice (Fig. 2C; p < 0.001). Furthermore, compared to the APP/PS1 mice, the neoline mice took less time to reach the target quadrant in Zone 3 (platform in Zone 1 location opposite Zone 3 and right/left adjacent Zone 2, Zone 4), and the time spent in the target quadrant was increased significantly in Zone 1 but there was no significance when compared to donepezil (Fig. 2D, E; p < 0.01). These results confirmed that, in this mouse model, neoline treatment might ameliorate the cognitive impairment observed in spatial learning and memory function in AD.
Neoline reduced the anxiety behaviors in the AD mouse model
AD is mostly accompanied by anxiety and depression. Therefore, the anti-anxiety effects of neoline were measured through an open field test, and the anti-depressive effect was examined using a forced swimming test and tail suspension test. The anti-depressant effects of neoline were not observed in either method (data not shown). Nevertheless, neoline increased the time in the exterior significantly and reduced the time in the center compared to the control group, but donepezil did not (Fig. 3A, B, D; p < 0.001). In addition, neoline increased the total distance significantly compared to the control group and donepezil group (Fig. 3C, D; p < 0.001), which was dose-dependent. High-concentrations showed the best effect. That means neoline has better anti-anxiety effects than donepezil.
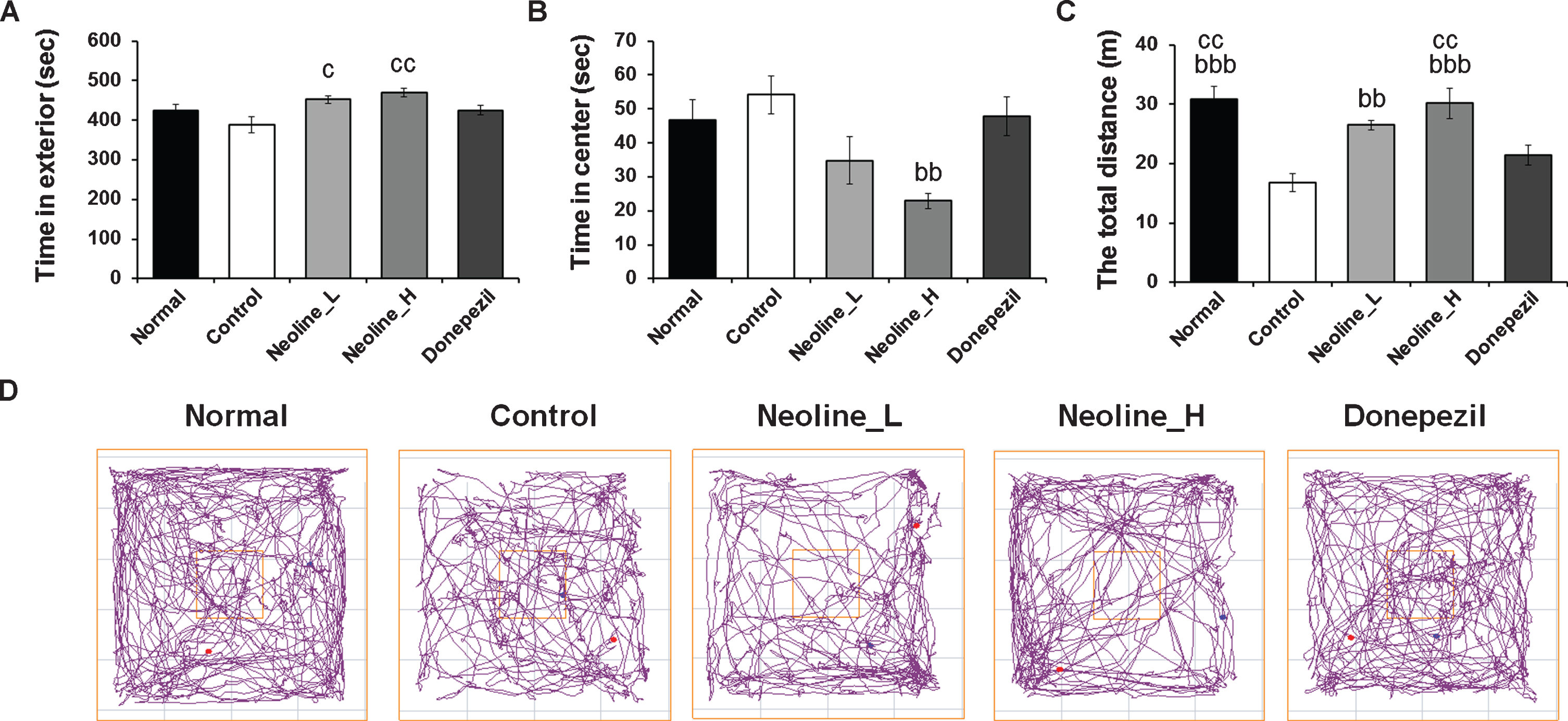
Administration of neoline improved anxiety of Tg-APPswe/PS1dE9 mice. Anxiety was measured with a behavioral test explanatory diagram and the distance the animal moved in the open field test. A) Time in the exterior. B) Time in the center. C) Total distance. D) Search paths from representative mice in different groups. The data are suggested as the mean±SEM (n = 7, bbp < 0.01, bbbp < 0.001 versusControl; cp < 0.05, ccp < 0.01 versus Donepezil). Normal: C57BL/6J mice, Control: APP/PS1 mice, Neoline_L: Neoline 0.05 mg/kg/day treated in APP/PS1 mice, Neoline_H: Neoline 0.1 mg/kg/day treated in APP/PS1 mice, Donepezil: donepezil 1 mg/kg/day treated in APP/PS1 mice.
Neoline decreased the Aβ peptide levels and plaque deposition in the brain of AD mice
ELISA was performed in the prefrontal cortex homogenates to examine the downregulation of the Aβ peptide levels by the neoline treatment. The level of insoluble Aβ1 - 42 was reduced significantly after the high-dose neoline or donepezil treated AD mice compared to the no treated mice (Fig. 4A; p < 0.05). Consistent with the ELISA data, neoline and donepezil also reduced plaque deposition significantly in the prefrontal cortex and hippocampus of Tg-APPswe/PS1dE9 mice in the brain sections stained with anti-Aβ1 - 42 antibody (Fig. 4B, C; p < 0.05).
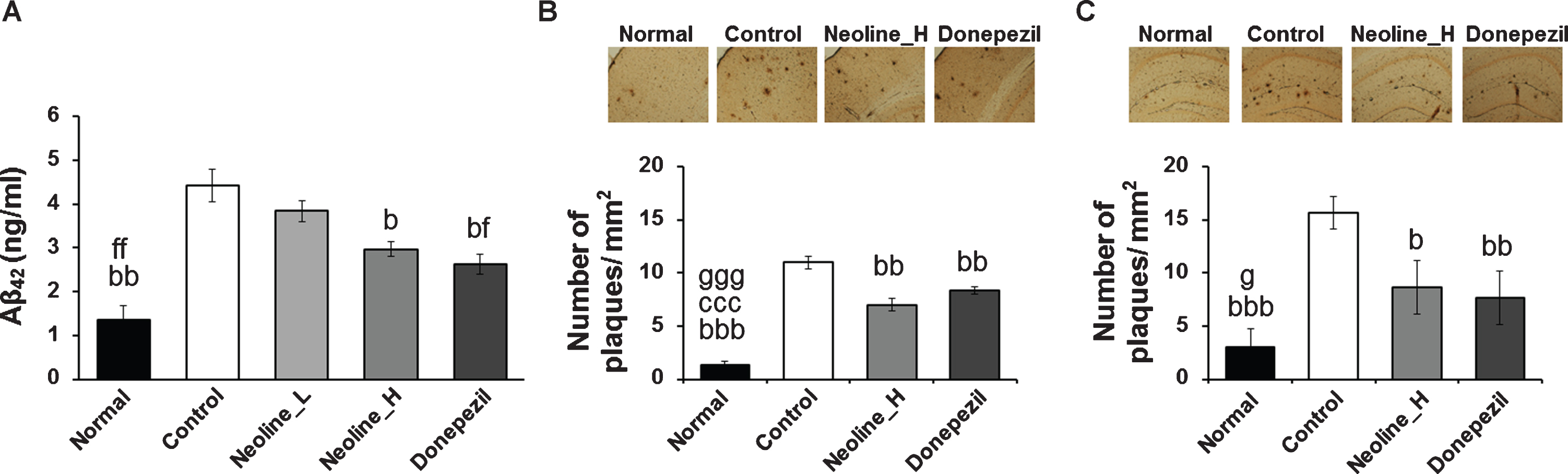
Administration of neoline reduced Aβ42 level and plaque degradation in the brain of Tg-APPswe/PS1dE9 mice. A) ELISA investigation has displaying that the volume of Aβ42 level in the prefrontal cortex of Tg-APPswe/PS1dE9 mouse. B, C) Photomicrographs displaying anti-Aβ antibody (Bam-10)-stained parietal cortex (B) and hippocampus (C) of Tg-APPswe/PS1dE9 mice. Scale bar, 200μm. Plaque numbers in the parietal cortex or hippocampus of Tg-APPswe/PS1dE9 mice were counted using the TOMORO ScopeEye 3.6 program. The data are conveyed as mean±SEM (n = 7, bp < 0.05, bbp < 0.01, bbbp < 0.001 versus Control; fp < 0.05, fp < 0.01 versus Neoline_L; gp < 0.05, gggp < 0.001 versus Neoline_H). Normal: C57BL/6J mice, Control: APP/PS1 mice, Neoline_L: Neoline 0.05 mg/kg/day treated in APP/PS1 mice, Neoline_H: Neoline 0.1 mg/kg/day treated in APP/PS1 mice, Donepezil: donepezil 1 mg/kg/day treated in APP/PS1 mice.
The protein levels of Aβ1 - 42, phosphorylated tau, BACE1, and phosphorylated AMPKα, which are involved in the downregulation of Aβ, were examined to confirm the anti-AD effects of neoline in the hippocampus. Chronic treatment of neoline decreased the levels of Aβ1 - 42, phosphorylated tau, and BACE1 expression significantly and increased the levels of phosphorylated AMPKα in the hippocampus of AD mice (Fig. 5; p < 0.05).
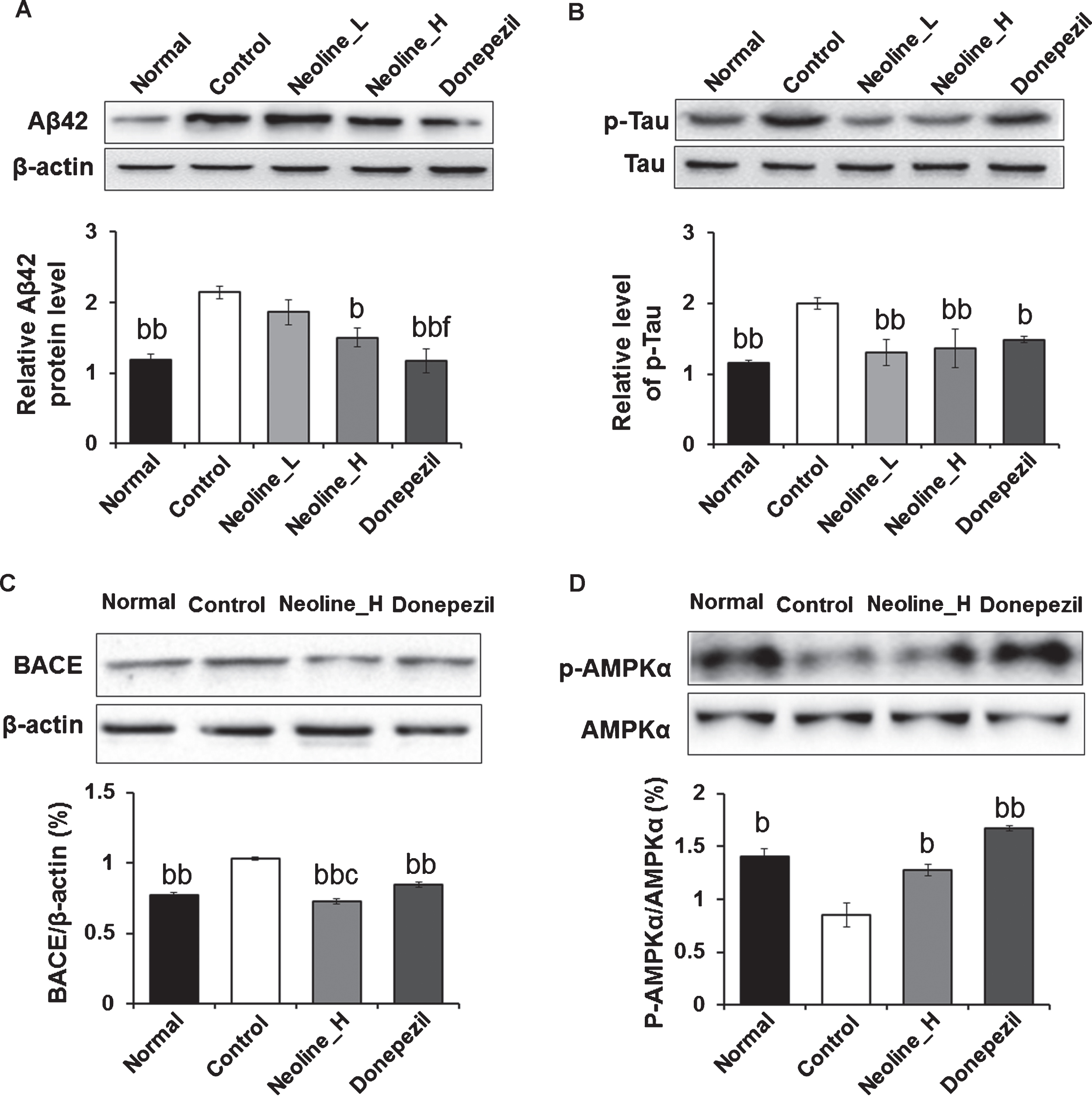
Administration of neoline reduced the Aβ, BACE1, and tau phosphorylation through AMPK phosphorylation in the hippocampus of Tg-APPswe/PS1dE9 mice. The hippocampal lysates were electrophoresed in 10%SDS-PAGE as well as immunoblotted for each antibody. The concentration of the protein bands was calculated by densitometry. The total bands were normalized to β-actin, and the phosphorylated form was normalized versus the total form. The data are communicated as the mean±SEM (n = 7, bp < 0.05, bbp < 0.01 versus Control; cp < 0.05 versus Donepezil; fp < 0.05 versus Neoline_L). Normal: C57BL/6J mice, Control: APP/PS1 mice, Neoline_L: Neoline 0.05 mg/kg/day treated in APP/PS1 mice, Neoline_H: Neoline 0.1 mg/kg/day treated in APP/PS1 mice, Donepezil: donepezil 1 mg/kg/day treated in APP/PS1 mice.
DISCUSSION
The chronic administration of neoline, a single compound from the processed lateral root of Aconitum carmichaelii Debeaux, improved memory impairment and decreased Aβ production and senile plaques through the downregulation of BACE1 in the APP/PS AD mouse models. Moreover, neoline decreased tau phosphorylation levels and induced AMPK activation in the hippocampus of APP/PS AD mouse models, indicating that chronic administration of neoline enhances brain functions involved in learning and memory processes in AD.
The processed lateral root of Aconitum carmich-aelii Debeaux and its components show wide-rea-ching biological activities, including effects on the cardiovascular system, anti-inflammation, analgesic action, anti-tumor activity, effects on the immune system, hypoglycemic and hypolipidemic effects, anti-aging effects, effects of protecting kidney, and effects on energy metabolism [19]. Among them, neoline is an active component displaying effective-ness against a range of neuropathic pain models [15–17, 20] and has a protective action against sodium pentobarbital-induced damage to cardiomyocytes by restoring the beating rhythm and increasing the cell viability [21]. In addition, it was recently reported that neoline ameliorates mechanical hyperalgesia in diabetes by inhibiting the Nav1.7 voltage-gated sodium channel current [22]. It was reported that 1 g of neoline is extracted from 14 kg of Aconitum carmichaelii Debeaux [23] and in clinical study, 4 g of Aconite root is administered in the morning and evening, so neoline is taken 0.58 mg of per day. Based on 60 kg of humans, 0.5 mg of neoline per day converts to 0.1 mg/kg of mice [24]. Therefore, testing at low concentrations of 0.1 mg/kg neoline is similar to the clinical dose and 0.5 mg/kg neoline was used in high concentration. In the pain model, however, the administration route of neoline was a subcutaneous injection, and the dose was 10 mg/kg, which was 20 times more than the dose (0.5 mg/kg) used in this experiment. Moreover, the route of administration was also different: oral administration. Furthermore, the effects of neoline against AD-like phenotypes were manifested by the long-term administration of more than two months (Fig. 1), suggesting that the mechanism of action of neoline against AD might not be related to the inhibition of the Nav1.7 voltage-gated sodium channel.
As mentioned above, Aconitum carmichaelii, incl-uding neoline, is involved in the anti-inflammatory and energy metabolism and pain suppression [19]. In this study, the chronic administration of neoline recovered AMPK activation in the hippocampus of Tg-APPswe/PS1dE9 transgenic mice (Fig. 5D). AMPK functions as a master energy sensor of cells. Essentially, AMPK is activated under low–energy conditions, leading to the inhibition of anabolic reactions and promotion of catabolic processes to maintain energy homeostasis [25, 26]. Moreover, AMPK can phosphorylate the tau protein directly at multiple sites in the microtubule-binding domain and within the flanking regions [27, 28]. Hyperphosphorylation can induce the self-assembly of tau proteins and subsequently to neurofibrillary tangles [29, 30]. On the other hand, AMPK may also reduce the levels of tau and tau phosphorylation through a range of downstream mechanisms, such as silent information regulator type 1 (SIRT1), a class III protein deacetylase, leading to decreased acetylation and degradation of tau protein (including phospho-tau) [31]. Recently, AMPK was reported to reduce tau phosphorylation and improve the brain function in an AD-like model [32]. Although it is unclear which pathway is involved in the inhibition of tau phosphorylation, the administration of neoline inhibited tau phosphorylation in the hippocampus of Tg-APPswe/PS1dE9 transgenic mice (Fig. 5B), suggesting that neoline reduces tau phosphorylation through AMPK activation.
AMPK can also affect amyloidosis through the downregulation of BACE1, a major AβPP cleavage enzyme, and then reduce the production of Aβ [33] and reduce Aβ production by activating autophagocytosis through the mTORC1 signaling, which can facilitate the clearance of Aβ [34]. Thus, efforts to target Aβ clearance by downregulation of BACE1 are useful for improving AD symptoms. In this study, chronic treatment of neoline reduced BACE1 and Aβ expression in the hippocampus, suggesting that neoline-induced AMPK activation may be involved in the downregulation of BACE1, which ameliorate AD-like behaviors and decrease Aβ plaques in the brain of Tg-APPswe/PS1dE9 transgenic mice (Fig. 5C).
Anxiety is associated with cognitive decline/dementia [35], and the amyloid status can be used to predict faster cognitive deterioration in AD [36]. Moreover, anxiety predicts the risk of AD [37]. In this study, neoline ameliorated the anxiety behavior in Tg-APPswe/PS1dE9 transgenic mice, suggesting that a decrease in anxiety symptoms by neoline administration demonstrates the anti-AD effects of neoline.
To compare the anti-AD effects of neoline, donepezil was used as a positive control. Neoline and donepezil had similar effects on increasing cognitive function and reducing Aβ deposition. However, in the anxiety experiments, the group that took neoline significantly improved anxiety, whereas in donepezil, there was no significance. These results suggest that neoline has better anti-anxiety effects than donepezil.
In conclusion, neoline decreased BACE1 expression, causing the suppression of Aβ production and tau phosphorylation through AMPK activation in the hippocampus of Tg-APPswe/PS1dE9 transgenic mice. Moreover, neoline strongly ameliorated the memory impairment and anxiety of APPswe/PS1dE9 transgenic mice. These results highlight the potential of neoline in anti-AD drug research.